
Abstract
Multidrug-resistant extended-spectrum beta lactamase (ESBL)-producing Escherichia coli strains are emerging globally in both humans and animals. Antimicrobial susceptibility testing and ESBL screening were performed on pure cultures of 216 E. coli isolates from human and animal fecal samples as well as beef. Polymerase chain reaction was performed for the detection of resistance genes. Representative isolates of ESBL-producing E. coli were randomly selected for multilocus sequence typing and pulsed field gel electrophoresis (PFGE). Sixty of the isolates were identified as ESBL producers, and seven resistance genes were amplified in them: TEM (61.7%), blaCTX-M-15 (51.7%), AAC-6-LB (43.3%), blaCTX-M-1 (38.3%), blaCTX-M-9 (33.3%), blaCTX-M-2 (21.7%), and SHV (11.7%); they were classified into four phylogroups: A (25%), B1 (45%), B2 (20%), and D (10%). Thirty of these isolates were clustered into 10 sequence types with ST131 being mostly prevalent. Six PFGE types were discovered, each of which was shared by isolates from different subjects and had the same phylogroups and resistance gene profiles. There was a dissemination of PFGE types across various groups among humans, animals, and beef. This underlines the fact that the spread of ESBL E. coli could be from humans to animals, from animals to humans, as well as across animal species.
Introduction
E
The development of resistance, especially by extended-spectrum beta lactamase (ESBL)-producing bacteria, is emerging worldwide in both humans and animals. 6 Antibiotic-resistant bacteria constitute a major threat to public health, as the antibiotic resistance determinants can be transferred to other pathogenic bacteria, potentially compromising the treatment of severe bacterial infections. 6 The World Health Organization (WHO) reported that ∼50% of the universally produced antibiotics are used for veterinary purposes such as prophylactic and growth promotion.2,7 The overuse and misuse of these antimicrobials can lead to the selection of resistant strains and proliferation of antimicrobial resistance (AMR) in human and animal pathogens and commensals.7,8
The multidrug-resistant strains of bacteria and their resistance genes have been reported to be disseminated from food-producing animals to humans by direct contact or via their food products. 7 The dissemination of antimicrobial resistant strains from humans to animals may also be possible as exchange of bacteria could occur both ways. Various genetic tools have been used to establish this genetic relatedness among human and animal isolates.8–10
With the poor personal hygiene, inadequate toilet facilities (at homes and in public places), and poor sewage systems that are rife in Nigeria, human to human dissemination of antimicrobial-resistant bacteria strains should not be ignored. Most farm animal production settings in Nigeria lack basic requirements for appropriate waste disposal; most of these farms are located either within vicinities of homes or within the major cities in urban settings, thereby aiding prompt dissemination of AMR genetic traits across humans and animals. This has been reported to aid the high rate of transmission between humans and animals and their products. 6 Therefore, we investigated the resistance patterns and phylogenetic relatedness of E. coli isolates from humans, food-producing animals, and beef in Osun and Oyo States, Southwestern Nigeria.
Materials and Methods
Study location
Four major farm locations (Oyo State: Ogbomosho and Ibadan, Osun State: Osogbo and Ilesa) in Southwestern Nigeria were sampled between July 2015 and June 2016. These states are situated in the tropical rainforest belt of the southwestern part of Nigeria and lie approximately on latitude 7° 46′ 12″N and longitude 4° 34′ 54″E. These locations are the major city centers that serve as the food and animal depots for the residents of the two states (an approximate population of 10 million).
Sample collection, isolation, and identification of E. coli
A total of 280 samples {(fecal samples: animals (100), humans (100); beef (80)} were collected. Fresh fecal samples of apparently healthy humans and individual animals were collected in sterile universal bottles; beef samples were collected from market retailers in sterile buffered peptone water and immediately transported on ice to the Microbiology Laboratory, Department of Medical Microbiology and Parasitology, Isale Osun, Osogbo, for analysis. The samples were routinely cultured in buffered peptone water and incubated aerobically overnight at 37°C. Subcultures were made on selective plates using CHROMagar orientation medium (Mast Diagnostica GmbH, Reinfeld, Germany) and presumptive E. coli colonies selected underwent further identification tests as previously described. 6
Ethical concerns
Ethical approval of this study was obtained from the National Ethical Committee of the Osun State Ministry of Health. Verbal informed consent was obtained from all participating subjects, including custodians of animal subjects. Inclusion of participants at all categories listed in this study was based on informed consent given by the participants, while individuals who did not give their consents were excluded.
