
Abstract
Polymyxin B and colistin (polymyxin E) are increasingly used as the last line of therapy for infections caused by multidrug-resistant (MDR) gram-negative pathogens. However, nephrotoxicity is still a limiting factor for the use of polymyxin. Therefore, better tolerated and more effective polymyxin derivatives are urgently needed. In this study, we aimed to evaluate the activity of the low-alkalinity polymyxin derivative, AL-6, against MDR Acinetobacter baumannii (Ab) clinical isolates in vitro and A. baumannii ATCC 19606 in vivo. Additionally, we performed a preliminarily study of the antibacterial mechanism. AL-6 showed much higher activity (0.125–0.25 μg/mL) against MDR A. baumannii clinical isolates than polymyxin E2 (PE2, 0.5–1 μg/mL). AL-6 also showed much higher activity (0.5–256 μg/mL) against polymyxin-resistant strains than PE2 (16–1024 μg/mL). Additionally, AL-6 showed slow resistance against A. baumannii. AL-6 also increased the survival rates of mice by 10% at 48 h compared with PE2 (5 mg/kg). AL-6 could be used at a dose of up to 10 mg/kg, increasing the survival rate to 30% at 72 h after infection. A preliminary study of the antibacterial mechanism showed that AL-6 permeabilized the outer membrane and destroyed cell membrane integrity. Moreover, there was a substantial increase in zeta potential (i.e., less negative) upon AL-6 exposure for A. baumannii. Overall, AL-6 carrying only four positive charges showed high activity against A. baumannii in vitro by disrupting cell membrane integrity. Higher doses of AL-6 could increase survival rates of mice. Thus, AL-6 may have potential applications as a bactericidal agent.
Introduction
The multidrug-resistant (MDR) gram-negative pathogen, Acinetobacter baumannii, is a frequently isolated hospital-acquired pathogen that can cause infections, particularly in patients in the intensive care unit (ICU) and in long-term care facilities. Infections associated with A. baumannii include pneumonia, bloodstream infections, urinary tract infections, skin and soft tissue infections, intra-abdominal infections, and central nervous system infections.1–3 Most patients infected with A. baumannii are immunocompromised or critically ill; thus, mortality rates can reach 35% in patients with ventilator-associated pneumonia and bloodstream infections caused by A. baumannii. 2
Rice 4 studied the ESKAPE pathogens (i.e., Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, A. baumannii, Pseudomonas aeruginosa, and Enterobacter spp.) as early as 2008. These pathogens mainly cause nosocomial infections and show common mechanisms of pathogenesis, transmission, and resistance. Among the ESKAPE pathogens, MDR A. baumannii has been isolated worldwide, including at military treatment facilities 5 and from patients in the ICU.6–8
Polymyxin was first made available for clinical use in the 1950s. 9 However, because of its nephrotoxicity, polymyxin was replaced by other antibiotics in the 1970s.9–12 In the 21st century, the emergence of antimicrobial resistance has become a major issue. Thus, the old antibiotic, polymyxin, has again attracted considerable attention for potential applications in treatment of MDR gram-negative pathogens. Currently, polymyxin can be used as a last-line therapy to treat MDR A. baumannii infections. However, the dosage regimens require suboptimal polymyxin exposure, which could lead to the emergence of polymyxin resistance.13–15 Therefore, better tolerated and more effective polymyxin derivatives are urgently needed.
Polymyxins B and E (colistin) are fermentation mixtures of cyclic polypeptides. The two major components of polymyxins B and E are polymyxins B1 and B2 and polymyxins E1 and E2, respectively (Fig. 1).16–19 Analysis of the mechanisms of action of polymyxin has mostly been carried out using polymyxin B (Fig. 1). The positively charged amino groups (Dab1,3,5,8,9) of polymyxin B can bind negatively charged phosphate groups of lipid A, which is the lipopolysaccharide (LPS) component of the gram-negative bacterial outer membrane. Two conserved hydrophobic domains (the N-terminal fatty acyl chain and D-Phe 6 -L-Leu 7 segment) of polymyxin B fold and form two different faces, as elucidated by the three-dimensional, solution-state, nuclear magnetic resonance spectroscopy structure of polymyxin B in complex with LPS. Then, insertion of the hydrophobic domains into the outer membrane disrupts the integrity of the cell.20–22

Structures of AL-6 and polymyxins E1, E2, B1, and B2.
