
Abstract
The increasing spread of carbapenem resistance is a serious global public health concern that negatively affects human and animal health. In this study we characterized the carbapenemase production in gram-negative bacteria isolated from different meat and meat products in Egypt. Phenotypic and genotypic susceptibility testing were investigated. Two Enterobacter cloacae complex strains, isolated from kofta and beef burger, and one Pseudomonas aeruginosa isolated from minced meat, were found to harbor VIM-1 and VIM-2, respectively. These isolates showed multidrug resistance phenotype. The phenotypic carbapenemase production was confirmed with Carba NP test in addition to modified Hodge test, modified carbapenem inactivation method, and ethylenediaminetetraacetic acid inhibition test. The blaVIM-1 gene in both non-clonally related E. cloacae complex strains was part of a class 1 integron that also carried other resistance gene cassettes such as aacA7, dfrA1, ΔaadA, and smr. This integron was uncommonly disrupted by the insertion sequence ISPa21, located on a self-conjugative plasmid of either the A/C or HI2 incompatibility group with a size of >93 kb. The blaVIM-2 gene was identified within a class 1 integron, followed downstream by resistance genes aadB and blaOXA-10. The transfer of blaVIM-2 gene from P. aeruginosa failed, suggesting that this gene was located on the chromosome. Further studies are needed to screen the dissemination of carbapenemase-producing bacteria in both the environment and food chain.
Introduction
The alarming emergence and spread of antibiotic resistance is a serious threat to global public health. The overuse and misuse of antibiotics in food animals either for disease treatment or growth promotion allows antimicrobial-resistant bacteria and antimicrobial resistance genes to spread from food animals to humans.1,2 Meat and dairy products are considered as potential vectors for the transfer of multidrug-resistant (MDR) bacteria between animals and humans. 3 Among the most clinically significant MDR bacteria are carbapenemase-producing Enterobacterales (CRE). Carbapenemases are classified into three classes of β-lactamases: Ambler class A (KPC, GES SME, IMI, and NMC), Ambler class B (VIM, IMP, SPM, GIM, and NDM) that are zinc-dependent metallo-β-lactamases (MBLs) strongly hydrolyzing all β-lactams except aztreonam, and Ambler class D (OXA-type) weakly hydrolyzing carbapenems.4,5
The VIM-type MBLs were first identified in Pseudomonas aeruginosa in Europe. VIM-1 was identified in a carbapenem-resistant P. aeruginosa clinical isolate from a patient at Verona university hospital, Italy and VIM-2 from a P. aeruginosa isolate in a blood culture of a neutropenic patient in Marseilles, France.4,6 CRE infections are associated with increased morbidity and mortality owing to the limited availability of treatment options and the lack of development of new antimicrobial agents. 7 Recently, Khalifa et al. reported a high incidence (50.8%) of carbapenem-resistant gram-negative bacteria in Egypt, with OXA-48-like, NDM-1, and VIM being detected in 49.2%, 47.7%, and 26.2% of carbapenemase-positive isolates, respectively. 8 In addition, various types of carbapenemase producers with numerous carbapenemase-encoding genes (i.e., NDM-1, NDM-5, VIM-2, VIM-24, and OXA-181) were identified from a university teaching hospital in Egypt. 9 In 2016, NDM-4- and NDM-5-producing Klebsiella pneumoniae coinfection in an Egyptian 6-month-old infant was reported. 10 Several carbapenemases have been reported among Enterobacterales in Africa such as NDM-5 in Egypt and Algeria, OXA-181 in São Tomé and Príncipe, Angola, and Nigeria, and VIM-2 in South Africa, Ivory Coast, and Algeria.9,10 IMP was identified in Egypt, Morocco, Tunisia, and Tanzania.11,12
Moreover, the dissemination of carbapenem resistance has been mainly recognized in the human sector and is poorly recognized in food and food products. Therefore, our study focused on the molecular mechanisms underlying carbapenem resistance and their prevalence among gram-negative bacteria of food origin.
Materials and Methods
Bacterial isolates
A total of 110 different meat and meat product samples were randomly collected between June and December 2017 from different supermarkets, slaughterhouses, and butchers' shops in four different cities (Qena, Nag Hammadi, Luxor, and Esna) located in Qena (25°41′0′′ N, 32°39′0′′ E) and Luxor (26°10′12′′ N 32°43′38″ E) Governorates, Upper Egypt. The samples included fresh beef (n = 21), frozen beef (n = 10), mutton (n = 15), minced meat (n = 12), burger (n = 13), sausage (n = 16), luncheon (n = 6), kofta (n = 13), and pastirma (n = 4). The samples (25 g each) were homogenized and enriched in buffered peptone water (225 mL) for 24 hr at 37°C with shaking, followed by direct spreading on MacConkey agar supplemented with 2 mg/L meropenem (MEM, sigma). Colonies of different morphology, size, and color from each plate were selected and purified.