Antibiotic susceptibility testing
Antimicrobial susceptibility testing was performed using the Kirby–Bauer disc diffusion method on Mueller Hinton agar. This was carried out according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI) 11 using disks containing several different antibiotics: amikacin (AK-30 μg), amoxicillin/clavulanic acid (AMC-30 μg), ampicillin (AMP-10 μg), ampicillin/sulbactam (SAM-30 μg), cefalexin (CL-30 μg), cefazolin (KZ-30 μg), cefotaxime (CTX-30 μg), ceftazidime (CAZ-10 μg), cefoxitin (FOX-10 μg), cefuroxime (CXM-30 μg), chloramphenicol (C-30 μg), ciprofloxacin (CIP-5 μg), clindamycin (CC-10 μg), doxycycline (DO-30 μg), enrofloxacin (ENR-5 μg), kanamycin (K-5 μg), gentamicin (CN-10 μg), imipenem (IPM-10 μg), levofloxacin (LEV-5 μg), meropenem (MEM-10 μg), neomycin (N-30 μg), penicillin (1-unit), piperacillin/tazobactam (TZP-40 μg), spectinomycin (SH-25 μg), streptomycin (S-25 μg), sulfamethoxazole (RL-2 μg), sulfamethoxazole/trimethoprim (SXT-25 μg), tetracycline (TE-30 μg), ticarcillin (TIC-75 μg), tobramycin (TOB-30 μg), and trimethoprim (TMP-2.5 μg). Zone diameters of inhibition to each disk were recorded, and strains were classified as resistant or susceptible according to the criteria of CLSI. 11 E. coli ATCC 25922 was used as quality control.
ESBL confirmation by combination disk method
The isolates showing reduced susceptibility to ceftazidime or cefotaxime were tested for ESBL production by the combination disk method according to CLSI guidelines. 11 Combination disk method was performed using four disks: cefotaxime (CTX) (30 μg) and cefotaxime (30 μg) + clavulanic acid (10 μg), ceftazidime (CAZ) (30 μg), and ceftazidime (30 μg) + clavulanic acid (10 μg). A 5 mm increase in a zone diameter for the tested antimicrobial agent (CAZ or CTX) in combination with clavulanic acid versus its zone when tested alone was considered ESBL-positive. Quality control for ESBL production was performed using E. coli ATCC 25922 as negative control.
DNA extraction
The genomic DNA of the E. coli isolates was extracted by the boiling method. This was carried out by picking one colony of each isolate, suspended in 500 μL of sterile distilled water and heating at 100°C for 15 min. After centrifugation, the DNA lysate was stored at −20°C for the duration of this work. Quantification and purification of DNA samples were done using the spectrophotometric method at 260 and 280 nm as described in Ref. 12
Detection of genes
Polymerase chain reaction (PCR) for the detection of blaCTX-Ms, AAC-6-LB, TEM, and SHV genes was carried out on multidrug-resistant strains (resistant to more than two classes of antibiotics) of E. coli using DNA lysates, specific primers, and appropriate PCR conditions specific for the targeted genes. Amplification reactions were performed in a total volume of 25 μL of reaction mixture containing 5 μL of 10 × PCR buffer, 2.5 mM MgCl2, 200 mM dNTPs, and 1.25 U of Taq polymerase, and 10 pmol of each primer and 1 μL of the sample DNA. The primer details and PCR conditions with expected base pairs are shown in Table 1. Positive and negative controls were used for each of the genes.
Primer Sequences, Their Molecular Sizes, and Conditions for Amplification
PCR, polymerase chain reaction.
