
Abstract
In this study, we report a high incidence of New Delhi metallo-β-lactamase (NDM)-producing and ampicillin-catabolizing bacteria within carbapenem-resistant bacterial populations in the waters of two important rivers, Mahananda and Karala, bisecting two most populous towns, Siliguri and Jalpaiguri, respectively, in the northern West Bengal, India. Isolates producing NDM belonged to four genera, Acinetobacter, Escherichia, Proteus, and Pseudomonas; among which few were phylogenetically determined as putatively novel species. Class 1 integrons with the frequent presence of aadA and aac(6′)-Ib gene cassettes in 50% of NDM-bearing isolates are indicative of possible selective pressures generated out of unregulated use of streptomycin, in agriculture practiced by the cultivators and tea planters living in locales drained by these two rivers, in their up- and downstream, and amikacin in the most crowded government-sponsored “sadar” and district hospitals of Siliguri and Jalpaiguri. NDM-delivering bacteria in rivers have genuine consequences for city inhabitants who are dependent on public water and sanitation facilities. Standard reconnaissance of antibiotic resistance, consolidating ecological sampling just as the assessment of clinical isolates, should be set up as a need.
Introduction
The high incidence of antibiotic-resistant bacterial community is a rising ecological fret with genuine consequences for general well-being. The part played by the environment, in the evolution and dissemination of antibiotic-resistant bacteria, is progressively picking up consideration. Several studies have shown abundances of antibiotic residues, antibiotic-resistant bacteria, and antibiotic-resistance genes (ARGs) in different environments, especially water systems that are affected by effluent from sewage, horticultural and pharmaceutical industries.1–3 Rivers, which are contaminated with community sewage, livestock wastewaters, and clinic effluents, are continuously enriched with antibiotic-resistant bacteria. The river is, therefore, viewed as a hot spot for the procurement and spread of ARGs to different ecological niches. 1 The occurrence of genes coding for extended-spectrum β-lactamases and metallo-β-lactamases (MBLs) in the aquatic ecosystem has significant consequences for individuals who are dependent on natural water sources. The spread of resistance in an ecological framework happens to correspond to the absence of sanitation, which could be related to the South Asian condition with expanded range β-lactamases as well as carbapenemase-producing Enterobacteriaceae. 3
Carbapenem resistance, primarily among Gram-negative pathogens, is a global medical issue, and in particular, when reinforced by resistance-gene-transfer potential across several bacterial genera, the problem in treating any bacterial infection gets worsened. Since the discovery of a novel gene coding for New Delhi MBL-1 (NDM-1) in a pathogenic bacterium, India has become the point of convergence of worldwide research interest on antimicrobial resistance. 4
The molecular basis of the transmission of blaNDM-1-positive Enterobacteriaceae has indicated ecological and evolutionary implication. 5 The dissemination rate of NDM-producing bacteria in a human environment has significant implication for individuals living in the city who are reliant on open water and sanitation facilities.6,7 There has been a pressing demand to assess the spread of NDM producers among both the clinical isolates and pathogens procured from the surface-water network. 8
In this study, we have investigated the incidence of NDM-producing bacteria occurring within carbapenem-resistant bacterial populations in samples of two significant rivers, Mahananda and Karala, bisecting two most populous towns, Siliguri and Jalpaiguri, respectively, in the northern West Bengal, India. The NDM-producing isolates were also tested for their ability to consume ampicillin as sole carbon and energy source.
Materials and Methods
Sampling stations
After examination of the topographic map of Karala and Mahananda river of northern West Bengal, three sampling stations were selected individually on both the rivers (upstream, midstream, and downstream). Sampling stations of each river are shown in Table 1.
Sample Collection Details of Two Rivers of North Bengal Mahananda and Karala
Sample collection and processing
River water samples were collected during premonsoon (March and April) and postmonsoon (November and December) months. Three grab samples, from each sampling site, were collected from the left, right, and middle of the river. All three samples were analyzed as a single sample. Altogether, 24 composite water samples (6 sampling sites [3 sites on individual river × 2 rivers] × 1 composite sample from each site × 4 times sampling) were collected.
For sampling, sterilized water bottles were used. The bottles were opened underwater, rinsed thoroughly with the river water even it was precleaned, and were half-filled by opening and closing the bottles underneath the water. The minimum elapsed time between collection and analysis in no case did exceed 30 hr for samples held at 4°C, after reaching the laboratory. Serially diluted water samples were plated on Mueller–Hinton Agar (MHA) plates, supplemented with imipenem (20 mg/L). Plates were incubated at 37°C for 24 hr.
Laboratory studies designed to culture multiple-antibiotic-resistant bacteria had a formal approval from the Departmental Review committee and was conducted in a BSL-2 containment.
Screening of MBL-positive strains from imipenem-resistant colonies
Replica plating method was employed for phenotypic screening of MBL-positive isolates. Discrete colonies evident on imipenem (20 μg/mL) containing MHA plates were numbered serially (1,2,3,…n). Approximately, 50 ± 10 random numbers (of >150 discrete colonies) were generated using a research Randomizer tool. Colonies corresponding to the random numbers were then picked up with the help of sterile loop and transferred to the respective grids of the first-generation master plates of MHA plates containing imipenem (20 μg/mL). The first-generation master plates were then replicated on the ethylenediaminetetraacetic acid (EDTA)-impregnated (200 μL of 50 mM EDTA solution was used) imipenem (20 μg/mL) plates and incubated at 37°C for 24 hr. Isolates that did not show any growth on EDTA-impregnated plates after proper incubation were considered as potential MBL producers.
