
Abstract
Salmonella enterica serovars Heidelberg and Minnesota frequently display several genetic mobile elements making them potential spreaders of resistance genes. Here, we phenotypically determined the antibiotic resistance profile and subsequently performed whole-genome sequencing on 36 isolates recovered from samples of fresh poultry meat, within the Portuguese Official Inspection Plan for Imported Foodstuffs. Several isolates of both serovars showed high genetic relatedness either with isolates from raw poultry meat imported to the Netherlands from Brazil or with isolates from samples from the broiler production chain in Brazil. The multidrug-resistant (MDR) character was common to the vast majority (94.4%) of isolates from both serovars, and several isolates carried the plasmid IncA/C2 containing the β-lactamase gene blaCMY-2 and IncX1 containing a type IV secretion system. These results somehow mirror the scenario observed in the Netherlands, showing the introduction, through fresh imported poultry meat in compliance with European legislation, of MDR Salmonella enterica serovars Heidelberg and Minnesota in Europe, with the potential spread of resistance markers. These data suggest the need to revise the hygiene criteria for foodstuffs monitoring before its placement on the market, with the determination of the resistome being an invaluable contribute to limit the dissemination of resistance markers.
Introduction
One of the main problems that 21st century medicine is facing concerns the rise of multidrug-resistant (MDR) bacteria (bacteria resistant to three or more different classes of antimicrobials).1,2 The use of antibiotics in humans, animals, and plants has led to the widespread of mobile genetic elements carrying multiple antimicrobial resistance traits. 3 As such, the food production industry and its globalization play an important role in the transmission, evolution, and dissemination of clinically relevant MDR pathogens worldwide. 4 Salmonella remains the second most common cause of gastrointestinal disease in the European Union/European Economic Area, 5 and MDR Salmonella have become a major public health concern over the years, causing invasive infections that frequently require hospitalization and present high mortality rates.6,7
In the food chain, poultry, especially chicken and turkey, are regarded as significant contributors for the salmonellosis burden since their intestinal tracts are very often colonized with Salmonella. 8 Additionally, the transmission of Salmonella in flocks may occur from different routes, either from the infected breeding flocks (vertical transmission) or through previously infected flocks, contaminated feed, or water (horizontal transmission). Consequently, the elimination of Salmonella from a facility may prove a difficult task. 9 Several countries, including the United States, Brazil, and the European Union (EU), have implemented, in the past decade, Salmonella control programs in poultry flocks, aimed to reduce the presence of Salmonella spp., especially Salmonella enterica serovar Enteritidis and Salmonella enterica serovar Typhimurium, which cause most part of human infections.8,10
Although these measures resulted in a significant decrease of Salmonella enterica serovar Enteritidis infection (until 2013), they also yielded a shift in the prevalence of Salmonella serovars in poultry and also in human salmonellosis. In fact, less frequent serovars showed a relative increase when compared with previous years, 8 specifically clones possessing advantageous features to deal with the environment and host. These include, for example, tolerance to stress factors, increased adhesion, and invasion properties, usually accompanied with MDR mobile elements.8,11–13
Salmonella enterica serovar Heidelberg and Salmonella enterica serovar Minnesota are rarely reported in European countries, not only in human clinical cases but also in animals and foodstuffs (e.g., poultry meat).5,14 Regarding human cases, data from The European Surveillance System (TESSy) 15 show that between 2012 and 2018, a total of 858 cases of Salmonella enterica serovar Heidelberg and 158 cases of Salmonella enterica serovar Minnesota were reported in the European Union and European Economic Area (EU/EEA), while Portugal reported only one case of Salmonella enterica serovar Heidelberg. However, Salmonella enterica serovar Heidelberg is a frequent cause of gastroenteritis in North and South America and is frequently isolated in poultry meat.16–22
Salmonella enterica serovar Minnesota has also been frequently isolated in poultry production in Brazil.10,23 Portugal is a large consumer and importer of poultry meat, and these foodstuffs are collected within the Portuguese Official Inspection Plan for Imported Foodstuffs coordinated by the General Directorate of Food and Veterinary (DGAV).
In this study, our team at the Portuguese National Institute of Health (INSA) aimed at performing a deep phenotypic and genomic characterization of Salmonella spp. isolated from samples of imported fresh poultry meat. We determined the antibiotic resistance profile on Salmonella enterica serovars Heidelberg and Minnesota and performed whole-genome sequencing (WGS) of the isolates to check their genomic relatedness with publicly available genomes from these serovars and to identify antimicrobial resistance markers and other virulence features.
Materials and Methods
Sample collection and characterization of Salmonella isolates
In total, 163 samples (a sample is composed of several units) of chicken gizzards were collected by the General Directorate of Food and Veterinary, as part of official border control, between 2012 and 2019. For each sample, 25 g of five different representative portions (units) were analyzed for the presence of Salmonella according to VIDAS® SLM (bioMérieux, Marcy-l'Étoile, France), and positive results were confirmed by ISO 6579 method. 24 From a total of 815 units (five units from 163 samples), 57 (from 48 samples) tested positive for Salmonella spp. Due to confidentiality issues, precise data for the analyzed samples were not made available. Thirty-six isolates were randomly selected for further phenotypic and genotypic analysis to have at least one representative isolate from each batch (composed by several samples) of imported fresh poultry meat (Supplementary Table S1).
For genomic analysis (see Gene-by-gene analyses section), a set of 221 genome sequences available in the public databases were also included.
Phenotypic characterization of isolates from poultry meat imported to Portugal
Salmonella isolates were serotyped by the slide agglutination method, according to the Kauffman–White–Le Minor scheme, 25 and antimicrobial susceptibility testing was performed by disc diffusion, following the European Committee on Antimicrobial Susceptibility Testing (EUCAST) 26 recommendations, on a panel of 18 antimicrobials: ampicillin, amoxicillin–clavulanic acid, azithromycin, cefepime, cefotaxime, cefoxitin, ceftazidime, ceftriaxone, chloramphenicol, erythromycin, gentamicin, meropenem, nalidixic acid, pefloxacin, sulfamethoxazole, tetracycline, tigecycline, and trimethoprim. Results were interpreted using the current EUCAST breakpoints and epidemiological cutoff values26,27 (Supplementary Table S1).
Whole-genome sequencing
Total DNA was extracted from the 36 fresh cultures on the NucliSens® easyMAG platform (bioMérieux) according to the manufacturer's instructions. WGS and bacterial de novo assembly were performed as previously described. 28 Briefly, for WGS, high-quality (nonfragmented) DNA samples quantified using Qubit (ThermoFisher, Waltham, MA) were subjected to dual-indexed Nextera XT Illumina library preparation (Illumina, San Diego, CA), before cluster generation and paired-end sequencing (2 × 150 bp) on a NextSeq 550 instrument (Illumina) available at INSA, according to the manufacturer's instructions.
Genome assembly and annotation
All genomes were de novo assembled using the INNUca v3.1 pipeline, 29 an integrative bioinformatics pipeline that consists of several integrated modules for reads QA/QC (i.e., quality assurance/quality control), de novo assembly, and post-assembly optimization steps. Briefly, after reads' quality analysis using FastQC v0.11.530 and cleaning with Trimmomatic v0.36, 31 genomes were assembled with SPAdes 3.1032 and subsequently improved using Pilon v1.18. 33 Draft genome sequences were annotated with RAST server.34–36 All WGS-associated metrics are presented in Supplementary Table S2.
Genotypic characterization of Salmonella isolates from poultry meat imported to Portugal
For each strain, in silico multilocus sequence type (MLST) prediction was performed using the mlst v2.4 software, 37 while serotype was confirmed with SeqSero software 38 using both raw and trimmed reads as well as the assembled genomes.
The ResFinder 3.2 web server 39 (accessed in March 2020) was used to identify acquired antimicrobial resistance genes and/or chromosomal mutations, using a threshold of 80% identity. This analysis focused not only on the genes and mutations that are responsible for the resistance profile to the 18 antimicrobials we tested by disc diffusion but also on colistin and fosfomycin resistance genes. The predicted results were then compared with antimicrobial susceptibility testing results. PlasmidFinder 2.0 and plasmid MLST 40 were used to detect and characterize the plasmids present in these isolates, with a threshold for %ID ≥80% and a minimum % coverage of 60%.
The identification of resistance-associated genes' location in plasmids was confirmed by RAST annotation and blastn suite from NCBI. As a means to potentiate isolate discrimination, assemblies were also analyzed using PHASTER 41 to determine the presence of phages. For simplification purposes, only hits with intact phages were considered for further analysis, given that the detection of phage fragments could result from the assembly process hampering the distinction between the presence of intact phages and phage remnants that were excised during the isolate evolutionary process, for which the biological importance is even more uncertain. Finally, SPIFinder 1.0 (accessed in February 2019) helped to identify Salmonella pathogenicity islands (SPIs) in the sequenced isolates. 42
Gene-by-gene analyses
A broad gene-by-gene analysis was first performed by taking advantage of the Salmonella database 43 hosted on EnteroBase v1.1.244 (accessed January 15, 2020). Briefly, all publicly available genomes of Salmonella enterica serovars Minnesota (n = 262 isolates) and Heidelberg (n = 4,982 isolates) were subjected to the cgMLST V2 schema with 3,002 loci. Serovar phylogenetic inferences were based on the number of shared cgMLST alleles, with unique allelic profiles linked on a single-linkage clustering criterion (HierCC V1).
Considering the worldwide clustering of the Portuguese isolates (see WGS-based genomic relatedness analysis), a narrow gene-by-gene analysis was subsequently performed, targeting only genetically related isolates for which metadata and additional information was available. After a literature review, two studies fulfilling these requisites were selected. As such, along with the 36 isolates from samples of fresh poultry meat imported to Portugal from third countries, 133 serovar Heidelberg isolates identified in the Netherlands, which were present in poultry meat imported from Brazil, 45 as well as 88 isolates of both serovars (82 serovar Heidelberg and 6 serovar Minnesota) identified in poultry meat in Brazil 4 were included in the analysis.
Paired reads of this set of 221 isolates characterized in Brazil and in the Netherlands were downloaded from the European Nucleotide Archive (ENA) (Supplementary Table S3) and assembled as described above. This fine-tune gene-by-gene analysis was performed by taking advantage of a publicly available panel of 8,558 loci 46 derived from the EnteroBase Salmonella wgMLST schema, 43 curated and prepared using the chewBBACA free software, 47 downloaded on August 2018.
For all genomes, allele calling was performed using chewBBACA v2.0.1147 using a BLAST Score Ratio threshold of 0.6 to remove paralogous loci and a publicly available training file for S. enterica. Exact and inferred matches were used to construct an allelic profile matrix, where other allelic classifications were assumed as “missing” loci. To evaluate the genetic relationship between strains, minimum spanning trees (MSTs) were constructed taking advantage of goeBURST algorithm 48 implemented in the PHYLOViZ online web-based tool, 49 based on 100% shared loci between all strains (i.e., shared-genome MLST). 50
To increase the resolution power for cluster analysis, the PHYLOViZ online 2.0 Beta version 51 was used, as it allows maximization of the shared genome in a dynamic manner, that is, for each subset of strains under comparison, the maximum number of shared loci between them is automatically used for subtree construction.
Data availability
All raw sequence reads used in the present study were deposited in the ENA under the study accession number PRJEB32515 (Supplementary Table S2).
Results
Characterization of the isolates
In total, we identified 37 Salmonella enterica serovar Heidelberg isolates, 19 Salmonella enterica serovar Minnesota, and 1 Salmonella enterica serovar 4,[5],12:i:- in fresh poultry meat imported from third countries to Portugal. Serotyping data presented in Supplementary Table S1 were confirmed for all 36 isolates by in silico analysis. Regarding Salmonella enterica serovar Heidelberg, all isolates belong to ST15 with the exception of one isolate that belongs to ST7556. Salmonella enterica serovar Minnesota isolates were found to belong to ST548 (n = 10), ST7557 (n = 1), and ST7558 (n = 1), except one isolate for which it was not possible to determine in silico MLST (Supplementary Table S1).
All the isolates phenotypically tested for antimicrobial resistance by disc diffusion were susceptible to meropenem, chloramphenicol, and cefepime, although 94.4% were multidrug resistant (MDR) (Fig. 1 and Supplementary Table S1). Sulfamethoxazole and tetracycline resistance were the most common (94.0%), followed by nalidixic acid and ampicillin (83.3%), and pefloxacin resistance (80.5%). Resistance to second-generation cephalosporin cefoxitin (77.7%) and to third-generation cephalosporins cefotaxime (80.5%), ceftazidime (77.7%), and ceftriaxone (72.2%) was also frequently observed. Of note, resistance to tigecycline, trimethoprim, gentamicin, and azithromycin was also found in a few isolates (Fig. 1).
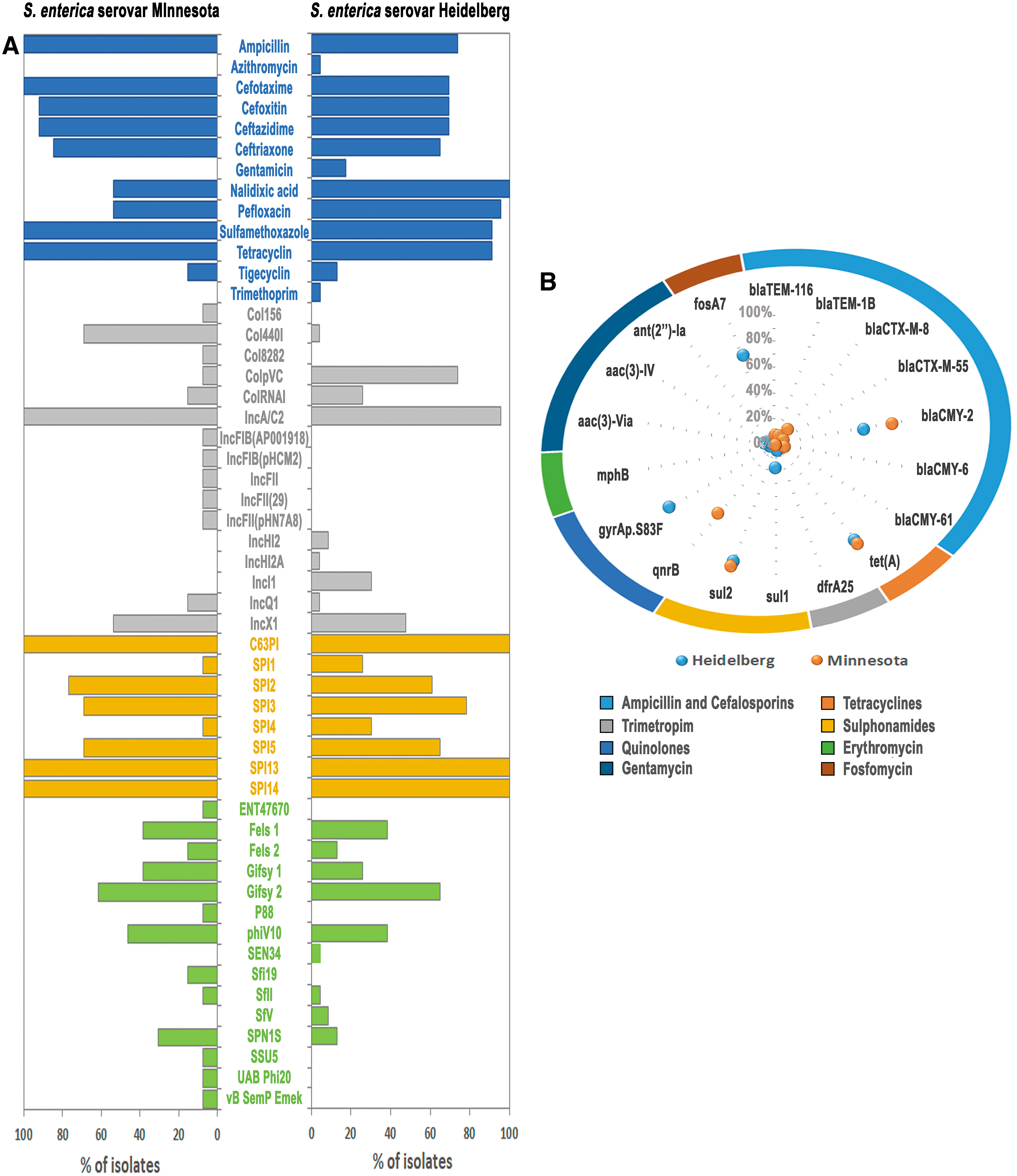
Phenotypic and genotypic data of the Salmonella enterica serovar Minnesota and Salmonella enterica serovar Heidelberg isolates identified in fresh poultry meat samples imported to Portugal.
For the most part, these phenotypic results were confirmed by the in silico predictions of ResFinder. Sulfonamide resistance was mainly caused by the presence of sul2, although four Salmonella enterica serovar Heidelberg isolates additionally presented sul1 (PT8, PT15, PT20, and PT21). Tetracyclines resistance was mediated by tet(A) (Supplementary Table S1). In addition, fluoroquinolone resistance was caused by the presence of mutations in gyrA (S83F) and parC (T57S) in Salmonella enterica serovar Heidelberg, whereas in Salmonella enterica serovar Minnesota, it was mediated by the presence of qnrB genes (Supplementary Table S1).
Generally, β-lactams resistance was caused by the presence of the β-lactamase CMY-2 (77.7%), but other β-lactamase (bla) genes, including blaTEM-1B, blaTEM-116, blaCTX-M-8, and blaCTX-M-55, were also detected (PT2, PT8, PT24, PT34, and PT36) (Supplementary Table S1). The gene mphB, encoding the macrolide 2′-phosphotransferase II, was detected on an erythromycin-resistant Salmonella enterica serovar Heidelberg isolate (PT8), whereas no known mutations associated with macrolides resistance or any common genes were found in the azithromycin-resistant Salmonella enterica serovar Heidelberg PT19 isolate (Supplementary Table S1). Gentamicin resistance was mediated either by an aminoglycoside acetyltransferases [AAC(3)-IV and AAC(3)-Via] or by an aminoglycoside nucleotidyltransferase [ANT(2”)-Ia]. The gene dfrA25 was detected in the trimethoprim-resistant isolate (PT20). Finally, 17 Salmonella enterica serovar Heidelberg isolates presented the gene fosA7, conferring resistance to fosfomycin, and none of the isolates sequenced presented the mcr genes that confer resistance to colistin.
Regarding the presence of plasmids, the 36 sequenced isolates harbored at least one, with 16 different plasmids being detected in total (Fig. 1 and Supplementary Table S1). Overall, we detected 5 different Col plasmids (Col440I, Col8282, ColpVC, Col156, and ColRNAI) and 11 plasmids of 7 different incompatibility groups [IncA/C2, IncFIB(pHCM2), IncFIB(AP001918), IncFII, IncFII(pHN7A8), IncFII(29), IncX1, IncI1, IncHI2, IncHI2A, and IncQ1].
IncA/C2 (ST2) was present in 35 isolates and frequently carried tet(A), sul2, blaCMY-2, and the mercury resistance (mer) operon. In five Salmonella enterica serovar Heidelberg isolates, the plasmidal/chromosomal location of blaCMY-2 gene could not be determined through WGS data. Additionally, IncX1 was present in 18 Salmonella enterica serovar Heidelberg isolates and harbored the type IV secretion system (T4SS). Of note, the mer operon was only absent in the non-MDR isolate PT13. The qnr genes in Salmonella enterica serovar Minnesota were all located in Col440I.
PHASTER analysis identified 14 different phages (Fig. 1 and Supplementary Table S4) among both serovars, 6 in Salmonella enterica serovar Heidelberg isolates (Gifsy-2, Sfi19, UAB Phi20, SPN1S, phiV10, and Gifsy-1) and 11 in Salmonella enterica serovar Minnesota (phiV10, Gifsy-1, Fels-2, Fels-1, SEN34, SfV, vB SemP Emek, SfII, ENT47670, P88, and SSU5).
All Salmonella enterica serovar Heidelberg isolates harbored lambdoid prophage Gifsy-2, which was absent in all Salmonella enterica serovar Minnesota isolates. SPI analysis revealed that all isolates harbor SPI13, SPI14, and Centisome 63 pathogenicity island (C63PI) (Fig. 1 and Supplementary Table S1). Also present in several isolates were SPI1, SPI2, SPI3, SPI4, and SPI5. Of note, SPIFinder and PHASTER results must be faced with caution as they are highly dependent on both the genomic data and on the settings applied for their search. In particular, assemblies originated from short-reads sequencing may strongly underestimate their presence due to the level of genome fragmentation after assembly (Supplementary Table S2).