Phylogroup determination of the multidrug-resistant E. coli isolates
The determination of phylogenetic origin was carried out following the revised method described by Clermont et al. 16 and da Silva et al. 17 The method is based on a quadruplex PCR targeting four genes: arpA, chuA, yjaA, and TspE4.C2 (Table 2). Phylogroups were assigned based on the amplification patterns of these four genes in the following order: A (+ − − −), B1 (+ − − +), B2 (− + + +) or (− + + −) or (− + − +), and F (− + − −). Strains presenting other amplification patterns were subjected to a second PCR to discriminate among phylogroups A, C, D, and E. Prototype strains E. coli ATCC 2223, 2741, 2730, 2779, and 25922, and E. coli Nissle were used as positive controls.
Primer Sequences, Their Molecular Sizes, and Their Different Conditions of Amplification for Quadruplex Polymerase Chain Reaction
Multilocus sequence typing
Thirty of the multidrug-resistant ESBL isolates were randomly selected and subjected to MLST. According to the Warwick MLST website, seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) were designed and used for PCR. The products of the PCR amplification were sequenced by Agowa genomics and the results were submitted to the GenTel online database for processing. The allele number of each housekeeping gene was obtained, and the sequence type (ST type) of each strain was acquired.
Pulse field gel electrophoresis
The 30 ESBL E. coli isolates used for MLST analysis were also subjected to macrorestriction analysis. PFGE was performed with XbaI as a restriction enzyme according to Tenover et al. 19 The genetic profiles were compared using BioNumerics version 6.6 (Applied Maths, Sint-Martens-Latem, Belgium). PFGE types were defined with a cutoff value of 90% homology for relatedness.
Results
Distribution of the multidrug-resistant E. coli isolates by subject categories
From the total of 280 samples, 216 E. coli strains were isolated, out of which 60 were multidrug resistant: humans (23.9%), goats (50.0%), pigs (42.1%), poultry (38.9%), beef (22.6%), cattle (33.3%), and sheep (5.5%) (Table 3).
Distribution of 60 Multidrug-Resistant Escherichia coli Isolates by Subject Categories
Antibiotic resistance patterns and ESBL profile of the multiresistant E. coli isolates
Out of the 216 E. coli isolates that were obtained from the different sources, 60 (27.8%) were multiple drug resistant and were also ESBL positive. The sources included humans 23 (38.3%), goats 8 (13.3%), sheep 1 (1.7%), pigs 8 (13.3%), poultry 7 (11.7%), cattle 6 (10.0%), and beef 7 (11.7%) (Fig. 1). All the E. coli isolates (100%) were phenotypically resistant to clindamycin and penicillin. In contrast, the isolates exhibited 100% susceptibility to imipenem (Fig. 1). High rates of resistance were also experienced for ceftazidime, tobramycin, cefazolin, enrofloxacin, levofloxacin, sulfamethoxazole/trimethoprim, kanamycin, cefuroxime, piperacillin/tazobactam, ampicillin, cefalexin, streptomycin, doxycycline, neomycin, spectinomycin, amoxicillin/clavulanate, sulfamethoxazole, ampicillin/sulbactam, cefotaxime, ticarcillin, ciprofloxacin, trimethoprim, and tetracycline (50–98.3%). Low levels of resistance rates were recorded for amikacin, cefoxitin, chloramphenicol, gentamicin, and meropenem (25–41.7%).