Genotypic confirmation of MBL-positive phenotype through multiplex PCR
Multiplex PCR was used for further confirmation of MBL-positive phenotype of the isolates that did not grow on EDTA-impregnated imipenem plates. The PCR methodology followed in this study was described earlier by Poirel et al. 9 The sequences of the primers used in this study to amplify blaIMP, blaVIM, blaNDM, blaSPM, blaAIM, blaDIM, blaGIM, blaSIM, blaKPC, blaBIC, and blaOXA are shown in Supplementary Table S1.
Determination of minimum inhibitory concentration values
Minimum inhibitory concentrations (MICs) of β-lactams, penicillins+β-lactamase inhibitors, cephalosporins, carbapenems, fluoroquinolones, aminoglycosides, trimethoprim, tetracycline, and chloramphenicol were determined by a standard broth macrodilution method, in Mueller–Hinton broth, following Clinical and Laboratory Standards Institute (CLSI) guidelines. 10 The inoculum used per experiment was 104 to 105 cfu/mL.
Determination of antimicrobial susceptibility of MBL-positive isolates
MHA being nonselective, nondifferential, containing starch that absorbs toxins released from bacteria to prevent interference with antibiotics and also facilitate diffusion of antibiotics, was employed for the determination of antimicrobial susceptibility following the Kirby–Bauer disk-diffusion method. Zone diameter determining resistant, intermediate, and sensitive results for the isolates (identified as Acinetobacter sp. or Escherichia sp. or Pseudomonas sp. or Proteus sp.) was concluded from performance standards obtained from CLSI. 10 Commercially available antibiotic disks (HiMedia Co., India) were used.
Viability and growth of MBL-positive isolates in diluted 0.1X and 0.01X Luria broth
The inoculum was prepared by transferring a single colony of 24 hr-old cultures of MBL-positive isolate into 10 mL sterile LB (pH 7.0) in 100 mL Erlenmeyer flask. The inoculated medium was incubated at 37°C overnight. The culture was harvested by centrifugation at 5,000 rpm for 5 min at 4°C and washed twice with sterile distilled water to remove traces of media. The washed pellet was finally suspended in 1 mL sterile distilled water. Aliquots of 100 μL of concentrated (1 × 108 cells/mL) cell suspension (s) were added separately to 10 mL of 0.01X Luria broth in 100 mL Erlenmeyer flasks. The flasks were kept at 37°C (without shaking). Growth was observed continuously up to 48 hr.
Amplification, cloning, and sequence characterization of the variable region of class 1 integron in MBL-positive isolates
All MBL-positive isolates were examined for the presence of class 1 integron using the CS-PCR method described earlier. 11 The amplicons were cloned and sequenced according to the method described previously. 12 Sequence analyses were done with the available Bioinformatics tools (Blast software www.ncbi.nlm.nih.gov/BLAST, ORF finder, ProtParam analysis, and sequence alignment by CLUSTAL W program).
Amplification, cloning, and sequencing of blaNDM genes
The complete coding DNA sequence of blaNDM was amplified using EcoRI-NDM-F (5′-GGGAATTCATGGAATTGCCCAATATTATG-3′) and PstI-NDM-R (5′-AACTGCAGTCAGCGCAGCTTGTCGGCCAT-3′) as the forward and reverse primer following the method described earlier. 13
Amplification, sequencing, and phylogenetic analysis of 16S rRNA genes of MBL-positive isolates
Amplification of 16S rRNA genes was done using 27F and 1492R universal primers following the earlier method. 14 Whole-cell DNA extraction, cloning, and sequencing were done as described previously. 12 The 16S rRNA gene sequence of each of the isolates was used as a query to search for a homologous sequence in the nucleotide sequence databases by using BlastN program. 15 The 16S rRNA gene sequences showing high similarities were retrieved from the GenBank database and were aligned with 16S rRNA gene sequences of the isolate by using CLUSTAL W in MEGA X software. MEGA X was used for the generation of phylogenetic trees. The evolutionary distances were calculated according to the Kimura 2-parameter method. All phylogenetic trees were constructed by the NJ (neighbor-joining) method. Tree topology was evaluated by the bootstrap analysis (based on 1,000 resamplings).
Growth studies of MBL-positive strains to demonstrate utilization of ampicillin as a sole source of carbon and energy
To examine the growth of MBL-positive isolates (one isolate from each genus) in mineral medium supplemented with ampicillin as a sole source of carbon and energy, the inoculum was prepared as follows: log-phase cells were inoculated in 10 mL Luria broth containing imipenem (50 μg/mL) and incubated at 30°C in a shaking incubator at 180 rpm for 18 hr. About 1.5 mL of 18 hr grown culture of KR01, KR23, KR25, and MR40 were aseptically transferred to an individual sterile microcentrifuge tube, and the tubes were centrifuged at 3,000 rpm for 5 min. After discarding the supernatant, the pellet was resuspended in 1.5 mL fresh sterile minimal media, centrifuged, and the supernatant discarded. The cell pellet was washed another time following the same procedure described above, and finally redissolved in 1.5 mL sterile minimal medium. From such 1.5 mL stocks, aliquots of 200 μL cell suspension were taken to inoculate each 100 mL Erlenmeyer flask containing 20 mL minimal media with different supplementations.
The basal minimal medium, used in this study, contained (g/L) the following: KH2PO4, 3.0; Na2HPO4, 6.0; NaCl, 5.0; and MgSO4, 0.1 (pH 7.0 ± 0.2). Ampicillin was used as the carbon source, and the concentrations of ampicillin used were 0.5, 1, 2, and 4 g/L, respectively. Four MBL-negative, but β-lactams-resistant control strains, Acinetobacter sp. MB 03 (Ac. No. FM865448), Pseudomonas sp. MB 08 (Ac. No. FM865444), Escherichia sp. MB 27 (Ac. No. FN396607), and Proteus sp. MB 34 (Ac. No. HF562223) (earlier reported as class 1 integron-bearing multiple-antibiotic-resistant bacteria), 16 were grown in minimal medium containing 0.1 or 0.15 or 0.25 g/L ampicillin. Flasks were incubated at 30°C for 192 hr in a shaking incubator at 120 rpm. All experiments were done in triplicate. Mean OD600 values were presented with standard error.