Carbapenem resistance in gram-negative bacteria is a major public health concern, and many researchers have attempted to identify a new polymyxin derivative with improved activity and decreased toxicity to treat serious infections. 23 Recently, we reported the synthesis and activities of polymyxin derivatives and discovered a new polymyxin derivative, AL-6 (also named S2), which showed lower alkalinity, higher activity (minimum inhibitory concentration [MIC]: 0.25 μg/mL) against A. baumannii ATCC 19606, and lower renal cytotoxicity (50% cytotoxic concentration [CC50]: >500 μg/mL) and acute toxicity (lethal dose 50 [LD50]: 17.0 mg/kg) than polymyxin E (MIC: 1 μg/mL; CC50: 154 ± 13 μg/mL; LD50: 8.5 mg/kg). 19
Accordingly, in this study, we further evaluated the activity of AL-6 against MDR A. baumannii clinical isolates and polymyxin-resistant strains in vitro. Time–kill assays and resistance studies of AL-6 and PE2 were performed using A. baumannii ATCC 19606. The survival rates of mice infected with A. baumannii were determined after treatment with AL-6 or PE2. Finally, we preliminarily studied the antibacterial action mechanisms of AL-6 and PE2.
Materials and Methods
Materials
The A. baumannii ATCC 19606 strain was obtained from American Type Culture Collection (ATCC, Manassas). MDR A. baumannii strains (Table 1) were obtained from Shanghai Huashan Hospital (Shanghai, China). The polymyxin-resistant strain, Ab39, was obtained from our transposon mutant library of ATCC 19606, and other polymyxin-resistant, site-directed mutation strains based on strain ATCC 19606 were previously constructed in our laboratory. 24 AL-6 was synthesized and purified in our laboratory. 19 PE2 was purified from commercial polymyxin E (Zhejiang Apeloa Kangyu Pharmaceutical Co., Ltd., Zhejiang, China). The purity levels of AL-6 and PE2 were both greater than 95%, as determined by high-performance liquid chromatography. Meropenem (MEM), gentamicin (GEN), ampicillin (AMP), and levofloxacin (LEV) were purchased from Shanghai Yuanye Bio-Technology Co., Ltd. (Shanghai, China). 1-N-phenyl naphthylamine (NPN) and 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) were purchased from Sinopharm Chemical Reagent Co., Ltd. (Beijing, China). Cyclophosphamide was purchased from Huamaike Bio (Beijing, China).
Antibacterial Activity of AL-6 and PE2 Against Multidrug-Resistant A. baumannii Clinical Isolates and Polymyxin-Resistant Strains
MDR A. baumannii clinical isolates.
Polymyxin-resistant strains.
Ab, Acinetobacter baumannii; AMP, ampicillin; GEN, gentamicin; LEV, levofloxacin; MEM, meropenem; MIC, minimum inhibitory concentration.
In vitro antibacterial activity
MICs were determined using the broth microdilution method according to Clinical and Laboratory Standards Institute (CLSI) guidelines. 25 MICs were measured by diluting overnight cultures to ∼5 × 105 colony-forming units (CFU)/mL and then adding twofold dilutions of each antibiotic in 96-well plates and incubating at 37°C for 16–20 h. The test medium was cation-adjusted Mueller–Hinton broth (MHB). The MICs of each antibiotic were then recorded as the lowest concentration of antibiotic with no visible growth. At least two independent experiments were performed for each compound.
Time–kill assays
Overnight cultures of A. baumannii ATCC 19606 were diluted 1:1000 into 2 mL of fresh MHB and incubated at 37°C with aeration. Bacteria were then challenged with AL-6 or PE2 at 2 × , 4 × , and 8 × MICs. No drug was used as a blank control. For analysis, 100-μL aliquots were removed from cultures at 0, 2, 4, 8, 12, and 24 h, and 10-fold serially diluted suspensions were then plated on Mueller–Hinton agar plates and incubated at 37°C overnight. Colonies were counted, and CFU/mL was calculated. Three independent experiments were performed for each compound.
Resistance study
Overnight cultures of A. baumannii ATCC 19606 were grown in the presence of subinhibitory concentrations (0.25 × MIC) of AL-6 or PE2 to increase the probability of obtaining resistant mutants. The cells were treated with AL-6 or PE2 at 0.25 × MIC, 0.5 × MIC, and 1 × MIC. At 24-h intervals, the cultures were checked for growth. Cultures from the highest concentrations (Cn, n = passage number) that allowed growth (optical density at 600 nm, OD600 ≥2) were diluted 1:1000 in fresh medium containing 0.5 × Cn, 1 × Cn, and 2 × Cn of AL-6 or PE2. This serial passaging was repeated daily for 20 days. Three independent experiments were performed for each compound.
The frequency of spontaneous resistance development by single exposure to AL-6 or PE2 was estimated by single-step selection. Overnight cultures of A. baumannii ATCC 19606 were diluted 1:1000 into 2 mL of fresh Luria–Bertani (LB) broth with sublethal concentrations (0.25 × MIC) of AL-6 or PE2 and incubated at 37°C for 24 h. No drugs were used in the control group. Plates containing the 10 × MIC AL-6 or PE2 were inoculated with 2 × 106, 2 × 107, or 2 × 108 CFU. The mutation frequency was calculated as the average CFU generated on the drug-supplemented plate divided by the CFU in the inoculum. Three independent experiments were performed for each group.