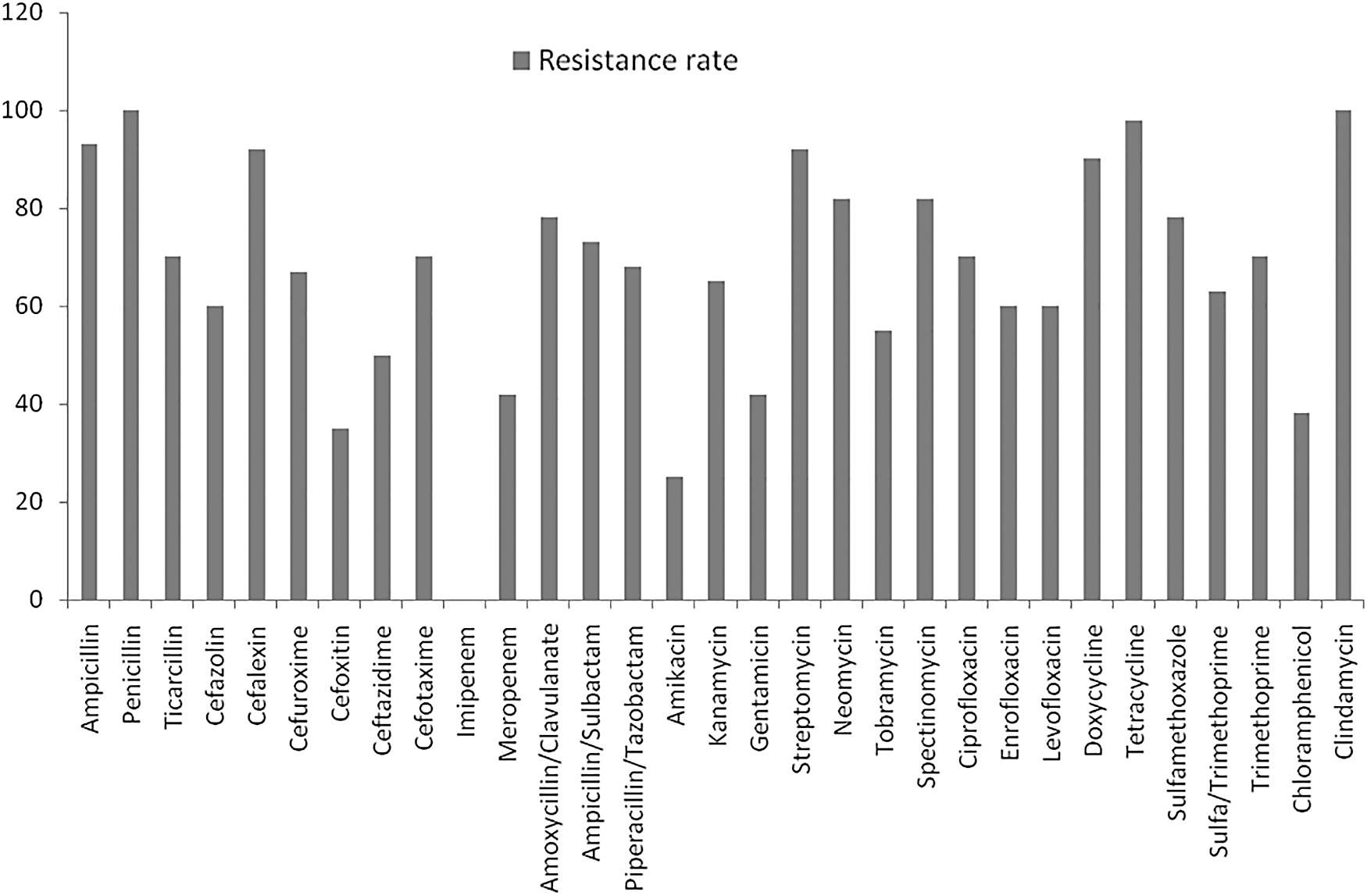
Antibiotic resistance patterns of the 60 multidrug-resistant ESBL-producing Escherichia coli isolates. ESBL, extended-spectrum beta lactamase.
The phylogroups of the multidrug-resistant ESBL E. coli isolates
The 60 ESBL E. coli isolates were divided into four phylogroups by quadruplex phylotyping: A (25%), B1 (45%), B2 (20%), and D (10%) (Fig. 2). The B1 group was the most prevalent in the human isolates and the only multidrug-resistant isolate recovered from sheep fell into this group. The second-most predominant group was A, which mostly consisted of isolates from goats, pigs, poultry, and beef. The group B2 was the third-most prevalent, being mostly populated by isolates of the human category. Isolates of humans and cattle were predominant in the least occurring phylogroup (D).