WGS-based genomic relatedness analysis
When we performed an initial comparative genomic analysis of the Portuguese isolates and all publicly available genomes of these two serovars, we observed a phylogenetic clustering with isolates from the United Kingdom and Brazil for Salmonella enterica serovar Minnesota (Fig. 2) and with isolates from the United Kingdom, Brazil, and Netherlands for Salmonella enterica serovar Heidelberg (Fig. 3). A literature review revealed metadata and additional information solely for the Brazilian and Dutch isolates. These multiresistant serovars were reported in poultry meat in Brazil, 4 and the multiresistant serovar Heidelberg was additionally identified in the Netherlands in poultry meat imported from Brazil. 45
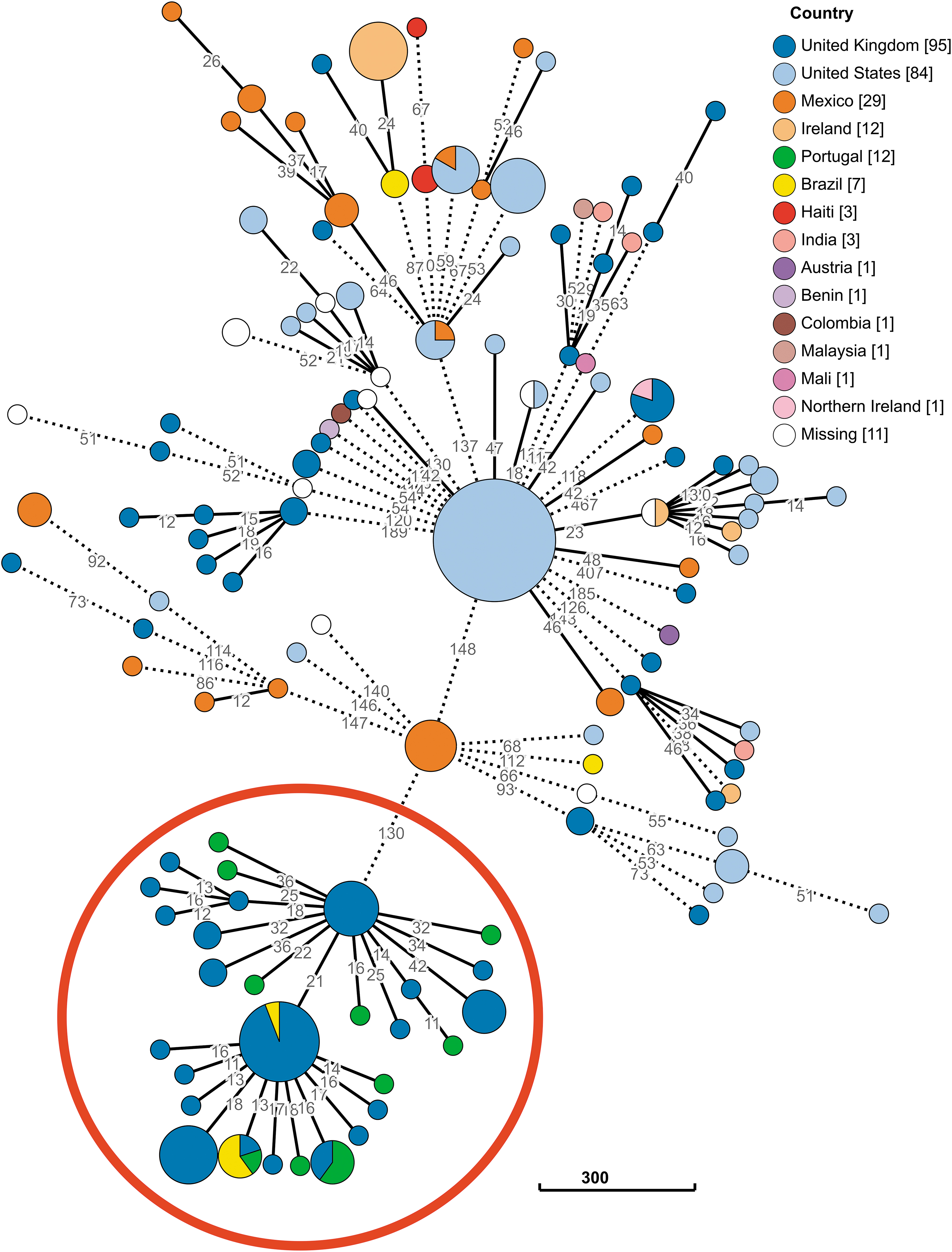
Phylogeny of Salmonella enterica serovar Minnesota based on the cgMLST V2 schema with 3,002 loci from EnteroBase. The MSTree enrolls all Salmonella Minnesota isolates with ST harbored on EnteroBase at 15th January (n = 262), and it was constructed based on the allelic diversity of cgMLST loci. Filled circles (nodes) representing unique allelic profiles are colored according to isolates' isolation country, and are linked on a single-linkage clustering criterion (HierCC V1) with AD shown. For better visualization, branches displaying at least 10 AD were collapsed, whereas branches exhibiting more than 50 AD were shortened and represented by black dashed lines. The red circle highlights the massive cluster where the Portuguese isolates are grouped. Color images are available online.

Phylogeny of Salmonella enterica serovar Heidelberg based on the cgMLST V2 schema with 3,002 loci from EnteroBase. The MSTree enrolls all Salmonella Heidelberg isolates with ST harbored on EnteroBase at 15th January (n = 4,982), and it was constructed based on the allelic diversity of cgMLST loci. Filled circles (nodes) representing unique allelic profiles are colored according to isolates' isolation country and are linked on a single-linkage clustering criterion (HierCC V1) with AD shown. For better visualization, branches displaying at least 10 AD were collapsed, whereas branches exhibiting more than 50 AD were shortened and are represented by black dashed lines. The red circle highlights the massive cluster where the Portuguese isolates are grouped. Color images are available online.
Although the information regarding the origin of the Portuguese isolates is confidential, considering the observed phylogenetic clustering, we hypothesized a scenario similar to the one observed in the Netherlands, that is, Brazil may be the country of origin of Portuguese isolates. As such, along with the 36 genomes of isolates (23 serovar Heidelberg and 13 serovar Minnesota) from samples of fresh poultry meat imported to Portugal from third countries, 133 genomes of serovar Heidelberg isolates from the Netherlands and 88 genomes of isolates identified in Brazil (82 serovar Heidelberg and 6 serovar Minnesota) were included in subsequent analysis.
Regarding serovar Minnesota, the MST analysis revealed that the isolates share 3,458 loci with an overall mean pairwise distance of 85.2 ± 80.6 allelic differences (AD) (median of 32 AD) (Fig. 4). Taking the isolates identified in Portugal into consideration, they only present a mean pairwise distance of 26.3 ± 7.1 AD (median of 27.5 AD), whereas the Brazilian isolates are quite heterogeneous, presenting a mean pairwise distance of 134.6 ± 81 AD (median of 176 AD). However, two Salmonella enterica serovar Minnesota Brazilian isolates (SRR7130551 and SRR7130561) present a mean pairwise distance to the Portuguese isolates of 25.7 ± 6.6 AD.
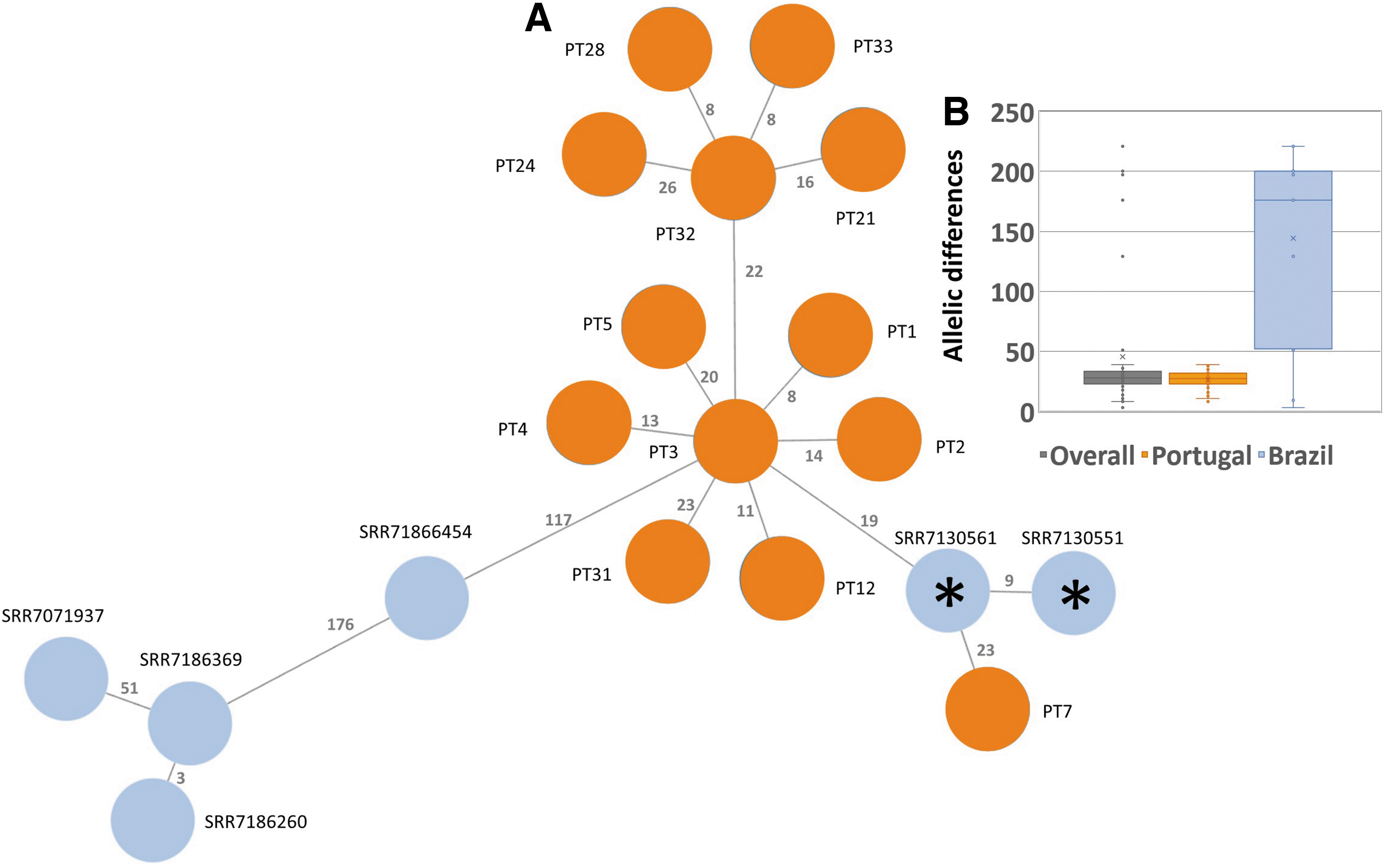
Genomic relatedness between Salmonella enterica serovar Minnesota isolates (n = 19) based on a dynamic gene-by-gene approach and AD.