Results
Sample collection and processing
Sample collection and processing has been detailed in Supplementary Fig. S1.
Screening (phenotypes) of MBL-positive strains from imipenem-resistant colonies
The incidence and abundance of imipenem-resistant bacteria in water samples collected at three different sampling sites, upstream, midstream, and downstream of the Mahananda and Karala rivers, pre- and postmonsoon in 2015 were evaluated (Fig. 1a). A higher number of imipenem-resistant bacteria obtained from the waters of both the rivers in the months of April (premonsoon) and November (postmonsoon) corroborated with the water quality data showing comparatively high biochemical oxygen demand and chloride content in conjunction with lower dissolved oxygen (DO) in April and November months of 2015, indicative of higher sewage contamination (Supplementary Table S3; data source: http://emis.wbpcb.gov.in/waterquality/viewsampledatacitizen.do).
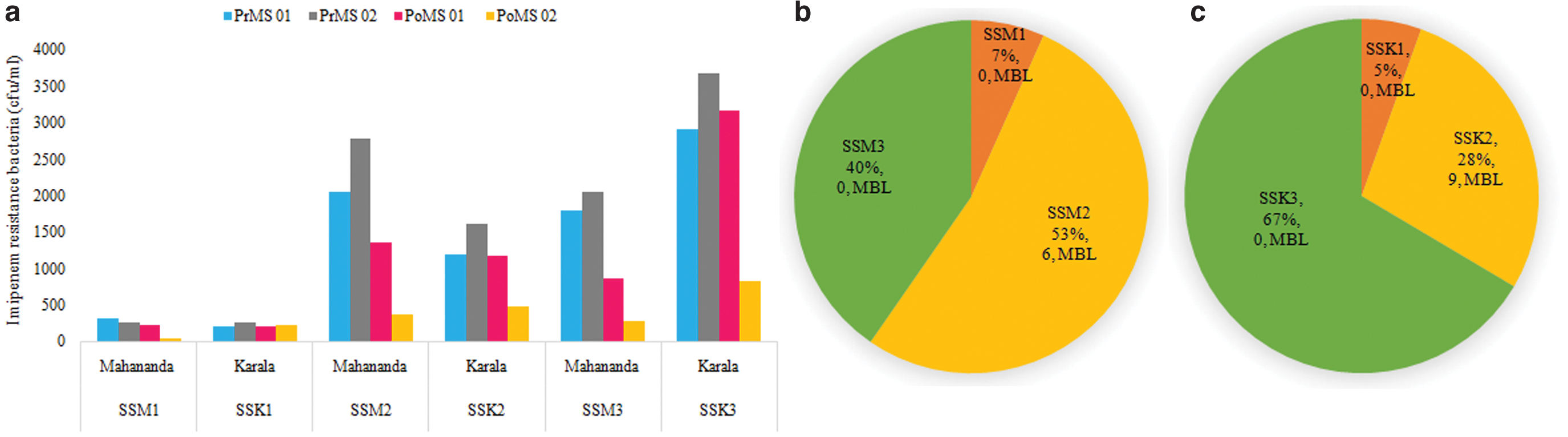
The pie charts (in Fig. 1) display the percentage distribution of imipenem-resistant strains isolated from three sampling stations SS1 (upstream), SS2 (midstream), and SS3 (downstream) of the two rivers, Mahananda (M) (Fig. 1b) and Karala (K) (Fig. 1c). It has to be noted that MBL-producing bacteria, nine and six in numbers, were isolated from the midstream (SSM2 or SSK2) samples only (Fig. 1b, c). On both the banks of the Mahananda and Karala at SSM2 or SSK2 are the highest concentrations of urban populations and hospitals. Maximum numbers of imipenem-resistant bacteria were obtained from midstream and downstream waters of the Mahananda and Karala, respectively. A total of 665 and 832 imipenem-resistant bacterial isolates were randomly selected from Mahananda and Karala river water samples for phenotypic screening of MBL-positive isolates using replica plate method. Primarily, 5.26% (35) of imipenem-resistant bacterial isolates from Mahananda river and 4.68% (39) from Karala river were unambiguously selected as MBL-positive strains as they did not show any growth on EDTA-impregnated imipenem plates.
Genotypic confirmation of phenotypically ascertained MBL-positive strains using multiplex PCR
Multiplex PCR was used to detect MBL genes within phenotypically ascertained MBL-positive isolates. Of 35 phenotypically ascertained MBL-positive bacteria isolated from Mahananda river, blaNDM gene was detected in 6 (17.14%) isolates only. About 20.51% (8 of 39 isolates) phenotypically ascertained MBL-positive isolates from Karala river had blaNDM and 2.56% (1 of 39 isolates) had blaVIM genes. Remarkably, MBL-positive strains were obtained only from imipenem-resistant bacteria isolated from midstream waters of both the rivers, and conspicuously absent in isolates of up- and downstream of both the rivers (numerically indicated in Fig. 1b, c).
Antibiotic susceptibility test (following the Kirby–Bauer disk-diffusion method) and MIC determination (following macrodilution method) for MBL-positive isolates
The antimicrobial susceptibility test by the Kirby–Bauer disk-diffusion method yielding zone diameter established for each antimicrobial (specific for Acinetobacter spp. or Pseudomonas spp. or members belonging to Enterobacteriaceae per CLSI guideline) determining resistant, intermediate, and sensitive results for MBL-positive isolates from two rivers, Mahananda and Karala, was presented in Supplementary Table S2a and b. Two Mahananda isolates, MR15 and MR20 (belonging to the genus Pseudomonas), and 2 Karala isolates, KR22 and KR23 (belonging to the genus Escherichia), were found resistant to each of the CLSI recommended genus-specific antibiotics used in this study; remaining 11 isolates, in spite of being resistant to multiple antibiotics, were also found sensitive or intermediate to one or more antibiotics (Supplementary Table S2a, b).
All the 15 MBL-positive isolates, used for determination of MIC against different groups of antibiotics, β-lactams (including cephalosporins and carbapenems), penicillins+β-lactamase inhibitors, fluoroquinolones, aminoglycoside, trimethoprim, tetracycline, colistin, and chloramphenicol, were confirmed to be multidrug resistant (MDR) per CLSI guideline to ascertain the range of resistance for a particular antibiotic (Table 2). Excluding Proteus strains, KR01 and KR04 (Proteus being intrinsically resistant to colistin), all other isolates belonging to the genera, Acinetobacter, Escherichia, and Pseudomonas, were found sensitive (<2 μg/mL) to colistin sulfate (the last resort antibiotic for the management of MDR Gram-negative bacteria) except Acinetobacter strain KR34 that has developed resistance to colistin (MIC = 64 μg/mL) (Table 2).
Resistance Profile (Differential MIC Value) of Metallo-β-Lactamase-Positive Isolates from Two City Waste Polluted River of Northern West Bengal, India
MIC, minimum inhibitory concentration; N/A, not applicable.
Exceedingly high MIC values were obtained for all MBL-positive strains against ampicillin (1,000 to >10,000 mg/L) and cefepime (512 to >1,024 mg/L; except the strain KR29). All MBL-positive isolates from Karala river, except KR29, have shown higher MIC values against imipenem (≥256 mg/L) and meropenem (≥256 mg/L) than shown by Mahananda river isolates (Table 2).
Demonstration of viability and growth of MBL-positive isolates in oligotrophic medium (diluted Luria broth, 0.1 and 0.01X)
The viability of all MBL-positive isolates in 0.1X and 0.01X Luria broth was observed. Except KR29, all the MBL-positive isolates have shown good growth in 0.1X LB. The growth response was different in 0.01X LB where five isolates (KR07, KR22, KR23, MR15, and MR20) grew well, four (KR01, KR04, MR40, and MR41) grew fairly, and the rest have shown poor growth (Supplementary Table S4).
Identification of integron borne gene cassettes in MBL-positive isolates
Nine different gene cassettes were detected in 7 (46.66%) of the 15 MBL-positive isolates (Table 3). The amplicon lengths, corresponding to the approximate sizes of the inserted gene cassette, varied from 0.7 to 3.5 kb. Sequences derived from the amplicons exhibited significant homology with already existing ARGs (Table 3). Four isolates MR40, MR41, KR22, and KR25 were found to carry two amplicons of different sizes. A single amplicon of 1.9 kb was detected in KR01, KR04, and KR07. The largest amplicon of ∼3.5 kb was recorded in MR40. The most common carriage by integron-positive isolates involved aminoglycoside adenyl transferase(s) (aadA1, aadA2, aadA5, aadB). The combination, aadA2-aadA1, was detected in KR04 and KR07. Aminoglycoside acetyltransferase [aac(6′)-Ib] was detected in MR40, KR22, and KR25.
MBL-Positive Isolates with MBL Genes and Characterized Class 1 Integron Gene Cassettes
MBL, metallo-β-lactamase; NDM, New Delhi metallo-β-lactamase; STR, streptomycin; TMP, trimethoprim.
Dihydrofolate reductase cassettes were next in abundance, and found in MR40 (dfrA5), MR41(dfrA15), and KR22 (dfrA17) (Table 3). The gene cassette(s) aac(6′)-Ib and aadA2 of Proteus sp. KR01 showed 99.81% and 99.62% similarity with the respective genes from Morganella morganii class 1 integron (MF370653); 1,197 bp amplicon of MR41 shared 100% similarity with the blaPME-1 gene of Proteus mirabilis class 1 integron (MF576130), while 730 bp amplicon exhibited 100% similarity with the dfrA15 gene of Acinetobacter baumannii class 1 integron (HQ880256).
Identification of blaNDM genes in MBL-positive isolates
blaNDM-1 was detected in six MBL-positive isolates from Mahananda river (MR15, MR20, MR26, MR29, MR40, and MR41). Five MBL-positive isolates (KR01, KR04, KR26, KR29, and KR34) from Karala river were found to carry blaNDM-1, while blaNDM-7 was found in KR07, KR22, and KR23.
Phylogenetic analysis of MBL-positive isolates by 16S rRNA gene sequences
Phylogenetic analysis based on 16S rRNA gene sequence indicated that the MBL-positive isolates under study fell in four bacterial genera, namely Acinetobacter, Escherichia, Proteus, and Pseudomonas (Supplementary Fig. S2). Seven MBL-positive isolates (MR26, MR29, MR40, MR41, KR26, KR29, and KR34) belonged to the genus Acinetobacter. In the phylogenetic tree, isolates MR26 and MR29 formed a completely separate cluster with KR26 supported by 100% bootstrap value; nearest to them were Acinetobacter equii and Acinetobacter pragensis (Supplementary Fig. S2a). KR29 and KR34 branched in a cluster with Acinetobacter Seohaensis, while MR40 and MR41 clustered with Acinetobacter variabilis (Supplementary Fig. S2a).
Three (20%) MBL-positive isolates, KR07, KR22, and KR23, were identified as the members of the genus Escherichia. 16S rRNA phylogeny showed that they were branching with Escherichia albertii with 95% bootstrap confidence value (Supplementary Fig. S2b). Only 13.33% (2 of 16) of the MBL-positive isolates belonged to the genus Proteus. In the phylogenetic tree, KR04 clustered with Proteus vulgaris, but the nearest members of KR01 were P. mirabilis and Proteus columbae (Supplementary Fig. S2c).
The genus Pseudomonas was represented by 20% (3 of 15) MBL-positive isolates (MR15, MR20, and KR25). Highest similarities at nucleotide level were recorded with Pseudomonas mendocina NCTC10897T (99.0% similarity with KR25), Pseudomonas taiwanensis DSM 21245T (99.6% similarity with MR15), and P. plecoglossicida DSM 15088T (99.4% similarity with MR20). In the phylogenetic tree, isolate-KR25 branched in a cluster with P. mendocina, but MR15 and MR20 clustered together separately from other members of Pseudomonas (Supplementary Fig. S2d).
Growth studies of MBL-positive strains in mineral medium supplemented with ampicillin as a sole source of carbon
Four MBL-positive strains, one from each genus, Proteus sp. (KR01), Escherichia sp. (KR23), Pseudomonas sp. (MR15), and Acinetobacter sp. (MR40), were selected to test the viability and growth in minimal media supplemented with ampicillin as a sole source of carbon. The concentrations of ampicillin used in this study were 1.0, 2.0, and 4.0 g/L to support the growth of KR01, KR23, and MR 15; and 0.5, 1.0, and 2.0 g/L ampicillin for MR40 (Fig. 2). The cell of Proteus sp. KR01 has demonstrated its viability till 72 hr followed by marginal cryptic growth (similar to growth that happens during stationary phase), but no visible growth showing persistent log phase (similar to a typical growth curve) took place (Fig. 2a).
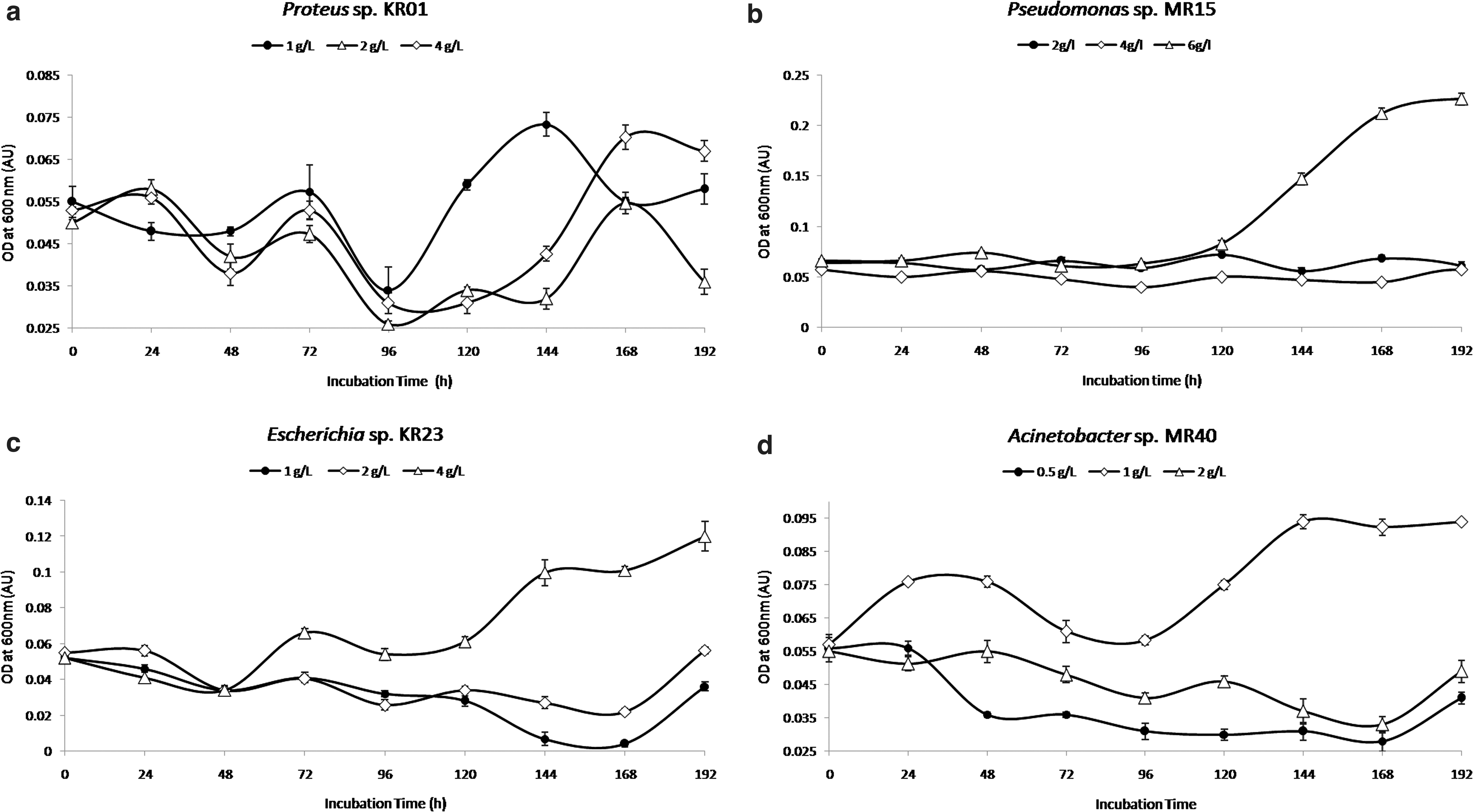
Of the different concentrations of ampicillin used as sole carbon and energy source to test growth, Pseudomonas sp. MR15, Escherichia sp. KR23, and Acinetobacter sp. MR40 demonstrated maximum growth in 6.0, 4.0, and 1.0 g/L ampicillin, respectively (Fig. 2b–d). It was found that the ampicillin concentration >250 μg/mL in minimal medium (containing glucose as the sole carbon and energy source) significantly inhibited the growth of the MBL-negative control strains, Escherichia sp. MB 27, Proteus sp. MB 34, Pseudomonas sp. MB 08, and Acinetobacter sp. MB 03, compared with their growth in the same medium in the absence of ampicillin.
Hence, we had finally selected 0.1 or 0.15 or 0.25 g/L ampicillin as sole carbon and energy source to perform their growth studies. None of the MBL-negative control strains were able to grow in minimal medium containing 0.1 or 0.15 or 0.25 g/L ampicillin under similar growth conditions.
Discussion
Imipenem and meropenem are carbapenem antimicrobial agents used to treat nosocomial and mixed bacterial infections. Resistance to carbapenems occurs either through bacterial production of β-lactamase enzymes or by reducing permeability of porin channels in the outer membrane. DNA coding for the β-lactamase production can be transmitted horizontally from organism to organism through plasmids, or vertically transferred through mutation of an existing β-lactamase enzyme. Published reports also indicate some intrinsic resistance to imipenem in a variety of clinical Gram-negative organisms, including Pseudomonas aeruginosa, Burkholderia cepacia, Acinetobacter sp., Proteus sp., Serratia marcescens, Enterobacter sp., Klebsiella pneumoniae, and Stenotrophomonas maltophilia.17,18
We have considered the changes in imipenem breakpoint criteria (S/R),10,19 and the data of an earlier study from Bangladesh 20 (showing that only 6.5% of the total MBL-producing isolates exhibited ≤8 μg/mL MIC toward imipenem; whereas 87% exhibited MIC values ≥32 μg/mL) before selecting 20 μg/mL of imipenem (a compromise concentration between 16 and 32 μg/mL) in MHA plates for obtaining more number of MBL-positive isolates from the pool of imipenem-resistant bacteria isolated from the river water samples. The level of resistance is determined by the efficiency of the enzyme for hydrolyzing the drug and by the number of resistance mechanisms present in the organism.
Organisms can produce more than one hydrolyzing enzyme and may show modifications in more than one porin, producing high-level resistance to the carbapenems (MIC ≥16 μg/mL); while organisms with decreased susceptibility produced by porin changes alone often have lower MICs (2–8 μg/mL). Earlier authors have also reported about the low prevalence of carbapenem-resistant bacteria in river water; intrinsic resistance being predominant. 21
The water quality data of the Mahananda and Karala river, of Siliguri and Jalpaiguri, respectively, of northern West Bengal, India, indicated that both are moderately polluted (unpolluted rivers typically have a BOD <1.0 mg/L) (Supplementary Table S3). The BOD was measured in the range of 1.6–2.1 mg/L in river water samples (from spots equivalent to midstream denoted in this study) in the sampling months of March and April (premonsoon), November and December (postmonsoon) of the year 2015. The DO is an important parameter to monitor the biological quality of water as it supports the aquatic life forms and regulates the biological degradation of organic impurities. In the premonsoon months, DO in the month of April for both the rivers was found to be lower than March data; while in the postmonsoon months DO recorded in the month of November was lower than December (Supplementary Table S3). A slight decrease in DO at the sampling location could be attributed to the addition of sewage in the upstream.
The chloride content of the Mahananda and Karala river water in the month of April was 1.9 and 2.7 times, respectively, the content recorded in the month of March (Supplementary Table S3). Again, in the postmonsoon months, the chloride content in the month of November of both Mahananda and Karala was higher than the value obtained in the month of December. The main source of chloride in the waters is the discharge of domestic sewage; therefore, its concentration serves as an indicator of pollution by sewage. 22
The physicochemical data corroborated the microbiological data on imipenem-resistant bacteria present in the water samples collected in the months of March and April (premonsoon), November and December (postmonsoon). Data confirm the maximum load of imipenem-resistant bacteria in the mid- and downstream of both rivers (Table 1) in the months of April and November (Fig. 1a). Among water samples collected in December from all the three sampling stations of the two rivers, imipenem-resistant bacteria were comparatively more in the downstream of Karala (Fig. 1c). This may be due to the pathway of Karala as its midstream is near to the District hospital, and in postmonsoon months maximum stagnation occurs in its downstream.
Of the total number of imipenem-resistant bacteria retrieved from Mahananda river, maximum was derived from the midstream water sample, followed by the downstream and upstream (Fig. 1b). This could be because all municipal waste channels carrying community and hospital wastewater are finally discharged in the midstream. The upstream and downstream of Mahananda drain predominantly less populated rural setting, and also fast flowing (due to persistent gradient) and less stagnating in pre- and postmonsoon months compared with Karala.
In this study, most of the imipenem-resistant but MBL-negative strains were resistant to broad-spectrum (ampicillin), broad-spectrum third-generation (ceftazidime), broad-spectrum fourth-generation (cefepime), and broadest spectrum (imipenem, meropenem) β-lactam antibiotics in addition to being resistant to tetracycline, aminoglycoside (amikacin), and fluoroquinolones (levofloxacin, ciprofloxacin). The antibiotic-resistance profiles of some representative MBL-negative, but carbapenem-resistant isolates have been shown in Supplementary Table S5. A study, from Portugal, using imipenem-containing culture media, has revealed that the majority of the river water isolates were intrinsically resistant not only to carbapenems but also to other β-lactams. 21
We have attempted to reason why there has been a low prevalence of NDM-1-positive strains in the nutrient-poor (oligotrophic) river water environment. Spread and maintenance of a resistance gene are directly linked to the fitness cost associated with the gene expression. Genes, like blaNDM-1, whose presence may significantly reduce cell fitness, are rapidly losing in the absence of selection, but few NDM-positive strains survived being the fittest (the content of carbapenems in river water being extremely reduced coupled with the oligotrophic condition compared with an environment where usages are high like in hospital setup or in copiotrophic condition prevailing in patients). Those that come at a low cost (like those contributing to intrinsic resistance) can stably persist in the cells, even when pathogens are not exposed to the antibiotic. 23 Therefore, knowing the fitness cost of a resistance mechanism is important for predicting the efficiency of its maintenance and rate of expansion. In one of the previous investigations, a decrease in fitness of blaNDM-1-bearing strains was observed for both Escherichia coli J53 and K. pneumoniae PRZ. 24
The β-lactamase genes encoding MBLs are frequently implanted in complex integrons, which are connected with transposons or plasmids having multiple drug-resistance genes that can be transferred between bacteria.16,25 One among the three MDR MBL-positive Pseudomonas spp., KR25, was found to carry blaVIM-2 within a gene cassette (Table 3). In an earlier study from China, a considerable prevalence of class 1 integrons in MDR clinical P. aeruginosa isolates was observed. 26 In the recent past, aquatic sediments receiving partially (un)treated effluents from communities of three different countries, Democratic Republic of Congo, India, and Switzerland, of diverse climatic conditions were investigated for the occurrence of MBL genes in Pseudomonas spp.
The study revealed that blaCTX-M was most frequent in Switzerland isolates, blaVIM-1 in Congo isolates but to a lesser degree in Indian isolates, but incidence of blaNDM-1 was highest among the Indian isolates and few from Switzerland isolates. 27 Out of a total of 19 class 1 integron borne gene cassettes, the most abundant (09/19) was aadA family of genes, which encode aminoglycoside-3′-adenylyltransferases (AAD) that confer resistance to streptomycin and spectinomycin by adenylation. The second most frequent (04/19) was aac(6′´)-Ib gene, which codes for aminoglycoside 6′-N-acetyltransferase type I b. This enzyme catalyzes the transfer of an acetyl group from acetyl CoA to the 6′-amino group of aminoglycoside conferring resistance to antibiotics containing the purpuroamine ring, including amikacin and kanamycin, found in a wide variety of Gram-negative pathogens.
The abundance of aadA genes in Mahananda and Karala rivers of northern West Bengal, India, could be due to the rampant use of streptomycin in agriculture. Paddy and tea are the main crops, and among horticultural crops, citrus fruits, tomatoes, and potatoes are widely grown in this region. The formulation of streptomycin sulfate + tetracycline hydrochloride (9:1) (an antimicrobial registered in India as fungicide and bactericide per insecticide act, 1968; sold as “Antibac” trade name) is sprayed in huge quantities to control fungal and bacterial diseases in tea, paddy, citrus, potato, and tomato. 28 Very recently, the state government of West Bengal had established fair price outlet for medicines where amikacin (250/500 mg injection vial) is included as important medicine under mandatory list (www.wbhealth.gov.in), and sold or distributed from district and Sadar hospitals in large quantities to treat diseases caused by Gram-negative pathogens.
Unprecedented use of amikacin in hospitals and community may have been the reason for the widespread dissemination and enrichment of aac(6′)-Ib gene in integrons. Two isolates from Karala river, KR01 and KR04, each bearing class 1 integron having two gene cassettes, aac(6′)-Ib-aadA2 and aadA2-aadA1, respectively, were found also to bear blaNDM-1. Large number of imipenem-resistant bacteria, which were not MBL-positive ones, could be those having mechanisms such as overexpression of efflux pumps, or are able to prevent carbapenems reaching their penicillin binding proteins or loss of specific portions, or mutation in Gram-positive bacteria. 29
Fourteen blaNDM possessing MDR isolates belonged to four genera, Acinetobacter, Escherichia, Proteus, and Pseudomonas, per 16S rRNA sequence phylogeny (Supplementary Fig. S2). Fifty percent of the blaNDM-1-positive isolates were found affiliated to the genus Acinetobacter. Two putative novel species (MR26 and MR29 belonging to one species; and KR26 to another species) (94.93% and 96.94%, respectively) branched out separately from the nearest cluster of A. equii and A. pragensis in the phylogenetic tree; two isolates, KR29 and KR34, were found close to A. seohaensis; and the remaining two isolates of the genus Acinetobacter, MR40 and MR41, were close to A. variabilis (Supplementary Fig. S2a). The knowledge of the occurrence and antimicrobial susceptibility pattern of Acinetobacter spp. is significant as they have come out as a cause of intensive care units (ICUs) infection. 30
Among the six most important nosocomial MDR pathogens, Acinetobacter baumannii has been reported to cause 2–10% of all hospital-borne infections caused by Gram-negative bacteria. 31 The first report from India about NDM-1 in Acinetobacter was published in the year 2012. 32 Earlier authors have indicated a higher propensity of isolation of NDM-1 producing A. baumannii from the sewage of the hospitals, but could not recover any NDM-1 producing isolates from a river or fish pond water. 33 Another important report from the state capital of West Bengal, Kolkata, India, revealed for the first time the significance of other Acinetobacter spp., comprising A. calcoaceticus, A. lwoffii, A. nosocomialis, A. junii, A. variabilis, A. haemolyticus, and genomic species 14TU, along with the high occurrence of A. baumannii strains, which caused neonatal sepsis over an extended period of 7 years. 34
We were pondering about the possession of blaNDM-1 in diverse species of the genus Acinetobacter in the waters of the Mahananda and Karala. On examining the water quality of both the rivers (Supplementary Table S3), it cannot be concluded that bacterial communities within it enjoy a nutrient-rich environment, rather they are subjected close to oligotrophic condition. All the MDR MBL-positive isolates bearing blaNDM-1 were facultatively oligotrophic in nature (Supplementary Table S4). Facultative oligotrophs are less reactive to sudden nutrient scarcity; instead, they switch on to exploit nutrient-poor conditions. Very recently, experiments on Ramlibacter aquaticus LMG 30558T have revealed that positive selection and gene expansions may embody successful adaptive strategies in oligotrophic environments.