Bacterial identification and antibiotic susceptibility testing
The isolates were identified using the API 20E System (BioMerieux) and further confirmed by 16S rRNA gene sequence analysis. 13 The isolates were subjected to susceptibility testing against 15 antimicrobial agents amoxicillin–clavulanic acid (20/10 μg), aztreonam (30 μg), chloramphenicol (30 μg), ciprofloxacin (5 μg), ceftriaxone (30 μg), cefotaxime (30 μg), doripenem (10 μg), cefoxitin (30 μg), gentamicin (10 μg), imipenem (10 μg), kanamycin (30 μg), MEM (10 μg), nalidixic acid (30 μg), tetracycline (30 μg), and ampicillin (10 μg) by the Kirby–Bauer disk diffusion assay according to the Clinical and Laboratory Standards Institute (CLSI) protocols, interpretive criteria, and reference strains. 14 The disks were purchased from Eiken Chemical Co. Ltd. (Tokyo, Japan). Strains exhibiting resistance to three or more antibiotic classes were classified as MDR. The minimum inhibitory concentrations (MICs) of selected antimicrobials were determined using the broth microdilution method. The results were interpreted according to the CLSI guidelines. The reference strain Escherichia coli ATCC 25922 was included as a quality control.
Phenotypic detection of carbapenemase activity
The carbapenemase activity was tested by modified Hodge test (MHT) against MEM using E. coli ATCC 25922 as the indicator organism. 14 E. coli ATCC 25922 (0.5 McFarland suspension) was streaked as lawn on to a Mueller–Hinton agar (MHA) plate. A 10 μg MEM disk was placed in the center of the test area. Each organism was subsequently streaked in a straight line from the disk to the edge of the plate. The plate was incubated overnight at 35 ± 2°C in aerobic conditions for 16–24 hr. MHT-positive result was indicated by the presence of a clover leaf-like indentation of the E. coli 25922 growing along the test organism growth streak within the disk diffusion zone. MBL production was also tested with modified carbapenem inactivation method (mCIM) and ethylenediaminetetraacetic acid (EDTA) inhibition test. 14 This combination phenotypic detection method was performed using MEM and MEM-EDTA. A growth zone difference >5 mm confirmed the presence of MBL. 15 mCIM was performed by incubating a 10 μg MEM disk with the test organism. The MEM disk was then removed and placed on the MHA plate previously inoculated with 0.5 MacFarland suspension of the MEM-susceptible E. coli strain ATCC 25922, which was then incubated in an inverted position for 18–24 hr at 35 ± 2°C in aerobic conditions. Inhibition zone diameter ≤15 mm around the MEM disk was considered a positive result (indicating the presence of carbapenemase production). The phenotypic carbapenemase production was also confirmed with Carba NP test. 16
Bacterial DNA preparation
DNA was prepared using boiled lysates, as previously with some modifications. 3 In brief, the bacterial isolates were grown in lysogeny broth (LB) overnight. These cultures were centrifuged at 13,000 rpm for 2 min. The pellets were resuspended in 1 mL of deionized water (dH2O), boiled at 95°C for 10 min, cooled in ice, and centrifuged. The supernatant was used as the DNA template. DNA template was stored at −20°C until further use.
Detection of acquired carbapenemase-encoding genes
All isolates were screened by PCR for the following carbapenemase encoding genes: blaVIM, blaIMP, blaKPC, blaNDM, blaOXA-48-like, blaBIC, blaAIM, blaSPM, blaDIM, blaGIM, and blaSIM as described previously. 17 The reaction products were subjected to electrophoresis in a 2% agarose gel, stained with ethidium bromide, and visualized under ultraviolet light. Then, PCR products were then treated with ExoSAP-IT reagent (Applied Biosystems™) by removing unused primers and nucleotides according to the manufacturer's instructions.