In vivo efficacy using a mouse blood infection model
The experimental procedures involving animals were approved by the Institutional Animal Care and Use Committee (IACUC) of Shanghai Jiaotong University (No. A2018097). Institute of Cancer Research (ICR) mice were purchased from Shanghai Laboratory Animal Research Center (Shanghai, China). Overnight cultures of A. baumannii ATCC 19606 were diluted 1:100 in MHB and incubated for 4 h at 37°C. Bacteria were concentrated by centrifugation and resuspended in sterile saline for inoculation into mice. The cell concentration (CFU/mL) in saline was estimated by determining the optical density (OD600) of the suspension and confirmed by viable cell counting. ICR mice (weight, 20–22 g; n = 10 mice/group; five males and five females) were rendered neutropenic by injecting two doses of cyclophosphamide intraperitoneally, 4 days (150 mg/kg) and 1 day (150 mg/kg) before inoculation. Bloodstream infection was established by injecting 0.5 mL of early bacterial suspension (∼2.7 × 109 CFU/mL) intraperitoneally. At 1 h after inoculation, mice in the treatment groups were injected intravenously with one drug solution at 0.2 mL. Saline was used as a blank control. The PE2 dose was 5 mg/kg, and the AL-6 doses were 5 and 10 mg/kg. Deaths in each group were monitored daily for 3 days.
Scanning electron microscope analysis
A. baumannii ATCC 19606 was grown in MHB medium to 107 CFU/mL and collected before drug treatment and 0.5 and 2 h after treatment with 16 × MIC of AL-6 and PE2 and then prepared for scanning electron microscope (SEM) analysis. Phosphate-buffered saline (PBS) buffer was used as a blank control. The cells were fixed in 2.5% glutaraldehyde (prepared in PBS) for 12 h and washed with PBS three times after fixation. Then, cells were dehydrated in a series of ethanol solutions (30%, 40%, 50%, 60%, 70%, 80%, 90%, and 100% ethanol) sequentially for 15 min each and finally suspended in tert-butanol for 15 min. The cells were critical point-dried, coated with gold, and subjected to SEM analysis (Hitachi S4800; Hitachi, Japan). After extensive examination at a range of magnifications (4000 × to 20000 × ), the micrographs of the fields of view representing typical bacterial characteristics were selected.
NPN uptake assay
A. baumannii ATCC 19606 mid-logarithmic-phase cells were centrifuged and washed three times with 5 mM HEPES buffer (pH 7.2) and then resuspended in the same buffer (OD600 = 0.3). Fifty microliters of 40 μM NPN in the HEPES buffer and the test compounds in 50 μL of buffer were pipetted into the plates in advance at a final concentration of 10 μM for NPN, 16 × MIC for AL-6 (2 μg/mL), PE2 (8 μg/mL), and MEM (8 μg/mL). The 100 μL bacterial suspension was added immediately before the measurement of fluorescence; values were recorded within 3 min. Control wells were prepared as follows: (1) buffer alone; (2) buffer and NPN; (3) bacterial suspension and buffer; and (4) bacterial suspension, NPN, and buffer. NPN fluorescence was measured with excitation and emission wavelengths set at 350 and 429 nm, respectively, and a slit width of 5 nm (BioTek Synergy 2 spectrophotometer; BioTek). Each assay was performed at least three times. For each test compound, it was ascertained that alone or with 10 μM NPN, there was no fluorescence increase compared with mere NPN in buffer. The NPN uptake factor was calculated as a ratio of background-corrected (subtracted by the value in the absence of NPN) fluorescence values of the bacterial suspension and the buffer, respectively.
Zeta potential study
A. baumannii ATCC 19606 mid-logarithmic-phase cells were centrifuged and washed three times with Milli-Q water. The washed cells were resuspended in the test solution to prepare bacterial suspensions containing ∼1 × 109 CFU/mL, after which a 10-fold dilution was performed in the same medium immediately before zeta potential measurement. AL-6 and PE2 were added to bacterial cell cultures to achieve final concentrations of 2, 4, 8, and 16 μg/mL, and bacterial broth cultures were then incubated at 37°C for 20 min. Milli-Q water was used as a blank control. Zeta potential data were measured using a NiComp 380 ZLS (Particle Sizing Systems). The zeta potential was evaluated six times for each sample independently. Statistical analysis was performed with Student's t test. A p-value of <0.05 was considered significant.