Distribution of the 60 multidrug-resistant Escherichia coli isolates in relation to their origin and phylogroups. Color images are available online.
The distribution of resistance genes among the phylogroups of the multidrug-resistant ESBL E. coli isolates
The TEM (25.0%), blaCTX-M-15 (18.3%), and AAC-6-LB (15.0%) genes were mostly associated with the B1 phylogroup (Fig. 3). This also followed for the A phylogroup (TEM—16.7%, blaCTX-M-15—11.7%, AAC-6-LB—11.7%). For the B2 group, the blaCTX-M-1 gene (16.7%) was the most prevalent followed by blaCTX-M-15, blaCTX-M-9, TEM, AAC-6-LB, and blaCTX-2 (15.0%, 13.3%, 13.3%, 13.3%, and 11.7%, respectively). All the resistance genes were found in isolates with the D phylogroup except the SHV gene.
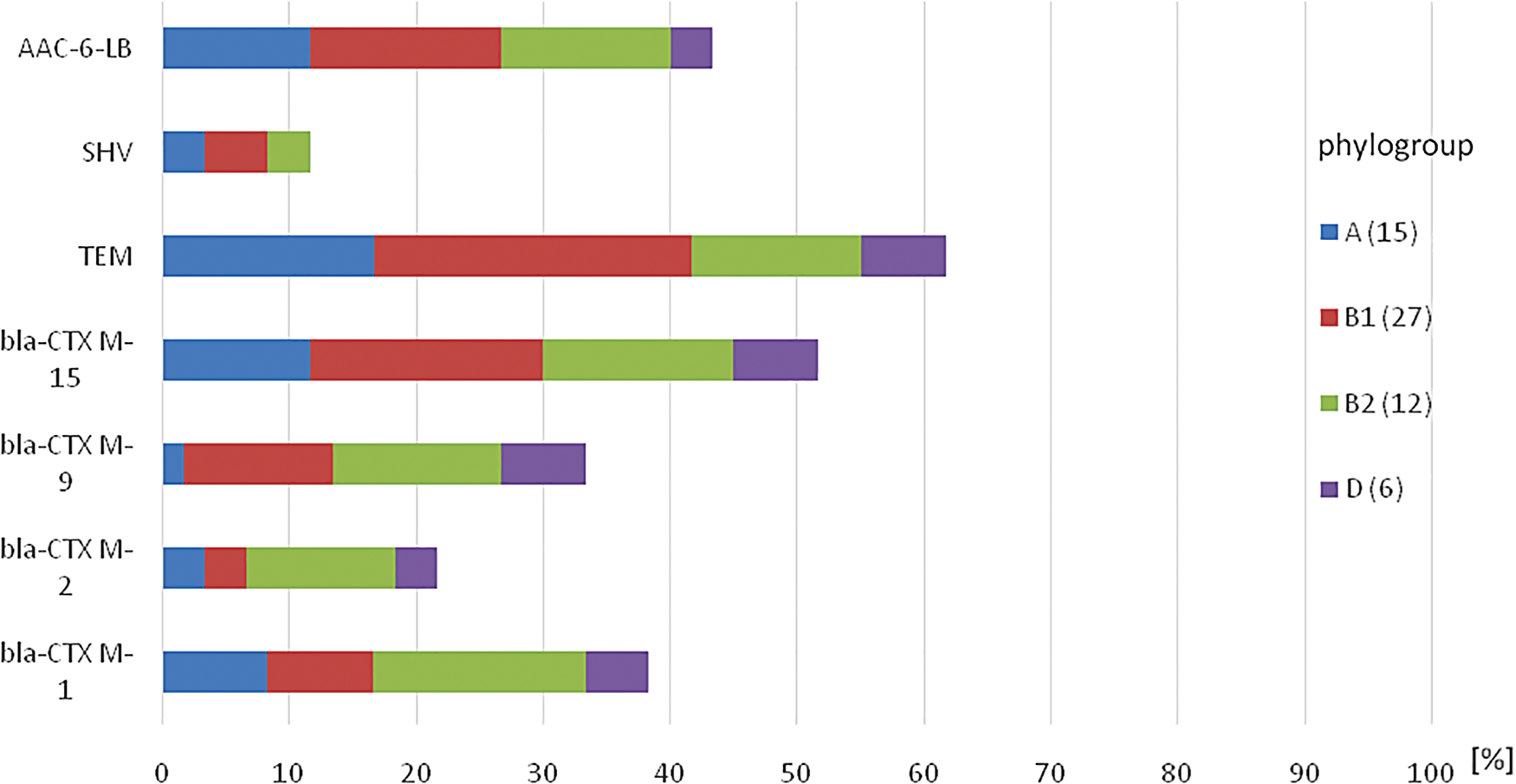
Distribution of the resistance genes among the phylogroups of the 60 multidrug-resistant Escherichia coli isolates. Color images are available online.