Conversely, aquatic systems are likely to offer first-rate chances for the amalgamation of mobile genetic elements (MGEs), and thus, for cell-to-cell and cell-to-MGE contacts. Three Karala river isolates, KR07, KR22, and KR23, exhibit closest phylogenetic relationship with E. albertii (99.53%) (Supplementary Fig. S2b). Very recently, blaTEM, blaSHV, and blaCTX-M were detected in E. albertii, an emerging member of the Enterobacteriaceae causing human and animal enteric infections. 35 Two Karala river isolates, KR01 and KR04, were found phylogenetically close to P. columbae and P. vulgaris, respectively (Supplementary Fig. S2c).
Earlier to this study, two P. mirabilis isolates were found to contain blaIMP-27 gene as an integral part of class 1 integron. 36 Another hospital isolates, P. mirabilis, was reported to bear blaVIM-1 and aadA2 cassettes in class 1 integron. 37 Again, the presence of bla genes in carbapenemase-producing river water isolates from Portugal has been reported very recently. 7 A single isolate, KR25, bearing blaVIM-2 belonged to the genus Pseudomonas (Table 3, Supplementary Fig. S2). KR25 is phylogenetically nearest to P. mendocina (Supplementary Fig. S2d). Other two Pseudomonas strains, MR15 and MR20, possessing blaNDM-1, belong to P. putida lineage, closest to P. monteilii and P. plecoglossicida (Supplementary Fig. S2d).
Among the various antibiotics, β-lactam is the drug of choice because of its selective toxicity. Manufacturing of β-lactams holds the major share of pharmaceutical industries and contributes to ∼60% of the global antibiotic market. Among different β-lactams, usage of ampicillin is still rampant in India. The 21st World Health Organization (WHO) model lists of essential medicines (2019) contain ampicillin, to be used as the first choice for treating community-acquired pneumonia (severe), complicated and severe acute malnutrition, and sepsis in neonates and children ( who.int/medicines/publications/essential medicines/en/). The antibiotic consumption rate of broad-spectrum penicillins, the most frequently used class of antibiotics in terms of defined daily doses, enhanced to 36% between 2000 and 2015 globally. 38
Although ampicillin is to a degree metabolized through hydrolysis of the β-lactam ring, it is excreted basically unmodified (73–92%) in the urine, while the rest is removed through biliary excretion or through metabolic pathways. 39 Thus, removal of β-lactam contaminants from the environment, terrestrial, and aquatic system (where rivers constitute the most essential components) in particular, should be taken up as an essential bioremediation strategy. Very recently, a pan-drug-resistant strain, Pseudomonas sp. MR 02 (isolated from Mahananda river), capable of expressing blaNDM-1, was found to consume ampicillin as sole carbon and energy source to sustain its growth in mineral medium. 40
We conjectured that possession of NDM-1 in a bacterium may (in addition to rendering carbapenem resistance) provide selective (alternative carbon source) advantage in β-lactams-enriched ecological niches. Our conjecture was based on published reports that revealed NDM-1's high ampicillin cleaving efficiency (Kcat/Km = 3.84 μM−1 S−1) and its uniqueness in securing to the inner leaflet of the outer membrane.41,42 In this study, differential consumption of ampicillin as a sole carbon source for growth at concentrations 1.0, 4.0, and 6.0 g/L was demonstrated with NDM-1 producing representative strains Acinetobacter sp. MR40, Escherichia sp. KR23, and Pseudomonas sp. MR15, respectively (Supplementary Fig. 2b–d).
The results have firmly established our earlier hypothesis that the NDM is the key enzyme to support β-lactam catabolism, provided the organism has functional penicillin amidase and enzymes for phenylacetic acid catabolic pathway (paa pathway).40,43 The search for specific genes in whole-genome sequences available in the databases for bacteria has revealed the existence of paa genes and genes encoding penicillin amidases in different species under genera Pseudomonas, Escherichia, and Acinetobacter, respectively, but not in the Proteus genome (data not shown). The failure of the Proteus strain KR01 to consume ampicillin for its growth (Fig. 2a) in spite of its functional NDM-1 may be because of the absence of penicillin amidase and paa genes.
The overall distribution of NDM-positive strains gives off an impression of being heterogeneous concerning prevalence. Two large-scale studies, SMART and INFORM, led from 2008 to 2014, involving at the very least, 40 nations, have revealed 0.28 (290/103,960) and 0.19 (72/38,266) % NDM positive, respectively, among carbapenem-resistant isolates belonging to Enterobacteriaceae.44,45 This study has demonstrated that the issue of NDM-1 was not kept only to clinical pathogenic strains confined within the hospital environment but also has expanded its residence in the environmental bacteria. In conclusion, there is still a lack of understanding of the factors that influence the likelihood of a particular gene such as blaNDM-1 being transferred to a more recent, but unknown, pathogen to become a relevant human pathogen.
Footnotes
Disclosure Statement
The authors declare that they have no conflict of interest. Human/Animal Rights: This article does not contain any studies with human or animal subjects performed by any of the authors.
Funding Information
Senior research fellowship of the Department of Biotechnology, Government of India (DBT, GOI), is provided to V.K.R. Junior research fellowship of CSIR Government of India is provided to C.B. The support for consumables was derived from both DBT and University of North Bengal grant.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.