Results
Minimum inhibitory concentrations
The antimicrobial activities of AL-6 and PE2 were evaluated against the A. baumannii ATCC 19606 strain and MDR A. baumannii clinical isolates. The MICs of AL-6 and PE2 are presented in Table 1. AL-6 (0.125–0.25 μg/mL) showed significantly increased antibacterial activities against both the ATCC 19606 strain and MDR A. baumannii clinical isolates compared with PE2 (0.5–1 μg/mL). AL-6 also showed much higher activity (0.5–256 μg/mL) against polymyxin-resistant strains than PE2 (16–1024 μg/mL). Thus, AL-6 was more effective against MDR A. baumannii clinical isolates and polymyxin-resistant strains in vitro than PE2.
Time–kill assays
Next, the bactericidal activities of AL-6 and PE2 were compared using time–kill assays (Fig. 2). A. baumannii ATCC 19606 was grown to early exponential phase and challenged with AL-6 (0.25, 0.5, and 1 μg/mL) and PE2 (1, 2, and 4 μg/mL) at 2 × , 4 × , and 8 × MICs, respectively. The bacterial concentration gradually dropped by two orders of magnitude in the presence of PE2 (1 μg/mL) at 2 × MIC within 4 h. In comparison, the bacterial concentration rapidly dropped by four orders of magnitude in the presence of AL-6 (0.25 μg/mL) at 2 × MIC within 4 h. The killing rates of AL-6 were faster than PE2 at 2 × , 4 × , and 8 × MICs, respectively. Accordingly, these time–kill assays showed that the lower concentrations of AL-6 (0.25, 0.5, and 1 μg/mL) display better bactericidal activity compared with PE2 (1, 2, and 4 μg/mL), respectively.
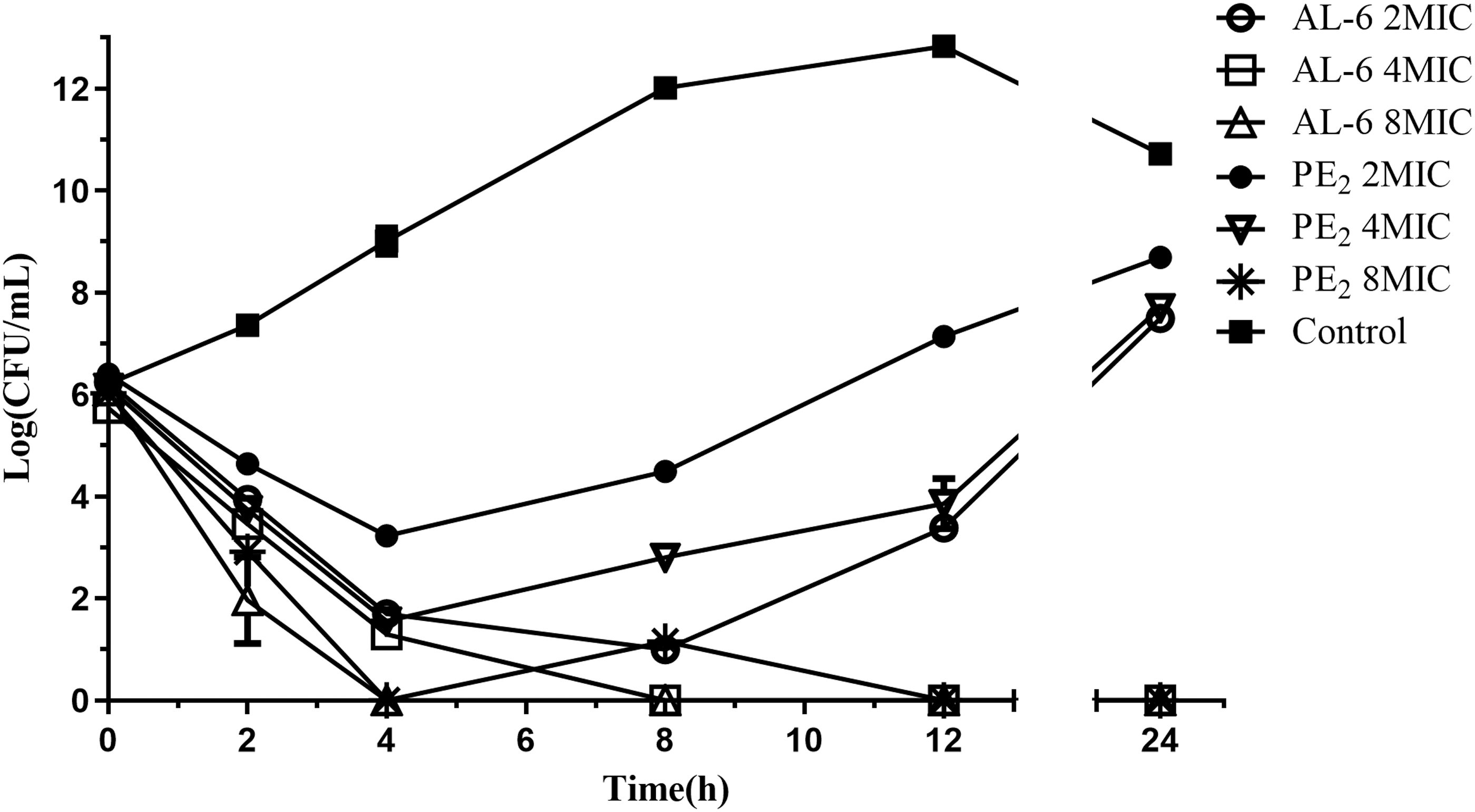
Time–kill curves for AL-6 and PE2 against A. baumannii ATCC 19606.
Resistance study
Serial passages of A. baumannii in the presence of subinhibitory concentrations of AL-6 and PE2 over a period of 20 days increased the MIC by 256 and 512-fold, respectively, that is, rendering the strain highly resistant to AL-6 and PE2. A. baumannii readily acquired resistance to PE2, with a 512-fold enhancement of the MIC in as few as 12 days (Fig. 3). Bacteria did develop resistance toward AL-6 as well, but to a greatly reduced degree, that is, 32-fold enhancement of the MIC within 12 days (Fig. 3). In comparison, A. baumannii slowly acquired resistance to AL-6 with a 256-fold enhancement of the MIC in 20 days. Therefore, AL-6 might slow the acquisition of resistance. Besides, the frequency of spontaneous resistance development by single exposure to AL-6 was lower than PE2 (Fig. 4).
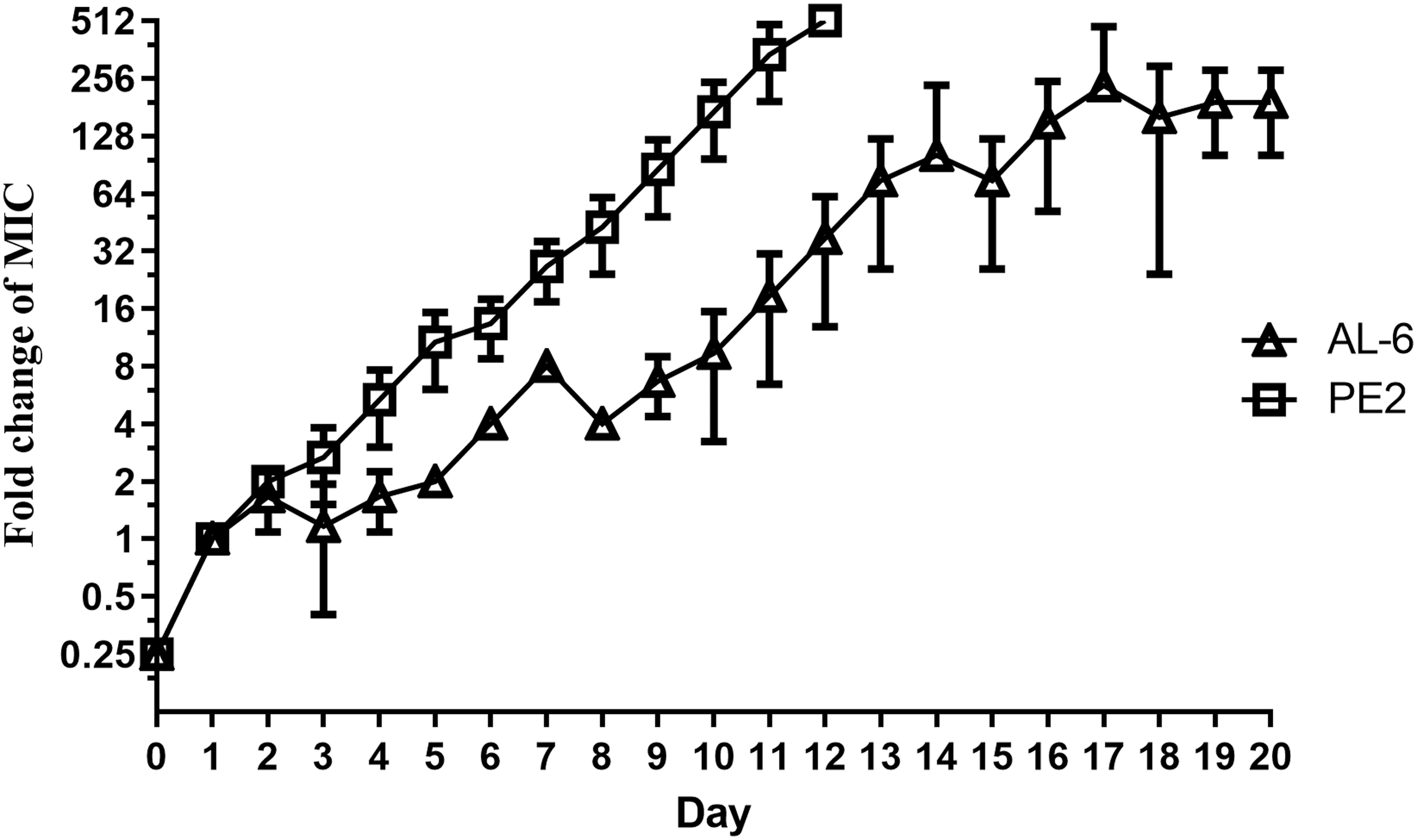
Acquisition of resistance by A. baumannii during serial passaging in subinhibitory concentrations of AL-6 and PE2.