Molecular characterization of other antimicrobial-resistance genes
PCR and DNA sequencing were used to detect extended-spectrum β-lactamases (ESBLs) (blaTEM, blaSHV, blaOXA, blaCTX-M, and blaCMY), plasmid-mediated quinolone-resistance determinants [qnrA, qnrB, qnrS, qepA and oqxAB, qnrC, qnrD, and aac(6′)-Ib-cr], a plasmid-mediated colistin-resistance gene (mcr-1 mcr-3, mcr-3, mcr-4, mcr-5, mcr-7 and mcr-8), and 16S rRNA methylases (armA, rmtB, npmA, rmtA, rmtC, and rmtD).18–25
PCR mapping and characterization of blaVIM containing class 1 integron
As the blaVIM gene is usually a part of gene cassettes included in class 1 integrons, the gene cassettes inserted into class 1 integron were amplified using primers designed based on the 5′-conserved segment (CS) and the 3′-CS. Primers used in PCR mapping and sequencing are listed in Table 1.22–25
Oligonucleotides Used in This Study
CS, conserved segment.
Conjugation and transformation experiments
To study the transferability of the blaVIM −1 and blaVIM-2 genes, conjugation experiments were performed by the filter mating method. 26 The azide-resistant E. coli strain J53 was used as the recipient. Both donor and recipient strains were cultured separately in LB broth (5 mL). After incubation, the donor and recipient strains were mixed in a ratio of 1:9 (donor:recipient) and centrifuged; the supernatant was removed, and the pellets were resuspended in 200 μL LB broth, which was plated on a conjugation filter on a LB agar plate. The plates were incubated for 3–4 hr at 37°C. Transconjugants were selected on LB agar plates supplemented with ampicillin (100 mg/L) and sodium azide (150 mg/L). After several attempts, conjugation failed to transfer the blaVIM-1 gene from Enterobacter cloacae complex M37-3 to the recipient. Therefore, the plasmid extracted from E. cloacae complex M37-3 culture by the alkaline lysis method was electroporated into E. coli DH10B with selection on LB plates containing ampicillin (100 mg/L). Conjugation was repeated with the E. coli DH10B electrotransformant as the donor and an azide-resistant E. coli J53 strain as the recipient. Transconjugants was confirmed by PCR targeting the VIM gene, antimicrobial susceptibility testing, and plasmid analysis.
Plasmid isolation, plasmid analysis
Plasmids were isolated from VIM-positive isolates using the alkaline lysis method. 27 The plasmid size was evaluated by agarose gel electrophoresis using E. coli O157:H7 as a size control, which harbors plasmids with sizes of 93 and 114 kb. 28 The PCR-based replicon typing method was used to identify the incompatibility group of VIM-positive plasmids as previously reported. 29 The major plasmid incompatibility groups among Enterobacterales including FIA, FIB, FIC, HI1, HI2, I1-Igamma, L/M, N, P, W, T, A/C, K, B/O, X, Y, F, and FIIA were recognized by five multiplex and three simplex PCRs.
Clonality evaluation
The epidemiological relatedness of the E. cloacae complex isolates was evaluated by pulsed-field gel electrophoresis (PFGE) analysis, enterobacterial repetitive intergenic consensus (ERIC) sequence PCR analysis, and multilocus sequence typing (MLST). The MLST was performed according to the Enterobacter and Pseudomonas sequence typing websites. Seven housekeeping genes each were used for E. cloacae (dnaA, fusA, gyrB, leuS, pyrG, rplB, and rpoB) and P. aeruginosa (acsA, aroE, guaA, mutL, nuoD, ppsA, and trpE) MLST analyses. ERIC PCR was performed using the ERIC1 (5′-ATGTAAGCTCCTGGGGATTCAC-3′) and ERIC2 (5′-AAGTAAGTACTGGGGTGAGCG-3′) primers. 25
Analysis of the sequence data
A similarity search was carried out using the BLAST program available at the National Center for Biotechnology Information BLAST homepage.
