
Abstract
The emergence and spread of carbapenem-resistant hypervirulent Klebsiella pneumoniae (CR-hvKP) is causing worldwide concern. Sequence type (ST) 86 K. pneumoniae, a major hvKP clone, is rarely resistant to carbapenem. In this study, we report the genomic features and virulence characteristics of a community-acquired bloodstream infection (CA-BSI)-causing CR-hvKP ST86 strain (KPN55602). This strain is resistant to carbapenem but sensitive to amikacin, gentamicin, tigecycline, and colistin. According to in vitro and in vivo virulence assessments, it was classified as hypervirulent. Whole-genome sequencing revealed that KPN55602 has a single 5.13 Mb chromosome and two plasmids. The chromosome of KPN55602 is phylogenetically similar to those of other sequenced ST86 strains. The incompatibility (Inc) group HI1B plasmid pK55602_1, harboring a set of virulence genes, was classified as a virulence plasmid. The IncX6 plasmid pK55602_2, carrying blaKPC-2, was transferable through conjugation and is highly homologous to all five sequenced blaKPC-bearing IncX6 plasmids. In conclusion, to our knowledge, this is the first report of a CA-BSI-causing CR-hvKP ST86 strain harboring an exogenous blaKPC-2-bearing IncX6 plasmid, supplementing existing knowledge on the CR-hvKP evolutionary scenario. The IncX6 plasmid may be an important vehicle for blaKPC, and its horizontal transfer may have led to CR-hvKP evolution in the community setting.
Introduction
K
Although most hvKP isolates remain sensitive to antibiotics, carbapenem-resistant hvKP (CR-hvKP) has recently emerged and been increasingly detected worldwide.7–11 To date, the evolutionary scenario of CR-hvKP appears variable, as acquisition of a virulence plasmid by carbapenem-resistant K. pneumoniae (CRKP) or acquisition of a carbapenemase-encoding plasmid by hvKP may have co-mediated this process.12–14 Klebsiella pneumoniae carbapenemase (KPC) is the most frequently identified carbapenemase worldwide, and KPC-encoding genes have been found on numerous plasmid types, such as incompatibility (Inc) group F, IncA/C, IncR, and IncX. 15 Among them, IncX plasmids have a narrow host range, circulating mainly among Enterobacteriaceae species, and can be phylogenetically grouped into several major IncX subgroups, IncX1 to IncX6. 16
In this study, the microbiological and genomic features of a clinical CR-hvKP strain harboring an atypical virulence plasmid and a KPC-2-encoding IncX6 plasmid were characterized. Our findings will supplement existing knowledge on the molecular epidemiological characteristics of CR-hvKP.
Materials and Methods
Strain identification and hypermucoviscous phenotype detection
The K. pneumoniae strain KPN55602, which exhibits a carbapenem-resistant phenotype (imipenem minimum inhibitory concentration [MIC] ≥4 μg/mL and ertapenem MIC ≥2 μg/mL), was collected in July 2017 at a tertiary hospital in Anhui Province, eastern China. Species identification was performed using a VITEK2 compact system (BioMérieux, Marcy-l'Étoile, France) and confirmed by 16S rRNA sequencing. The HM phenotype was detected using the string test as described previously. 17
Antimicrobial susceptibility testing
The MICs of commonly used antimicrobial agents, including amikacin, aztreonam, cefepime, ceftazidime, ceftriaxone, ciprofloxacin, fosfomycin, gentamicin, and levofloxacin (National Institutes for Food and Drug Control, Beijing, China); ertapenem and imipenem (Merck Sharp and Dohme Corp., Hangzhou, China); meropenem (Sumitomo Pharmaceuticals, Suzhou, China) and piperacillin-tazobactam (Pfizer, New York), were determined using the agar dilution method according to the Clinical and Laboratory Standards Institute guidelines and breakpoint criteria.18,19 The MICs of colistin (Sigma Aldrich, Shanghai, China), tigecycline (Pfizer), and potassium tellurite (Sangon Biotech, Shanghai, China) were determined by the broth microdilution method, whereas the MIC results of colistin and tigecycline were interpreted according to the EUCAST breakpoints. 20 The standard strain Escherichia coli ATCC25922 was used for quality control.
Genome sequencing, assembly, annotation, and comparative analysis
Genomic DNA of strain KPN55602 was extracted from overnight cultures using the QIAamp DNA Mini Kit (Qiagen, Hiden, Germany) according to the manufacturer's protocol. The harvested DNA was detected by agarose gel electrophoresis and quantified by Qubit 2.0 Fluorometer (Thermo Scientific, Waltham, MA). The genome of the strain KPN55602 was sequenced using the PacBio RS II platform (Pacific Biosciences, Menlo Park, CA) and Illumina HiSeq 4000 platform (Illumina, San Diego, CA). Before PacBio RS II sequencing, a 10-kb DNA fragment library was constructed using a single-molecule real-time (SMRT) Bell Express Template Preparation Kit 2.0 according to the manufacturer's instructions.
For HiSeq sequencing, a 300 bp paired-end library with a read length of 150 bp was constructed according to Illumina TruSeq DNA sample preparation recommendation.
Low-quality PacBio subreads (length <1 kb) were filtered out, and then the program Pbdagcon was used for self-correction. All of the filtered sequence were de novo assembled using the Hierarchical Genome Assembly Process in SMRT analysis software version 2.3.0. 21 Subsequently, correction of the PacBio assembly was performed with SOAP tool packages (SOAP2, SOAPsnp, and SOAPindel) using data from the Illumina sequencing. Finally, a single circular chromosome and two plasmids were produced for further analysis.
Functional annotation of the assembled genome sequences was performed by the NCBI Prokaryotic Genome Annotation Pipeline server. Putative virulence factors were predicted using an online database. 22 Antibiotic resistance genes were identified by Resfinder, 23 and the multilocus sequence type (MLST) was identified by submitting the genome sequence to MLST 2.0. 24 Capsular serotyping was achieved by submitting the genome sequence to Kaptive Web. 25 Each plasmid identified in this study was analyzed by PlasmidFinder to investigate the replicons, and sequence comparison was performed using BLASTn analysis and the BLAST Ring Image Generator. 26
Phylogenetic analysis
Single nucleotide polymorphism (SNP) analysis was performed using the CSI Phylogeny 1.4 server. The entire genomes of KPN55602 and other representative hvKP strains available at the National Center for Biotechnology Information website (Supplementary Table S1) were submitted to the website for analysis, and the minimum depth at SNP positions, minimum relative depth at SNP positions, minimum distance between SNPs and minimum SNP quality as input parameters were disabled during the analysis. The read mapping quality was set to a minimum of 25, and the z-score was set to 1.96. The published chromosome sequence of the well-documented hvKP strain NUTH-K2044 chromosome (GenBank accession number AP006725.1) was used as the reference. A maximum likelihood tree was visualized in FigTree 1.4.3.
Conjugation experiment
Conjugative assays were performed using the filter-mating method as previously described with some modifications.27,28 For the virulence plasmid, the donor strain (KPN55602 [the MIC of potassium tellurite was 64 μg/mL]) and the recipient strains (rifampin-resistant E.coli strain EC600 [MIC of potassium tellurite was 1 μg/mL] and three clinical gentamicin-resistant classic K. pneumoniae strains [the MICs of potassium tellurite were 1, 2, and 2 μg/mL, respectively]) were mixed at a ratio of 1:4 and spotted onto filter membranes placed on the surface of Muller–Hinton (M–H) agar plates. After incubation for 18–24 hr at 37°C, transconjugants were selected on China Blue agar plates containing both rifampin (600 μg/mL) and potassium tellurite (8 μg/mL) if the EC600 strain was used as the recipient, or on MacConkey agar containing both gentamicin (16 μg/mL) and potassium tellurite (8 μg/mL) if K. pneumoniae strains were used as the recipient. The presence of rmpA as a marker gene of the virulence plasmid in transconjugants was determined by PCR as described previously. 29
For the carbapenem-resistant plasmid, the donor strain (KPN55602) and the recipient sodium azide-resistant E. coli strain J53 were mixed at a ratio of 1:1, spotted onto a filter membrane placed on the surface of M–H agar plates, and incubated for 18–24 hr at 37°C. Transconjugants were selected on M–H agar containing both sodium azide (300 μg/mL) and ceftriaxone (32 μg/mL). The presence of the blaKPC-2 and other resistance genes in the transconjugants were identified using PCR and DNA sequencing. 11 The MICs of antimicrobial agents for the transconjugants were also assessed.
In vitro and in vivo virulence assessments
The virulence of KPN55602 was assessed by serum killing and Galleria mellonella lethality assays. For references, the hvKP strain KPN54798 (ST23, K1) and the classic K. pneumoniae strain KPN49 (ST11, K47) were employed as a hypervirulent control (H_ctrl) and a low-virulence control (L_ctrl), respectively. 11 For the serum killing assay, human blood was collected from five healthy volunteers and stored at −80°C. The assays were performed, and the results are expressed as previously described. 30
In the G. mellonella infection model, a series of 10-fold serial dilutions of bacterial suspensions containing 103–107 colony-forming units (CFUs) in phosphate-buffered saline (PBS) were injected into G. mellonella larvae (weighing 300 ± 25 mg) (Kaide Ruixin Co., Ltd., Tianjin) for 50% lethal dose (LD50) experiments. Twenty larvae were injected at each dilution; the larvae were incubated at 37°C and monitored every 12 hr for 72 hr. The experiments were performed three times on separate occasions. The survival rates were recorded, and the LD50 values were calculated using the Bliss method. 31 Negative control groups included larvae that were inoculated with 10 μL of PBS. 32
Clinical information collection and ethics
Clinical information was collected from electronic medical records. This retrospective study was approved by the ethics committee of the Second Hospital of Anhui Medical University with waiver of informed consent (approval number PJ-YX2018-001).
Nucleotide sequence accession numbers
The genome sequence of K. pneumoniae KPN55602 was submitted to GenBank under the accession numbers CP042977 (KPN55602 chromosome), CP042975 (plasmid pK55602_1), and CP042976 (plasmid pK55602_2).
Statistical analysis
The Shapiro–Wilk method was used to test normality. Normally distributed variables were summarized as the mean ± standard deviation. The serum killing assay data were summarized as the mean ± standard error. Survival data were plotted using the Kaplan–Meier method and analyzed using log-rank tests with GraphPad Prism 7.0 (GraphPad Software, La Jolla, CA). The LD50 values were expressed as log10 (lg) transformed values. A two-sample t-test was used to compare the data and was performed using SPSS v. 21.0 (IBM Corp., Armonk, NY). A two-sided p-value <0.05 was considered statistically significant.
Results
Clinical information
A 64-year-old man was admitted to the emergency department due to high fever and septic shock. This patient had diabetes and had not been hospitalized within the 3 months before this admission. He immediately received relevant supportive treatment and an empirical anti-infection treatment with meropenem (1 g, intravenous [i.v.], q8h period 3 days) after admission. Meanwhile, blood culture specimens were collected, and the strain KPN55602 was isolated from them. KPN55602 was identified as K. pneumoniae, and it showed resistance to cephalosporins, aztreonam, β-lactam/β-lactamase inhibitor, carbapenem, fosfomycin, and fluoroquinolones, but susceptibility to amikacin, gentamicin, tigecycline, and colistin (Table 1). Further examination revealed an abscess in the deep anterior side of the right lower limb.
Antimicrobial Susceptibility Patterns and Resistance Mechanism of the Strain KPN55602 and Its Transconjugant
The unit of MIC is μg/mL.
The corresponding coding sequence locus_tag of antimicrobial resistance genes in the genome is labeled in the parentheses, and the antimicrobial resistance genes harbored by the plasmid are underlined.
AMK, amikacin; ATM, aztreonam; ARG, antimicrobial resistance gene; CAZ, ceftazidime; CTX, ceftriaxone; CIP, ciprofloxacin; COL, colistin; ETP, ertapenem; FEP, cefepime; FOS, fosfomycin; GEN, gentamicin; IPM, imipenem; I, intermediate; LVX, levofloxacin; MEM, meropenem; MIC, minimum inhibitory concentration; ND, no detection; S, susceptible; R, resistant; TGC, tigecycline; TZP, piperacillin-tazobactam; Δ, truncated gene.
Through surgical debridement, combined treatment with amikacin (0.6 g, i.v., qd) and tigecycline (100 mg, i.v., q12h) for 7 days, and continuous irrigation with gentamicin-containing solution, the patient's temperature was gradually controlled and returned to normal. However, there was a deficiency in that the pus specimen was not collected for bacterial culture detection when the abscess was incised and drained. Finally, through systematic treatment, the patient was eventually cured and discharged after 21 days in the hospital. Since the time interval between the hospital admission and the positive blood culture specimens collected was within 48 hr, the bloodstream infection (BSI) suffered by this patient was defined as a community-acquired (CA) infection.
Genome features and SNP analysis
The K. pneumoniae strain KPN55602 was determined to be ST86 and capsular serotype K2, and its colony formed mucoid strings of <5 mm, although the colonies were mucoid on Colombia blood agar plates. The genome of KPN55602 comprises a single chromosome (5,127,934 bp) and two plasmids: pK55602_1 (270,776 bp) and pK55602_2 (43,296 bp). In total, 5,326 gene were identified, including 5,036 coding genes, 88 transfer RNAs, 25 ribosomal RNAs, 11 noncoding RNAs, and 166 pseudogenes. The final draft genome of the chromosome had a G + C content of 57.71%, and whole-genome SNP-based phylogenetic analysis showed that it is highly similar to the sequenced ST86 hvKP strains NUHL30457, hvKP1, HK787, and CG43 but distant from the clade composed of hvKP belonging to other STs (Fig. 1).
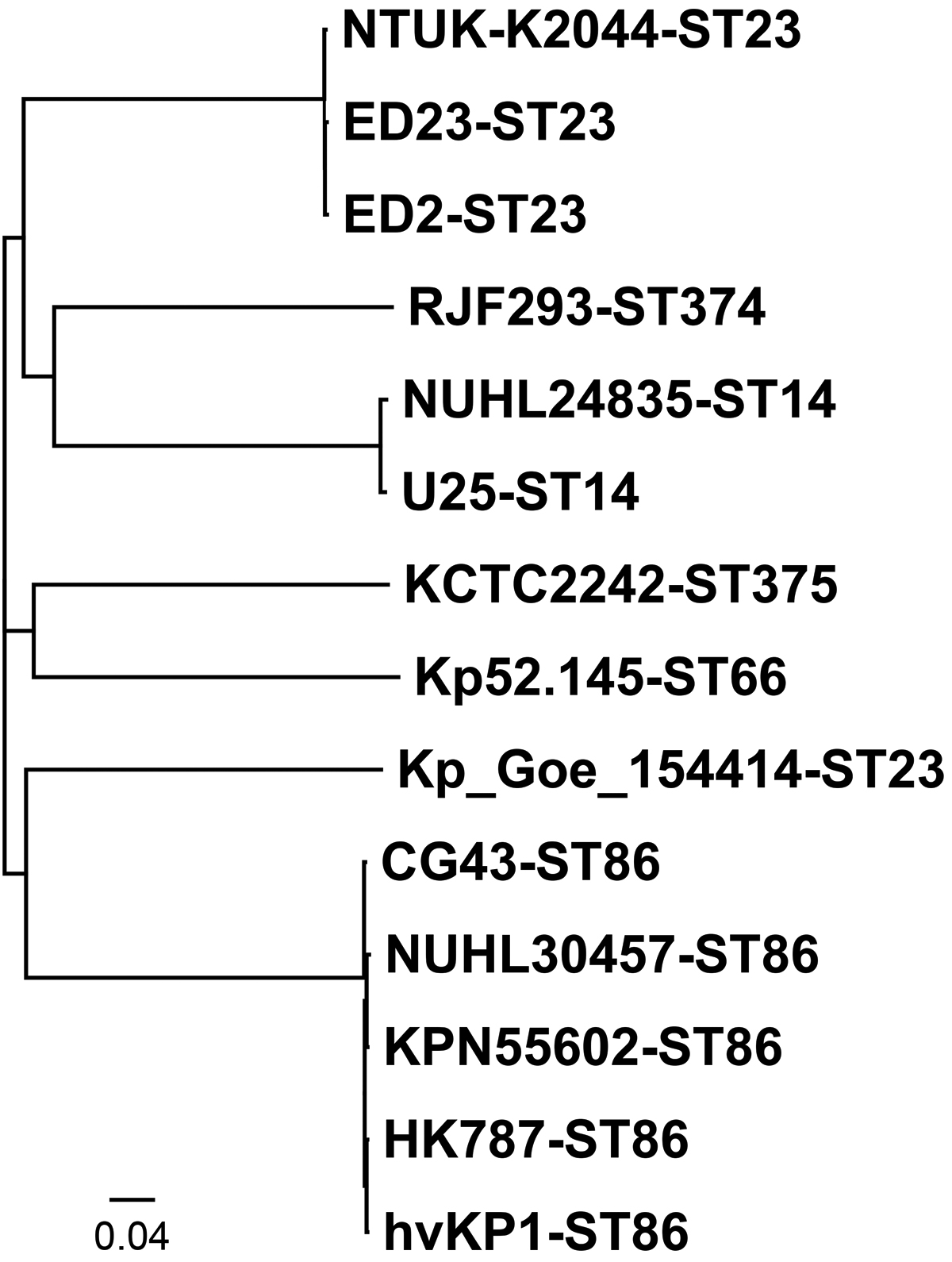
Phylogenetic tree of KPN55602 relative to other sequenced hvKP strains. The scale bar indicates a distance of 0.04 nucleotide substitutions per site. Each entry is represented by the isolate's name followed by the ST. ST, sequence type; hvKP, hypervirulent Klebsiella pneumoniae.
BLASTx analysis was performed to identify a number of antimicrobial resistance genes. Among them, blaKPC-2 and ΔblaTEM-1 were found to be located on the smaller plasmid (pK55602_2). The other antimicrobial resistance genes located on the chromosome are involved in the multidrug resistance mechanism, including β-lactamase (blaSHV-28), fosfomycin resistance gene (fosA), and fluoroquinolone resistance genes (oqxA/oqxB) (Table 1).
According to the virulence and heavy metal resistance gene analysis, aerobactin receptor gene (iutA), enterobactin gene cluster (entABCDEFS/fepABCDG), salmochelin gene cluster (iroEN), type 1 fimbriae gene cluster (fimABCDEFHIK), type 3 fimbriae gene cluster (mrkABCDFHIJ), copper resistance gene (nlpE), nick/cobalt resistance genes (rcnAB), silver resistance gene (cusR), and tellurium resistance genes (tehAB) are present on the chromosome, with mucoid factor encoding gene (rmpA), phoPQ-activated integral membrane protein gene (pagO), salmochelin genes cluster (iroBCDN), and tellurium resistance gene cluster (terABCDEFWXZ) present on the larger plasmid pK55602_1 (Table 2). Notably, further sequence alignment analysis revealed that the rmpA gene is considered to be a pseudogene due to its sequence containing frameshift mutation (Supplementary Fig. S1).
General Genomic Features of the Strain KPN55602 a
The corresponding coding sequence locus_tag of the main virulence and heavy metal resistance genes is listed in Supplementary Table S2.
Pseudogene.
Comparative genomic analysis of the virulence plasmid and conjugation experiment
Sequence analysis showed that pK55602_1 is a large virulence plasmid with a length of 270,776 bp and contains 249 coding genes. This plasmid belongs to the IncHI1B group with an average G + C content of 50.93%. BLASTn revealed that the sequence of pK55602_1 can be roughly divided into two distinct regions. The first region, the 1st to 150th kb, is highly similar to the sequences of the plasmids pLVPK (GenBank accession number NC_005249.1), pK2044 (GenBank accession number NC_006625.1), p30457-1 (GenBank accession number NZ_CP026587.1), pRJF293 (GenBank accession number NZ_CP014009.1), and pRJF999 (GenBank accession number NZ_CP014011.1), with 92–93% query coverage at 99% identity.
Virulence genes, including iroBCDN, pagO, and rmpA (pseudogene), which were previously reported to be associated with hvKP, are located in this region, but iucABCDiutA and rmpA2 are absent. Conversely, no known plasmid sequences closely matched those of the second region (Fig. 2). Conjugation experiments showed that the virulence plasmid pK55602_1 was not transferred to the recipients.
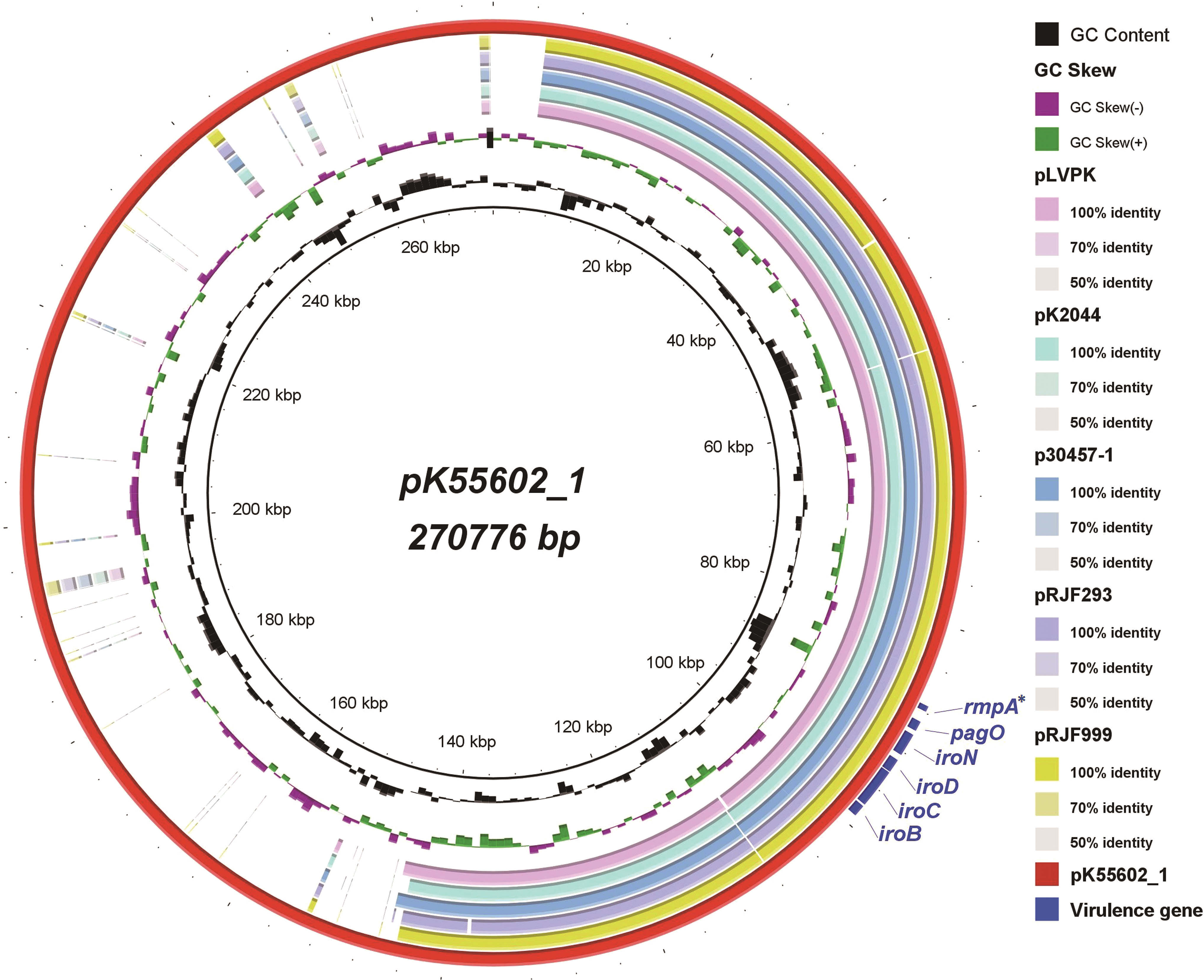
Genomic sequence comparative analysis of plasmid pK555602_1 and five reported virulence plasmids harbored by hvKP strains. The circles from inside to outside indicate the GC content of pK555602_1, GC skew of pK555602_1, and genomic sequences of the plasmids pLVPK, pK2044, p30457-1, pRJF293, pRJF999, and pK555602_1. The white and colored regions of the rings indicate absence and presence, respectively. The locations of the virulence genes iroBCDN, pagO, and rmpA are indicated. “*”, the rmpA gene harbored by plasmid pK55602_1 is a pseudogene. Color images are available online.
Comparative genomic analysis of the plasmid harboring blaKPC-2 and conjugation experiment
The sequence analysis showed that the plasmid pK55602_2 is 43,296 bp in length and belongs to the IncX6 group. Furthermore, BLASTn results revealed that this plasmid is genetically closely related to the five previously sequenced IncX6 plasmids, including pE20-KPC, pGN2-KPC, pGN26-KPC, pGN28-KPC (GenBank accession numbers MF156709 to MF156712), and pKPC3_SZ (GenBank accession numbers KU302800) (Fig. 3). Notably, with 100% query coverage and 99% identity by BLASTn, the genetic sequence of pK55602_2 is highly consistent with that of plasmid pKPC3_SZ, the first identified IncX6 plasmid. In conjugation experiments, the plasmid pK55602_2 harboring blaKPC-2 was transferred to E. coli J53, and PCR detection confirmed that the transconjugant (J53-KPN55602) was positive for blaKPC-2 and ΔblaTEM-1. Moreover, phenotypic testing of the transconjugant showed that it exhibited resistance to cephalosporins, aztreonam, and carbapenem but susceptibility to ciprofloxacin, levofloxacin, amikacin, gentamicin, tigecycline, and colistin (Table 1).

Genomic sequence comparative analysis of plasmid pK55602_2 and all five sequenced IncX6 plasmids. The circles from inside to outside indicate the GC content of pK555602_2, GC skew of pK555602_2, and genomic sequences of the plasmids pE20-KPC, pGN26-KPC, pGN28-KPC, pGN2-KPC, pKPC3_SZ, and pK555602_2. The white and colored regions of the rings indicate absence and presence, respectively. The locations of the resistance genes blaKPC-2 and ΔblaTEM-1 are indicated. Δ, truncated gene; KPC, Klebsiella pneumoniae carbapenemase. Color images are available online.
In vitro and in vivo virulence assessments
The serum killing assay demonstrated that both KPN54798H_ctrl and KPN55602 were serum resistant (grade 6 and grade 5, respectively), whereas KPN49L_ctrl was serum sensitive (grade 1) (Fig. 4a). In the G. mellonella infection model, time- and dose-dependent mortality was observed for all isolates tested. At the dose of 106 CFU/larvae, the observed larval lethality rate due to KPN55602 was higher than that due to KPN49L_ctrl (log-rank test, p < 0.0001), and it was not significantly different from that due to KPN54798H_ctrl (log-rank test, p = 0.072) (Fig. 4b). Data for the other doses of the tested strains are available in Supplementary Fig. S2. The lg LD50 of G. mellonella larvae due to KPN55602 (4.89 ± 0.18) was lower than that due to KPN49L_ctrl (6.21 ± 0.13) (two-sample t-test, p = 0.001), but there was no significant difference from that due to KPN54798H_ctrl (4.75 ± 0.13) (two-sample t-test, p = 0.343).
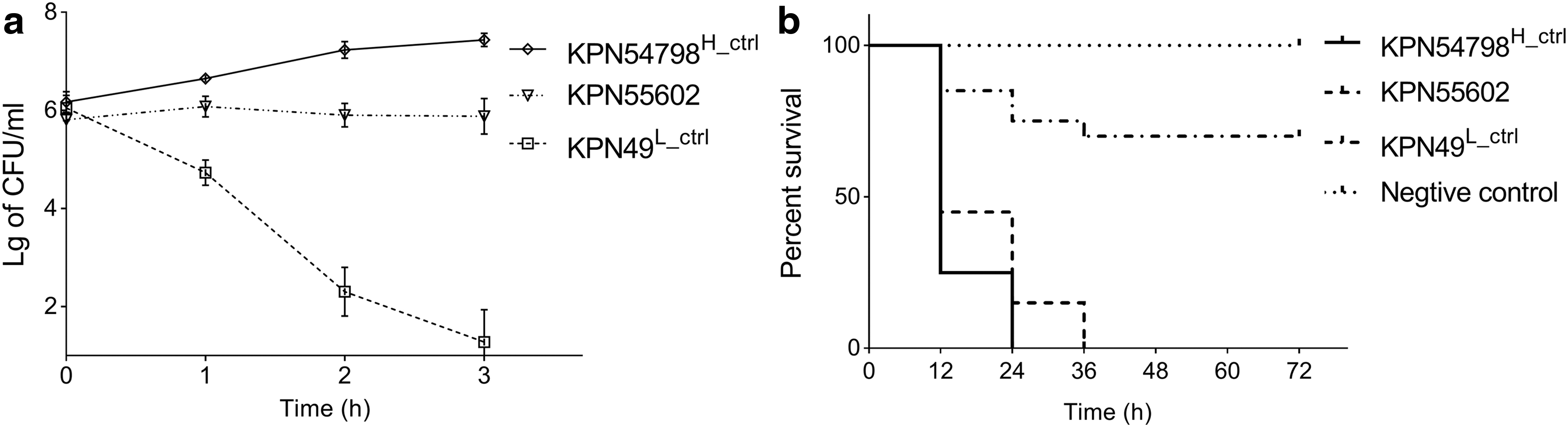
Virulence assessment of the strain KPN55602.