The frequency of spontaneous resistance development by single exposure to AL-6 and PE2.
In vivo efficacy using a mouse blood infection model
The in vivo efficacies of AL-6 and PE2 were examined against A. baumannii ATCC 19606 in a neutropenic, mouse blood infection model. AL-6 (LD50: 17.0 mg/kg) was well tolerated at 10 mg/kg and PE2 (LD50: 11.3 mg/kg) was well tolerated at 5 mg/kg. 19 All the infected immunosuppressed mice died within 36 h when left untreated (Fig. 5). Similarly, all mice treated with 5 mg/kg PE2 also died within 48 h. The same dose (5.0 mg/kg) of AL-6 increased the survival rate of mice by 10% within 48 h compared with the survival rate of mice treated with PE2. Moreover, the survival rate of mice reached 30% within 72 h when treated with 10.0 mg/kg AL-6. Thus, AL-6 showed higher activity in vivo than PE2 and exhibited dose-dependent protective effects in A. baumannii-infected immunosuppressed mice.
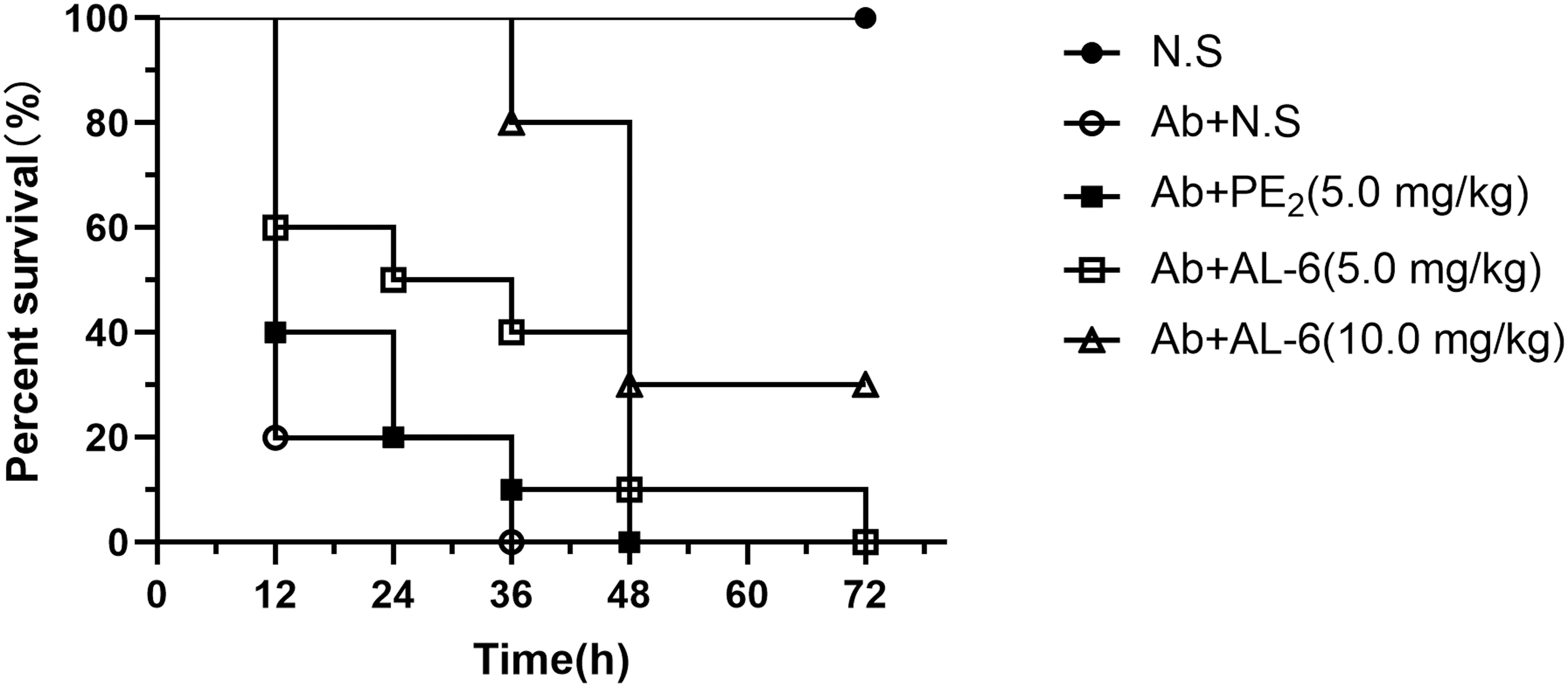
Effects of AL-6 and PE2 on the treatment of A. baumannii ATCC 19606-infected immunosuppressed mice. NS, normal saline.
Examination of the effects of AL-6 treatment on bacterial cells by SEM
Next, morphological examination of A. baumannii ATCC 19606 treated with AL-6 or PE2 was carried out using SEM (Fig. 6). AL-6 quickly disrupted the physical integrity of the cell membrane compared with PE2. After treatment with AL-6 for 30 min, the integrity of the cell membrane was obviously disrupted, and cell lysis was observed in some cells. After treatment for 2 h, AL-6 lysed almost all A. baumannii cells. Thus, these results suggested that the bactericidal effects of AL-6 may be related to rapid disruption of cell membrane integrity.

SEM images of A. baumannii ATCC 19606 with or without AL-6 and PE2 treatment.
Hydrophobic fluorescent probe uptake assays
NPN is an uncharged, hydrophobic fluorescent probe that is commonly used to evaluate the interactions of compounds with the outer membrane.26,27 NPN fluoresces more strongly in hydrophobic environments than in aqueous environments. Thus, we used NPN to study the bacterial outer membrane permeabilization activity of AL-6 and PE2 in A. baumannii ATCC 19606. Table 2 shows the effects of AL-6 (2 μg/mL), PE2 (8 μg/mL), and MEM (8 μg/mL) on the NPN uptake of A. baumannii ATCC 19606 in the form of the NPN uptake factor. Clearly, MEM rarely permeated the membrane in the control (NPN uptake factor: 1.35). In contrast, the NPN uptake factor in cells following treatment with AL-6 or PE2 was high (14.35 and 13.75, respectively), suggesting that AL-6 disrupted membrane integrity.
NPN Uptake by A. baumannii Induced by AL-6 and PE2
NPN, 1-N-phenyl naphthylamine; SD, standard deviation.
Zeta potential study
Zeta potential measurements were used to determine the influence of the compounds on the surface charge of A. baumannii.28,29 AL-6 and PE2 exposure resulted in a concentration-dependent increase (i.e., less negative) in zeta potential (Fig. 7). AL-6 exposure (2, 4, and 8 μg/mL) also significantly increased zeta potential compared with PE2 (t test, p < 0.05). However, the zeta potential values were similar for AL-6 exposure and PE2 exposure at high concentrations. Thus, the surface charge-neutralizing effects of AL-6 and PE2 exposure may be saturated at high concentrations. AL-6 carried four positive charges (Fig. 1, Dab1,5,8,9) at physiological pH, which was one positive charge less than PE2 (Fig. 1, Dab1,3,5,8,9). However, AL-6 showed stronger surface charge-neutralizing effects than PE2 at low concentrations. Thus, these results suggested that substitution of