In addition, they share the same antibiotic resistance phenotype and genotype of some of the isolates. SRR7130551 is resistant to cephalosporins, penicillins, tetracyclines, and fluoroquinolones, 4 presenting the same resistance markers, for instance of isolates PT24 and PT33, including plasmids IncA/C2 and Col440I (Supplementary Table S1). However, SRR7130551 contains an additional pathogenicity island, SPI4, absent in both PT isolates. SRR7130561 presents the same resistance profile and plasmids of PT31, but not SPI4 that is present in this isolate.
Nevertheless, we also found cases for which an apparent discrete allelic distance is not concordant with the differential presence of other genetic features. For instance, PT3 and PT4, isolated from the same batch and revealing 13 AD, present different resistance genes, plasmids and pathogenicity islands (Supplementary Table S1). The differential presence of mobile genetic elements in isolates with a similar genome makeup is not surprising considering the frequent gene exchange that characterizes Enterobacteriaceae,52,53 which may be favored when a mixed population is found in the same batch or food preparation surface.
Analysis of Salmonella enterica serovar Heidelberg isolates revealed that they share 3,041 loci, with a mean pairwise distance of 18.2 ± 9.5 AD (median of 16 AD) between all isolates (Fig. 5). This serovar revealed a low level of genetic variability, as previously described.54,55 Interestingly, isolate PT10, Brazilian isolate SRR7130373, 4 and several Dutch isolates are closely related, sharing a mean pairwise distance of 2.7 ± 2.1. PT11 presents a mean pairwise distance of five AD from several Brazilian isolates, original from Santa Catarina state in Brazil. 45 The same resistance genotype and plasmids were detected in these isolates. Curiously, although PT13 only shares five AD with isolates from the Netherlands, we identified a high level of genetic diversity among these isolates, mainly regarding their antimicrobial resistance profiles.

Genomic relatedness of Salmonella enterica serovar Heidelberg isolates (n = 238) based on a dynamic gene-by-gene approach and AD.
Discussion
One of the most known and appreciated typical dishes in Portugal are stewed chicken gizzards, to an extent that national production is not sufficient to cover the market's requirements. As such, part of this fresh poultry meat consumed in Portugal is imported from third countries. Portugal, being one of the entry points to the EU market, has Border Inspection Posts in its airports and ports, where samples of imported foods of animal origin are inspected by the competent authority DGAV to verify the compliance with the rules and criteria laid down in European legislation. Regarding Salmonella in fresh poultry meat, the Commission Regulation (EC) No 852/200456 and the Commission Regulation (EC) No 2073/2005, 57 and subsequent amendments, 58 lay down both the food hygiene and the food safety criteria for Salmonella in different types of fresh poultry meat.
The specific requirements concerning fresh poultry meat were revised in the light of the serovars of Salmonella causing the higher percentage of human salmonellosis, namely Salmonella enterica serovar Enteritidis, Salmonella enterica serovar Typhimurium, and Salmonella enterica serovar 4,[5],12:i:-. For compliance with the regulation for Salmonella enterica serovar Typhimurium, Salmonella enterica serovar 1,4,[5],12:i:-, and Salmonella enterica serovar Enteritidis, the result must be “Not detected” in 25 g of fresh poultry meat. This means that if any other serovar of Salmonella is detected, the sample is in compliance with the food safety criteria and the foodstuffs are proper for human consumption. However, these regulations do not take into account, for instance, the potential carriage of multiple resistance genes by the detected serovars.
In this study, we aimed to characterize the genotypic (genetic makeup) and phenotypic (antimicrobial resistance profiles and serovar determination) diversity of Salmonella isolates recovered from imported fresh poultry meat between 2012 and 2019. We isolated, from 163 samples (each one comprising 5 units), 37 Salmonella enterica serovar Heidelberg isolates, 19 Salmonella enterica serovar Minnesota, and 1 Salmonella enterica serovar 4,[5],12:i:-. With exception of the latter, all corresponding fresh poultry meat was in compliance with the EU regulations, with no restrictions for commercialization purposes.
WGS of a set of these isolates (involving representatives of all batches) revealed a high genetic diversity within serovar Heidelberg and serovar Minnesota, suggesting the existence of multiple contamination sources. In fact, when comparing the genomic features of these isolates with the ones detected in fresh poultry meat in Brazil 4 and in the Netherlands, 45 we found several potential genetic links, which were also supported by their common antibiotic resistance signature. This suggests the existence of common geographical contaminated sources, translated in several entry points in the EU and in the dissemination of diverse MDR clones.
Although Salmonella enterica serovar Heidelberg and Salmonella enterica serovar Minnesota are infrequently reported in Europe, the dissemination of these two serovars in the American continent has been frequently reported.4,18,20–23,59,60 While the detection of these serovars in fresh poultry meat placed on the market is in compliance with the Commission Regulation (EC) No. 2073/2005, the putative presence of antibiotic resistance markers may constitute a health threat due to the frequent and well-known horizontal contamination. This is especially a concern when those markers confer resistance to antimicrobials used to treat severe infections such is the case of cephalosporins and fluoroquinolones.
CMY-2 has been frequently associated with Escherichia coli isolated from poultry61–63 and also with Salmonella enterica serovar Heidelberg and Salmonella enterica serovar Minnesota.19,23,59,60 Additionally, it has been reported that this resistance marker is easily transferred from exogenous E. coli from human or animal origin to fecal microbiota, proving the potential problem of its spread. 64
In our study, antimicrobial resistance testing of several isolates revealed a high percentage of multidrug resistance (94.4%), which is most likely due to misuse of antimicrobials in poultry production. 23 A high frequency of isolates presenting resistance to sulfamethoxazole, tetracycline, β-lactams, and fluoroquinolones was detected. Fluoroquinolones, sulfonamides, and tetracyclines are usually administered in poultry industry as therapeutic agents for bacterial infections and growth promoters, which might explain the high frequency of isolates with resistance to these groups of antibiotics.23,65 Unfortunately, the small sample size did not allow to draw any conclusion regarding the trend of MDR Salmonella isolation in chicken gizzards. Through WGS, we also found a generalized presence of extended-spectrum beta-lactamase (ESBL) gene blaCMY-2, but also, less frequently, the presence of other widely disseminated ESBL genes. Nevertheless, no phenotypic assays were performed to confirm the ESBL production.
In this work, blaCMY-2 was associated with IncA/C2 other than Inc1, as previously reported.16,59 Typically, IncA/C2 also carried tet(A), sul2, and the mercury tolerance operon. This plasmid is widely distributed across the American continent and is known to be responsible for the dissemination of several important resistance markers. 59 Additionally, IncX1-carrying T4SS was identified in 18 Salmonella enterica serovar Heidelberg isolates. This secretion system is known to play an important part not only in pathogenesis, namely in the ability to invade and persist in macrophages and intestinal epithelial cells, but also in horizontal gene transfer since it can transfer nucleoprotein complexes besides effector proteins.55,66,67 Remarkably, these plasmids also carried toxin–antitoxin systems previously described to confer plasmid stability. 55 Other Inc plasmids were also detected, as well as Col plasmids.
We also detected the gene fosA7, previously identified in Salmonella enterica serovar Heidelberg isolated from broiler chickens in Canada, in 17 isolates of this serovar. 68
Considering that Salmonella can spread to humans through contaminated foods, to prevent or reduce this risk of human infection, whenever Salmonella is identified (but the requirements established in the European legislation for Salmonella in fresh poultry meat are fulfilled), it is recommended that “(…) the batches of those products placed on the market must be clearly labelled by the manufacturer in order to inform the consumer of the need of thorough cooking prior to consumption.” 57 Additionally, the principles contained in the World Health Organization (WHO) Manual “Five Keys to Safer Food” must be followed by consumers and retailers. 69
Nevertheless, although the Portuguese National Reference Laboratory for Gastrointestinal Infections has not identified to date cases of human infection caused by Salmonella enterica serovar Heidelberg or Salmonella enterica serovar Minnesota with these resistance profiles, the presence of these MDR isolates in fresh poultry meat for human consumption is still a risk. In fact, the further spread of these traits to other bacteria, before cooking, represents an added risk that should be taken into account. For instance, in the United States and Canada, Salmonella enterica serovar Heidelberg is more frequently associated with invasive human infections, such as myocarditis and septicemia, than other nontyphoidal Salmonella, and is the second serovar mostly associated with food-related deaths in the United States. 7
In this study, the isolates identified were resistant to fluoroquinolones (ciprofloxacin) and third-generation cephalosporins (ceftriaxone, ceftazidime, and cefotaxime), typically used as treatment for severe Salmonella infections. 70
In summary, we detected two distinct serovars of MDR Salmonella in fresh poultry meat imported from third countries and entering the EU through Portugal. A similar scenario was recently found in the Netherlands through the identification of MDR Salmonella enterica serovar Heidelberg in fresh poultry meat imported from Brazil. Curiously, some of the isolates recovered in Portugal revealed a close genetic proximity and a similar MDR profile to the isolates from the Netherlands (serovar Heidelberg) and also to isolates identified in Brazil (both serovars).
Both serovars present several fitness traits that enhance their capacity to spread and persist alike other successful Salmonella epidemic clones. In an era where the global spread of MDR bacteria is one of the major public health concerns, the criteria for foodstuffs monitoring, especially when concerning imported goods, are quite possibly in need of reviewing. For example, a more cautious approach could include not only the detection of the serovars responsible for most human salmonellosis but also the detection of MDR Salmonella. The surveillance of the antibiotic resistance of Salmonella serovars isolated from fresh poultry meat would be an important contribution for evaluating and understanding the dissemination of antibiotic resistance from animals for food production, to food, and then humans, before foodstuffs of this food category are placed on the market.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.