Results and Discussion
The emergence and worldwide spread of CRE is of great concern to public health services. We have described the detection of VIM-MBL enzymes in fresh meat product samples from Egypt for the first time. A total of 148 MEM-resistant isolates (MIC >2 mg/L) were selectively isolated on MacConkey agar supplemented with meropenem from food samples (meat and meat products) collected from different supermarkets, slaughterhouses, and butchers' shops in Egypt in 2017. The acquired carbapenemase-encoding genes, such as blaVIM, blaIMP, blaKPC, blaNDM, blaOXA-48-like, blaBIC, blaAIM, blaSPM, blaDIM, blaGIM, and blaSIM were screened among all isolates. The VIM-MBL gene was detected in only 3 of the 148 MEM-resistant strains: 2 E. cloacae complex strains isolated from kofta and beef burger harbored VIM-1, and 1 P. aeruginosa strain isolated from minced meat harbored VIM-2. Other carbapenemase-encoding genes were not detected and the overall prevalence rate of carbapenemase-producing gram-negative bacteria from meat and meat products in this area of Upper Egypt remains low. Abdallah et al. reported the identification of 12 NDM-producing Klebsiella spp. among 106 CRE (12/106; 11.32%) isolated from retail chicken meat purchased in Zagazig, lower Egypt in 2013. 30 In addition, a study conducted on 890 Salmonella isolates collected from retail chicken carcasses in China identified that 25.0% (24/96) of the ESBL-producing isolates carried blaVIM and 10.4% (10/96) carried the class A carbapenemase KPC. 31 Another study conducted in Guangzhou, China identified one E. coli strain, isolated from a chicken wing sample in 2014, carrying mcr-1 and NDM-9. 32 The MEM-resistant isolates showed an MDR phenotype. The antibiotic susceptibility testing indicated that both E. cloacae complex strains were resistant to amoxicillin–clavulanic acid, ampicillin, cefoperazone, cefotaxime, ceftriaxone, doripenem, imipenem, kanamycin, meropenem, and tetracycline but were still sensitive to colistin, whereas P. aeruginosa was resistant to all antimicrobials tested except colistin (Table 2). The MICs were determined using the broth microdilution method (Table 2). Phenotypic carbapenemase activity was confirmed with a Carba NP and MHT. All VIM-positive isolates were reported positive in the mCIM and the EDTA inhibition test indicating the production of MBL. The dissemination of enterobacterial isolates with acquired carbapenemases through the food chain may indicate further spread of VIM producers in the food environment. It further indicates that VIM producers may be a true One-Health issue disseminating in the environment and then in humans through the food chain.
Antibiotic Susceptibility Testing of Various Antibiotics Against VIM-Positive Isolates
Resistance to the antibiotics are given in bold.
The antibiotic resistance profile was determined using the Kirby–Bauer disk diffusion assay.
The MIC was determined using broth microdilution technique.
AMC, amoxicillin-clavulanate; AMP, ampicillin; AZT, aztreonam; CFP, cefoperazone; CHL, chloramphenicol; CIP, ciprofloxacin; CRX, ceftriaxone; CST, colistin; CTX, cefotaxime; DOR, doripenem; FOX, cefoxitin; GEN, gentamicin; IPM, imipenem; KAN, kanamycin; MEM, meropenem; NAL, nalidixic acid; NOR, norfloxacin; TET, tetracycline; MIC, minimum inhibitory concentration.
PCR mapping and sequencing of the class 1 integron in both E. cloacae complex strains revealed that the blaVIM-1 cassette was located as the first gene cassette directly downstream of the integrase gene intI1 (Fig. 1A). The class 1 integron also carried other resistance gene cassettes such as aacA7 [coding for aminoglycoside (6′)-N-acetyltransferase type Ib], dfrA1 (coding for dihydrofolate reductase I), ΔaadA (with 5′ side 76 bp deletion; nonfunctional owing to fusion on 5′-end; fusion of core site/aadA), and smr (coding for a small multidrug resistance protein) (Fig. 1A). A complete ISPa21 insertion sequence containing the tnpA gene flanked by two 13-bp inverted repeats, IRR (5′-ATGGACTCTCCCC-3′) and IRL (5′-GGGGAGAGTCCAT-3′), was located directly upstream of the 3′ CS conserved region of the integron. The association of ISPa21 with different antibiotic resistance genes may play an important role in further dissemination and mobilization of these genes among other pathogens. 33 Of interest, the same class 1 integron carrying blaVIM-1 in both E. cloacae complex strains was previously reported in clinical MDR P. aeruginosa strains of Egypt. 33 The sequence of this integron obtained from E. cloacae complex strains was similar to that of In416 previously reported in E. cloacae and K. pneumoniae from Italy and K. pneumoniae from France.34,35 In case of In416, the dfrI cassette was fused with the aadA1 gene by an 81 bp deletion, involving part of the attC recombination site of the dfrI cassette and the 5′ end of aadA1. 26 The blaVIM-2 gene in P. aeruginosa was identified within a class 1 integron, followed downstream by two resistance gene cassettes: aadB, and blaOXA-10 (Fig. 1B). To best of our knowledge, this is the first report showing such an unusual arrangement of gene cassettes, blaVIM-1, aacA7, dfrA1, ΔaadA, and smr gene cassettes disrupted by a complete ISPa21, in MDR E. cloacae of food origin.