Discussion
In this study, the K. pneumoniae ST86 strain KPN55602 was isolated from the blood of a patient who acquired infection in a community setting. This strain was not only as virulent as the reference ST23 hvKP strain but also resistant to carbapenem. To the best of our knowledge, this is the first report of a CA-BSI caused by an ST86 CR-hvKP, and our findings expand the CR-hvKP epidemiological database worldwide. To date, CA infection caused by CR-hvKP is still rare. 4 A recent study in eastern China reported a severe CA infection caused by a blaKPC-2 and blaNDM-1 co-harboring CR-hvKP strain NUHL30457; coincidentally, NUHL30457 also belongs to ST86. 33 Further whole-genome SNP-based phylogenetic analysis showed that the chromosomes of both isolates were highly similar to each other, suggesting that they share a common ancestor.
As an encapsulated pathogen, K. pneumoniae with the K2 serotype is strongly associated with hvKP. A previous study in Asia showed that two major ST groups, ST65-like and ST86-like, are strongly linked to K2 hvKP, with its colonies often exhibiting a hypermucoviscous phenotype. 4 Our study results show that the KPN55602 strain belongs to ST86 and serotype K2, although its colonies formed mucoid strings of <5 mm and it displayed a mucoid phenotype on Colombia blood agar plates. Two virulence genes, rmpA and rmpA2, which both contribute to the enhanced capsular production, have been defined as key regulators of the hypermucoviscous phenotype. 34 Atypically, in addition to the rmpA gene having as frameshift mutation, the rmpA2 gene was absent. According to some recent research, the mutation or absence of rmpA and rmpA2 genes results in a negative hypermucoviscosity phenotype in hvKP.35–37 Hence, in this study, the aforementioned reasons both contribute to that the colonies of KPN55602 did not exhibit a hypermucoviscous phenotype.
In addition to capsular serotypes and STs, a variety of virulence factors are associated with hvKP strains. Notably, a large virulence plasmid has been detected in all whole-genome-sequenced hvKP clonal lineages. Furthermore, this plasmid, which harbors iroBCDN, iucABCD, pagO, rmpA, and rmpA2, was found to be restricted to hvKP isolates. Among them, the iucABCD gene cluster encoding aerobactin was not carried by pK55602_1. Aerobactin is considered one of the most important siderophores, accounting for increased siderophore production, and is a major virulence determinant of hvKP. 38 Our in vitro and in vivo virulence assessments showed that the virulence of KPN55602 was similar to that of the ST23 reference strain KPN54798H_ctrl, which is positive for iucABCDiutA. 11 This result suggests that more complicated virulence mechanisms of hvKP may have yet to be revealed.
In China, the dominant clone of CRKP is ST11, and several studies have reported that most CR-hvKP strains harboring a virulence plasmid also belong to ST11.13,14,39 Hence, it has been speculated that the horizontal transfer of virulence plasmids is a major contributor to the evolution of CR-hvKP. However, CR-hvKP strains belonging to other ST have continued to emerge, indicating that the evolutionary scenario is not uniform.36,40,41 Our results show that the blaKPC-2-bearing plasmid pK55602_2 could transfer to a recipient by conjugation and that the MIC values of all β-lactams, including carbapenem, increased in the transconjugant (J53-KPN55602) compared with those in the parental strain J53. It is suggested that the horizontal transfer of this plasmid can cause dissemination of carbapenem resistance determinants.
Further BLASTn analysis revealed that the sequence of pK55602_2 is highly consistent with that of pKPC3-SZ, the first identified IncX6 plasmid, which was reported in 2016. 16 Notably, pKPC3-SZ was carried by a clinical Enterobacter cloacae strain causing lethal infection in a tertiary hospital in Jiangsu Province, eastern China. 16 Therefore, plasmid pK55602_2 may be an exogenous plasmid that was acquired by KPN55602. Another recent study also reported dissemination of the pKPC3-SZ-like IncX6 plasmid among multiple Enterobacteriaceae species in a single Chinese hospital in Beijing, Northern China. 42 Based on these findings, the IncX6 plasmid appears to be an important vehicle for the blaKPC gene in China.
In conclusion, we characterized the genomic features and virulence characteristics of a CA-BSI-causing CR-hvKP isolate, and although it did not exhibit a hypermucoviscous phenotype, its serum resistance and low LD50 confirmed the hypervirulence of the strain. Genomic analysis showed that this strain belongs to ST86 and serotype K2, a major hvKP clone, and harbors an exogenous KPC-2-encoding plasmid, suggesting that the horizontal transfer of carbapenem-encoding plasmid could have led to the evolution of CR-hvKP. Our study highlights an urgent need to monitor the plasmid-mediated convergence of hypervirulence and carbapenem resistance in K. pneumoniae in the community setting.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.