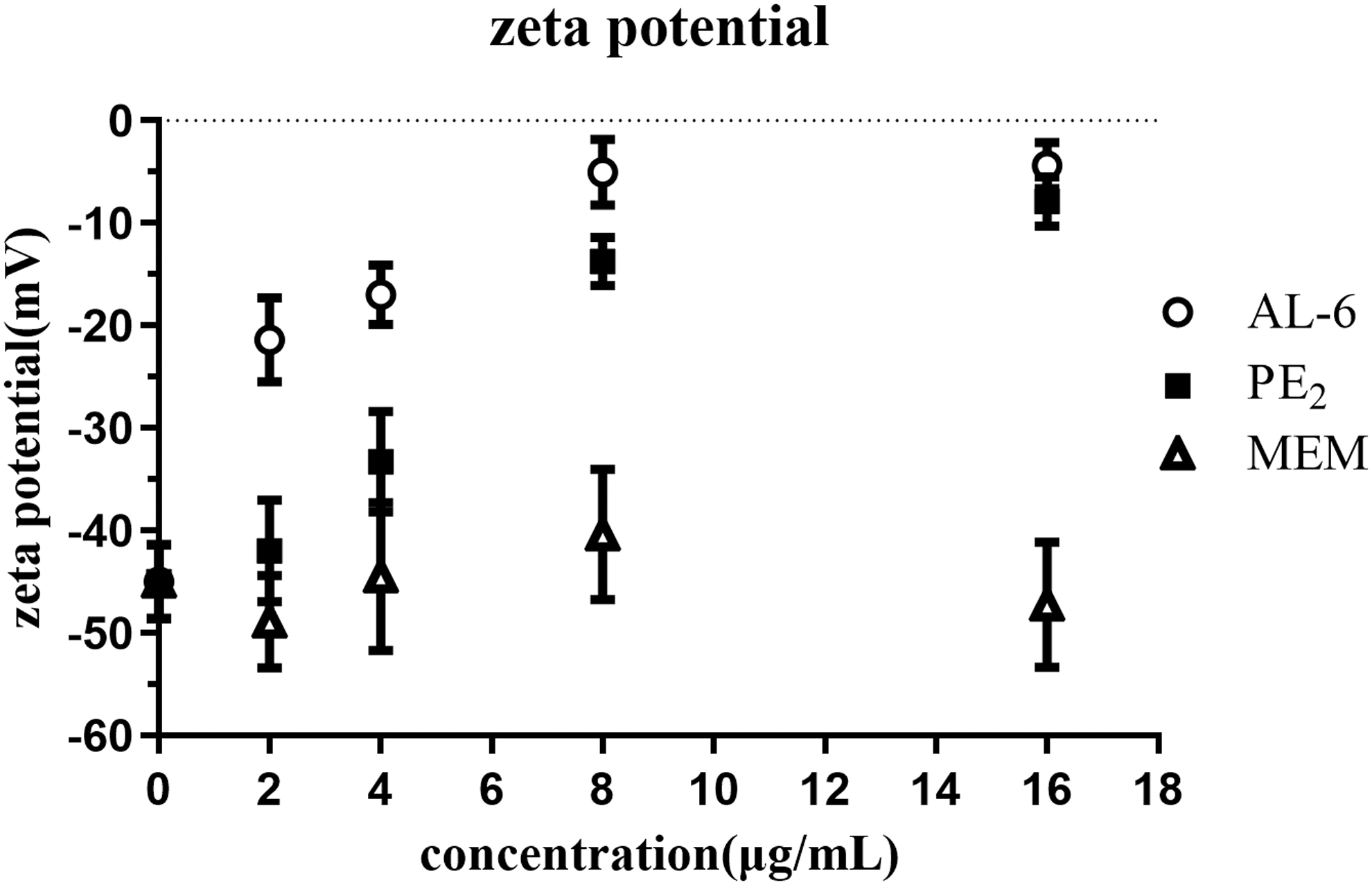
Zeta potential of A. baumannii treated with AL-6 and PE2.
Discussion
In this study, we found that the new derivative, AL-6, showed higher activity against MDR A. baumannii clinical isolates in vitro and A. baumannii ATCC 19606 in vivo than PE2. Moreover, we found that A. baumannii may not easily acquire AL-6 resistance. A preliminary analysis of the antibacterial mechanism of action demonstrated that AL-6 rapidly disrupted cell membrane integrity. Additionally, AL-6 showed stronger surface charge-neutralizing effects than PE2 at low concentrations.
The recent study showed that cationic polymyxin has a high-binding affinity for megalin and accumulates in the proximal tubule. The accumulation is then thought to drive cell death and cytotoxicity.
30
Thus, AL-6 with four positive charges has less renal toxicity.
19
Our results showed that MDR A. baumannii was more susceptible to AL-6 than PE2. Time–kill assays suggested that AL-6 displays better bactericidal activity compared with PE2. Kanazawa et al. reported that
Compared with PE2, it was slow to induce AL-6 resistance in A. baumannii. AL-6 may be a promising new bactericidal agent for the treatment of A. baumannii-induced infections. AL-6 has a heptapeptide ring and a tripeptide tail, similar to PE2. Therefore, we were not surprised that AL-6 disrupted the integrity of the cell membrane, as does PE2. Moreover, AL-6 rapidly disrupted cell membrane integrity. The strong surface charge-neutralizing effects of AL-6 indicated that the
In summary, we found that the low-alkalinity polymyxin derivative, AL-6, showed higher activity than PE2 against A. baumannii by quickly disrupting cell membrane integrity. Moreover, a higher dose (10 mg/kg) could be given to increase the survival rates of A. baumannii-infected mice. Accordingly, AL-6 might have potential applications as a bactericidal agent. Further antibacterial mechanism studies are needed.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (grant nos. 81773585, 81773616, 81573329, and 81872775), CAMS Innovation Fund for Medical Sciences (CIFMS; grant no. 2017-I2M-1-012), and Program of Shanghai Subject Chief Scientist (grant no. 17XD1423200).