The distribution of the multidrug-resistant ESBL E. coli sequence types among the subject categories
Thirty multidrug-resistant ESBL isolates that were subjected to MLST were clustered into 10 sequence types: ST58, ST131, ST155, ST156, ST167, ST405, ST406, ST1056, ST1771, and ST2348 (Table 4). The ST131 (8/30, the most prevalent cluster type followed by ST405 (4/30) and ST410 (3/30). Six of the isolates (humans: 3, pig: 1, goat: 1, beef: 1) had a new ST, which has not been reported. The ST131 consisted of isolates from humans (2), pigs (2) goat (1), cattle (1), poultry (1), and beef (1). ST405 comprised isolates from cattle (2), human (1), and beef (1), while ST410 was made up of only isolates from poultry (3). Three isolates (goat: 1 and poultry: 2) belonged to the ST167 (Table 4). The remaining clonal types had an isolate each from a subject category.
The Distribution of the Multidrug-Resistant Escherichia coli Sequence Types Among the Subject Categories
The PFGE pattern of the multidrug-resistant ESBL E. coli isolates
The XbaI PFGE pattern of these 30 E. coli strains revealed 20 PFGE types (14 diverse PFGE types and 6 PFGE types having more than one isolate in each type). Each of the six PFGE types having more than one isolate in each type comprised strains from different samples and subject groups (Table 5). Isolates of each of these six PFGE types belonged to the same phylogroups and at least one resistance gene was found to be common to all the isolates. Of these six PFGE types (C1–C6), PFGE type C1 comprised isolates from humans (2) and animals (1 goat). These three isolates belonged to different sequence types but the same phylogroup and had the same resistance gene profile. The other five PFGE types were made up of isolates from different animal species. C1, C2, C5, and C6 contained isolates of the same resistance gene profiles. Isolates in C3 and C6 had the same sequence types. C4 consisted of isolates from pig (1), cattle (2), and beef (2); those from cattle and beef were of ST405, while that of pig was of a different ST. Isolates from each of the PFGE types were from different farm locations (Table 5). The dendrogram that shows the PFGE patterns and MLSTs of the multidrug-resistant E. coli isolates grouped into different clusters is presented in Fig. 4.

Dendrogram showing the PFGE patterns and MLSTs of some of the multidrug-resistant Escherichia coli isolates grouped into different clusters. C1: 4,8,27 (human, human, goat); C2: 29, 33 (pig, poultry); C3: 31, 35 (pig, poultry); C4: 30, 40, 43, 49, 51(pig, cattle, cattle, beef, beef); C5: 36, 57 (poultry, goat) not shown in the figure; C6: 37, 60 (poultry, poultry). MLST, multilocus sequence typing; PFGE, pulsed field gel electrophoresis.
Clonal Relationship of Extended-Spectrum Beta Lactamase-Producing Escherichia coli Isolates with Respect to Location, Sequence Type, Phylogroup, and Resistance Gene Profile
PFGE, pulsed field gel electrophoresis.
Discussion
The increasing rate of antimicrobial-resistant bacteria, especially E. coli, is a global health problem that affects humans, animals, and the environment. The potential interhost spread of resistant clones or even their encoding genes through direct contact or ingestion of contaminated food poses a danger to public health. 6 Although in the last decades, the number of surveys in Africa has increased, available information is still scanty in many countries on clonal lineages that harbor resistance determinants. We investigated the epidemiology of multidrug-resistant ESBL E. coli from humans, food-producing animals, and beef using quadruplex phylotyping and MLST. This approach was geared toward knowing the spread of resistance genes in animals and man along the food chain.
Sixty multidrug-resistant ESBL E. coli strains were isolated from various human and animal fecal samples, as well as beef. E. coli strains are now classified into eight phylogroups, A, B1, B2, C, D, E, F, and Escherichia cryptic clade I. 20 Studies have shown that isolates belonging to the A and B1 phylogroups are commensals, while those that belong to the B2 and D groups are the extraintestinal pathogenic strains.20–22 Most of the human isolates in this study belonged to the group B1 (15/23) contrary to the findings of Maciuca et al., 23 where the majority of their human isolates fell into the group B2 (15/25). The reason for this is not far-fetched as the human isolates were all clinical, while our own isolates were from fecal samples of apparently healthy individuals.
Six (26%) out of the 23 human isolates belonged to the potentially pathogenic phylogroups B2 and D. This could have a clinical and epidemiological significance in terms of disease outbreak and human-to-human transmission of infections. The isolates from the food-producing animals and beef were majorly clustered into the groups A and B1, although few of them were found in the B2 group and very few in the group D. This goes to stress that these isolates were mostly commensals, which as a result of antibiotic exposure and other environmental and genetic factors may revert to being pathogenic. 24
The high prevalence of the commensals among these multidrug-resistant isolates is, however, not surprising as it has been shown that isolates of the B2 group (pathogenic) are usually more susceptible than those of the commensal groups 25 although they possess enhanced molecularly inferred virulence capacity. The isolates from the food-producing animals and beef that were of the B2 and D phylogroups also have clinical and epidemiological implications as they could be transferred along the food chain. These high rates of resistance exhibited by the commensal isolates may be a result of horizontal acquisition or random point mutations in their genetic makeup overtime. 24 A judicious use of antibiotics must therefore be ensured to prevent the cross-selection and coselection of resistant E. coli strains, which will also curtail their spread in the food chain.
Among these phylogroups, the TEM-gene was most prevalent followed by the blaCTX-M-15 and AAC-6-LB genes. blaCTX-M-1, blaCTX-M-9, blaCTX-M-2, and SHV genes were also found in descending order of prevalence. Although studies26–28 have shown that group B2 is mostly susceptible to antibiotics, our study found that isolates in this group carried the highest prevalence of resistance genes followed by those in the B1 phylogroup. A similar finding was reported by Vading et al. 29 This may be due to the currently prevailing multiple resistance phenotypes carried by most E. coli strains both in the community and hospital settings owing to the abuse of antibiotics in livestock and human medicines. It must be noted again that all the 60 isolates were multidrug resistant.
The blaCTX-M ESBL genes in combination with the TEM gene and the AAC-6-LB (gentamicin resistance) gene were common among the isolate phylogroups in this study, which is similar to the reports of Valentin et al. 30 The ESBL genes were broadly distributed within the isolates of the four phylogroups from all the subject categories. This may reflect the interaction of isolates among all these subject groups and may explain the transmission of these isolates along the animal-to- human food chain although a thorough molecular investigation should be channeled into this. These resistance traits may also reflect the ability of the various phylogroups to colonize and adapt to a wide range of animal host species.
The MLST of 30 of the multidrug-resistant ESBL E. coli isolates (randomly selected) revealed 10 sequence types, with ST131 being the most prevalent followed by ST405, ST410, and ST167. The ST131 consisted of isolates from humans, pigs, goats, cattle, poultry, and beef. This also applied for isolates in the ST405 PFGE type; no human isolates were, however, found in the STs 410 and 167, which were majorly populated with isolates from poultry. The shared sequence types by some of the isolates of the different subject categories imply a single parental origin, which again informs the likely circulation of the same clones within the subject groups. It may therefore be safe to infer an interhost transmission of the clones through different transfer routes.
Isolates from cattle shared similar characteristics with those from beef: four isolates (two from cattle and two from beef) had similar sequence type (ST 405), PGFE pattern (C4), and phylogroup (D). This suggests likely contamination along cattle slaughtering, processing, and beef marketing chain. Phylogroup D is a potentially pathogenic phylogroup associated with extraintestinal infections. Consumption of such contaminated beef could lead to clinical infections in consumers. There is a need for improved hygiene in beef/meat processing. ST131 has a high virulence potential, a broad host range, and multidrug resistance phenotypes. 20 Isolates of this clone harbor the blaCTX-M genes, and even now, the carbapenemase resistance genes, 20 which make infections caused by them to defy treatments both in humans and food-producing animals.
Day et al. 31 also found higher prevalence rates of ST131 and ST405, including other STs such as 10, 58, and 117 among isolates from humans, animals, and food in three European countries. They, however, found a high diversity of sequence types among their isolates from different sources, in contrast to our study where some isolates from different sources shared the same STs. The ST410 is distributed globally; isolates from meat and clinical samples having this sequence were reported in Spain to harbor the blaCTX-M genes and virulence genes.32,33 This clone as well as ST167 has been found in interspecies circulation, including food, which is also a potential risk of transmission vehicle to consumers if contaminated.34,35 Other STs such as 58 were also reported in Lebanese cattle.36,37
The PFGE results also revealed a clonal relationship among isolates of different subjects, suggesting an epidemiological overlap of the E. coli strains. 30 Six PFGE types were discovered that comprised strains from heterogeneous and homogeneous subjects. One of these PFGE types was shared by isolates of human and animal subjects, which were of the same phylogroup and resistance gene profile but of different STs. Similar studies that found an overlap of human and animal isolates in terms of phylogroups and antibiotic resistance profiles include Hammerum et al., 10 Valentin et al., 30 and Dohmen et al. 37 In another similar study conducted by Pietsch et al., 38 clonal relationship was also noticed in a few E. coli isolates from humans and food animals hinting toward an ancestral origin.
The remaining PFGE types had isolates from different animal subjects and beef. This suggests an interhost transmission of antibiotic resistance-bearing isolates and that transfer of ESBL E. coli isolates could be through multiple routes, including the food chain as was also suggested by Ojer-Usoz et al. 39 Isolates from each of the PFGE types were from different farm locations. A geographical spread of the PFGE types might have occurred. This observation was also documented by Irrgang et al. 40 Apart from the fact that these strains were isolated within the same time period and from the same geographical zone, it is also important to note that they harbored almost the same resistance profiles. It is therefore safe to suggest that there is a clonal spread of resistant E. coli strains among animal and human subjects as well as beef in our own environment.
Although we have been able to find a clonal spread of these isolates by MLST and PFGE, there is, however, a need to establish the routes and direction of transmission of the strains.
Conclusion
This study reports the detection of blaCTX-M-type ESBLs, TEM, SHV, as well as AAC-6-LB genes in multidrug-resistant E. coli isolates from four different phylogenetic groups in humans, food-producing animals, and beef in Southwest Nigeria. This presents a major concern because of the possibility of transfer of these resistance gene-bearing strains by direct contact or through animal product consumption and even from human to human. Similar PFGE types of isolates were found in human, animal, as well as beef samples in the same sampling period. This may indicate an animal origin of the isolates and cross-transmission between hosts.
A whole-genome sequence analysis is therefore necessary to detect the actual trends of the transmission. There is also a need for continuous surveillance of both human and veterinary antibiotic-resistant isolates to curtail the spread of existing clonal groups and the emergence of new clonal clusters.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work has been supported by the DAAD funding program “ID: Re-invitation program for former scholarship holder 2016” and by the grant Innoprofile Transfer 03 IPT 611X funded by the Federal Ministry of Education and Research (BMBF, Germany).