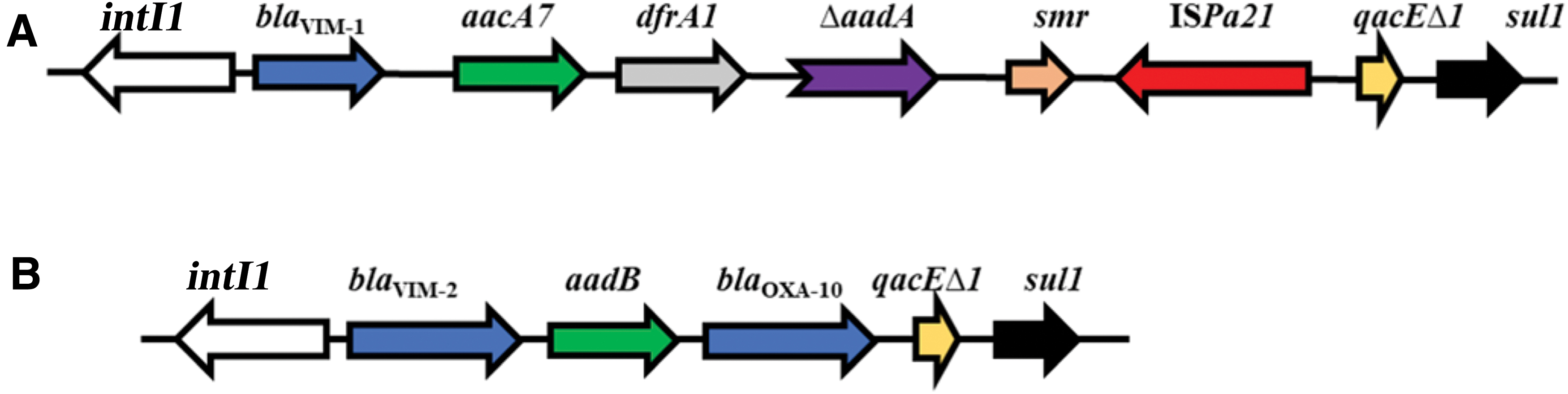
Schematic mapping of class 1 integron detected among Enterobacter cloacae complex
Conjugation and transformation experiments were performed to study the transferability of the blaVIM genes. The results showed that the blaVIM-1 harboring plasmid from E. cloacae could be transferred by conjugation to an azide-resistant E. coli J53 strain, indicating that it was carried by a self-transferable plasmid (MIC for MEM: 16 mg/L). After several attempts, conjugation failed to transfer the blaVIM-1 gene from E. cloacae M37-3. When conjugation was repeated using the E. coli DH10B electrotransformant as the donor, the plasmid was found to be conjugative and successfully transferred to an azide-resistant E. coli J53 strain (MIC for MEM: 16 mg/L). Several attempts to transfer the blaVIM-2 gene from P. aeruginosa M42-2 by conjugation and transformation failed, suggesting the chromosomal location of the blaVIM-2 gene.
PCR-based replicon typing revealed that the blaVIM-1 gene in both E. cloacae complex strains were located on a conjugative plasmid belonging to either A/C or HI2 with a size of >93 kb. The first A/C plasmid encoding VIM-1-MBL was reported in K. pneumoniae isolates from neonates in Naples. 36 PFGE patterns and ERIC PCR analysis identified of both E. cloacae complex strains showed different patterns corresponding to different sequence types (STs) because of their genetic diversity. MLST analysis revealed that E. cloacae complex M37-3 belongs to ST279, which has been previously reported in blaCTX-M-15-positive isolate in China. 37 A novel ST shown by E. cloacae M31-1 isolate M42-2 belonged to ST235, which is spread extensively throughout Belarus, Kazakhstan, and Russia, Malaysia, Thailand, Sri Lanka, Korea, and France.38–40
Conclusion
In this study, we reported the detection of carbapenemase encoding genes in gram-negative bacteria isolated from meat products in Egypt. The detection of carbapenemase producers in meat and meat products has important implications for food safety as retail meat and meat products could serve as a vehicle for these resistant bacteria, which could be transferred between animals and humans through the food chain. Further studies are needed to screen the dissemination of carbapenemase-producing bacteria in the environment and food chain.
Authors' Contributions
M.S. and Ta.S. designed and directed the study. M.S. performed bacterial identification, antimicrobial susceptibility, characterization of resistance genes, conjugation, transformation, plasmid analysis, and clonal relatedness. M.S. analyzed the results and drafted the article. Ta.S., H.N., A.M.S., and To.S. contributed to the analysis of the results and the writing of the article. All authors read and approved the final version of the article.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
M.S. was supported by a scholarship (Egypt–Japan partnership) from the Egyptian government. A.M.S. was supported by a fellowship (153532) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan.