
Abstract
In recent decades, the number of studies on the occurrence of resistant strains in wildlife animals has increased significantly, but data are still fragmentary. The aim of this study was to evaluate drug resistance of Escherichia coli strains isolated from wild carnivorous mammals, common in Poland. Selective media with antimicrobials (tetracycline, kanamycin, chloramphenicol, and cefotaxime) were used for isolation. Of 53 isolates shown to be distinct by the amplification of DNA fragments surrounding rare restriction site-fingerprinting method, 77.8% were multidrug-resistant (multidrug-resistant). All strains were resistant to ampicillin and many of them also exhibited resistance to tetracycline (76.2%), sulfamethoxazole (57.1%), streptomycin and kanamycin (49.2%), chloramphenicol (30.1%), and nalidixic acid (46%). In most cases, the phenotypic resistance profile was confirmed by detection of relevant genes mostly occurring in strains isolated from livestock animals and humans. Extended-spectrum β-lactamase-producing strains were detected in one mink and three martens. The strains were carriers of blaTEM-1, blaTEM-135, and blaCTX-M-15 genes. Our research confirmed a high carrier rate of MDR E. coli, even more than one MDR strain in a single individual; therefore, wider monitoring in this group of animals should be considered.
Introduction
E
However, at present, it seems important to determine the degree of contamination of the environment and to monitor the existing types of resistance and de novo emerging mechanisms increasing the insensitivity of bacteria to antimicrobials. Although the level and extent of bacterial resistance have been investigated since the beginning of the 2000s in various groups and species of wildlife animals,3,4,18 the current data do not fully reflect the real situation because of the large diversity of the species studied, wide geographical dispersion, and differences in the research methodologies used.
In this study, we investigated this phenomenon in several selected species of carnivorous wildlife animals. This is a particularly interesting group of animals, as they may accumulate multidrug-resistant (MDR) bacteria because of their behavior connected with higher synanthropization or great territorial coverage and the specific type of their diet. Moreover, several studies have reported a higher number of MDR bacteria in this group of animals compared with omnivores or herbivores.3,19
The aim of our study was to determine the phenotypic and genotypic profiles of antimicrobial resistance among E. coli isolated from carnivorous mammals in Poland and the genomic diversity of the strains in free-living animals.
Materials and Methods
The material for this study was obtained from wildlife animals collected from 2014 to 2018 in eastern Poland, mainly from the Lubelska Upland area (51°15′2.012′′ N; 22°34′12.368′ E). We analyzed 53 rectal swabs sampled from the following carnivorous species: the badger Meles meles (n = 13), the raccoon dog Nyctereutes procyonoides (n = 10), the American mink Neovison vison (n = 10), the beech marten Martes foina (n = 18), and the European polecat Mustela putorius (n = 2).
All samples were taken from wild mammals transported to the State Veterinary Laboratory in Lublin owing to suspected rabies, in accordance with regulations on combating infectious diseases in animals. 20 Because Poland is still not free from this disease, all suspected animals must be delivered and tested in official laboratories. Therefore, the sampling procedure followed the legal provisions described earlier, 21 and no permission from the ethics committee was required.
The samples were stored at −80°C for maximum 30 days until the negative results of virological test were obtained. 21
Isolation of Escherichia coli strains
Rectal swabs were initially incubated at 37°C for 24 hr in liquid medium (buffered peptone water; Biomaxima, Lublin, Poland). Subsequently, 100 μL of the suspension were plated on four types of selective MacConkey agar media (Biomaxima) supplemented with the following antimicrobials: cefotaxime (2 mg/L), tetracycline (8 mg/L), kanamycin (32 mg/L), and chloramphenicol (16 mg/L) (Sigma-Aldrich, Hamburg, Germany). The concentration of each antimicrobial was determined at the level qualifying the strain as intermediate according to the Clinical and Laboratory Standards Institute criteria. 22 After incubation (24 hr, 35 ± 2°C), one typical lactose-positive colony was randomly selected and subjected to phenotypic species identification according to the scheme described earlier. 12
Determination of phenotypic resistance profiles
We conducted preliminary selection of strains with the disk diffusion method (DDM) to eliminate the same isolates obtained from the same individual but growing on plates with different antimicrobials (MacConkey with one of the following compounds: kanamycin, tetracycline, chloramphenicol, or cefotaxime). Susceptibility to the following six antimicrobial agents was tested: gentamicin (10 μg), kanamycin (30 μg), tetracycline (30 μg), enrofloxacin (5 μg), florfenicol (30 μg), and colistin (10 μg) (Oxoid, Basingstoke, United Kingdom). The different values of growth inhibition zones classifying the isolates as resistant, intermediate, or susceptible 22 were adopted as a selection criterion in the case of differences in the susceptibility profile to at least two or more antimicrobials between isolates obtained from the same individual. This means that isolates obtained from different media but from the same individual were considered initially as distinct if their susceptibility profiles differed for at least one antimicrobial.
The minimum inhibitory concentration (MIC) was determined for the following panel of 11 antimicrobials: ampicillin, cefotaxime, ciprofloxacin, tetracycline, streptomycin, gentamicin, kanamycin, nalidixic acid, chloramphenicol, sulfamethoxazole, and nitrofurantoin (Sigma-Aldrich). The use of the interpretation criteria and both methods complied with the standards of the Clinical and Laboratory Standards Institute. 22 The extended-spectrum β-lactamase (ESBL) phenotype was determined by a double-disk diffusion test using cefotaxime, cefotaxime/clavulanic acid, ceftazidime, and ceftazidime/clavulanic acid. 23 E. coli ATCC25922 was used as a quality control. Multidrug resistance was defined as resistance to at least one agent in three or more antimicrobial classes. 24
Molecular analyses
DNA was isolated from 24-h bacterial cultures on nutrient agar (BioMaxima, Lublin, Poland) using readymade kits (Tissue and Bacterial DNA Purification Kit; Eurx, Gdańsk, Poland) according to the manufacturer's procedure. The DNA was used for the amplification of DNA fragments surrounding rare restriction sites (ADSRRS)-fingerprinting analysis and detection of resistance genes.
ADSRRS fingerprinting
The procedure was performed according to a protocol originally described for ESBL-producing Klebsiella pneumoniae 23 with some modification described by Nowakiewicz et al. 12 The procedure consisted of the following stages: restriction enzyme digestion with XbaI and BglII, ligation with appropriate adapters (Nzytech, Lisbon, Portugal), and PCR reaction carried out in a reaction mixture consisting of 5 μL of Gold Taq MIX (Syngen Biotech, Wrocław, Poland), 50 pmol of each primer (Genomed, Warsaw, Poland), and a mix obtained after the ligation reaction (2 μL). All oligonucleotide sequences used in these reactions were described by Krawczyk et al. 25
The PCR reaction conditions for the thermal cycler (T100™ Thermal Cycler; Bio-Rad, Hercules, CA) were as described elsewhere. 12 The amplification products were separated in 30% polyacrylamide gel (Sigma-Aldrich). Electrophoretic profiles were fixed using Vision-Capt Quantum (Vilber Lourmat). Cluster analysis of the strains was carried out with the unweighted pair group method with arithmetic mean using BIO-1D++ 11.9 software (Vilber Lourmat). The Dice correlation coefficient option of the software with position tolerance and optimization of 1% was used to estimate the similarity index of the isolates. Separate genotypes were distinguished at a similarity value <90% and appropriately marked with uppercase and lowercase letters of the Polish and Greek alphabets.
Determination of resistance gene profiles
The presence of selected resistance genes was tested using the PCR method. A panel of genes encoding resistance to the following antimicrobials was analyzed: phenicols (cmlA, cat, floR), sulfonamides (sul1, sul2, sul3), aminoglycosides [aph(3′)-Ia and aph(3′)-IIa], strA], aac(3)-II, aac(3)-III], and tetracycline (tetA, tetB). For molecular confirmation of the ESBL phenotype, multiplex PCR was used as in Fang et al. 26 All reactions were carried out in a thermal cycler (T100 Thermal Cycler; Bio-Rad) using PCR Mix Plus (A&A Biotechnology, Gdynia, Poland). The reaction conditions and sequences of relevant primers (Genomed) used in this study are described in Supplementary Table S1.
Statistical analysis
The frequency of resistance to the antimicrobials of the isolates associated with the host species was statistically analyzed with the R program (v 3.5.3). Data on samples containing two strains were not tested (from the European polecat), as they did not meet the criteria of the statistical tests. The Student's t-test was used for the statistical analysis of obtained values, and p < 0.05 was considered statistically significant.
Results
Among the 53 wildlife carnivorous mammals tested, 71.6% were positive for at least one resistant isolate (Table 1). No isolates were obtained from three badgers, one raccoon dog, three American minks, and eight beech martens. From 17 animals (23.9%), only single isolates were obtained (from a plate supplemented with tetracycline or kanamycin or chloramphenicol). The other isolates were obtained from samples showing bacterial growth on at least two MacConkey agar plates with various combinations of the antimicrobials: 13 samples were positive for double selective MacConkey media, whereas 5 and 3 samples were positive on triple and quadruple plates, respectively. In total, 70 bacterial strains were isolated and confirmed as E. coli (Table 1). The DDM analysis eliminated only 6 isolates (1 strain from each animal marked with an asterisk in Table 1) and 64 isolates differing in resistance to at least one antimicrobial were selected for further analyses (Table 1).
Distribution of All Cultured Escherichia coli Isolates (n = 70)
Growth on plate with: KAN—resistant to kanamycin, TET—resistant to tetracycline, CHL—resistant to chloramphenicol, CEF—resistant to cefotaxime.
Symbol and number of positive Escherichia coli-resistant animals.
Animals in which two isolates with the same resistance profile have been demonstrated using the DDM method.
Animal in which two isolates with identical ADSRRS profile have been demonstrated using fingerprinting method.
DDM, disk diffusion method; ADSRRS, amplification of DNA fragments surrounding rare restriction sites.
Characterization of genomic profiles with the ADSRRS method
The individual profiles differed in both the number of bands (from 9 to 18) and base pairs in the amplified fragments. The ADSRRS method allowed classification of most strains as distinct at a similarity level from 75% to 15%. Only five profiles grouping two isolates each were identified (Fig. 1). In this group, only two isolates (B2/CHL and B2/TET) of those detected in the samples from the badgers were classified as the same (profile assigned as “l”). The same ADSRRS profile (similarity at a level of 100%) was also obtained for strains from two different minks (N20/TET and N13/TET) (profile assigned as Δ). Another three pairs of isolates (B100/KAN and B100/TET; N13KAN and N13CHL; J85/KAN and J86/TET) were also assigned to the same genotypes (profiles assigned as m and m′, Σ and Σ′, and λ and λ′ respectively) owing to the high degree of similarity (from 90% to 91%), with the last pair of isolates originating from two different raccoon dogs.

Similarity of amplification of DNA fragments surrounding rare restriction sites-fingerprinting profiles of Escherichia coli strains.
The other strains were characterized by a high degree of differentiation (Fig. 1) and we found 14 different genomic profiles among strains isolated from the badgers, 13 among the raccoon dogs, and 11 among the minks. In turn, the number of profiles in the beech martens and European polecats was equal to the number of isolated strains (19 and 2, respectively).
Characterization of phenotypic resistance of strains
Because selective media were used for preliminary isolation, all tested strains were resistant to at least one antimicrobial agent. All strains were resistant to ampicillin and high resistance to tetracycline and sulfamethoxazole was observed among them (76.2% and 57.1%, respectively). In the case of aminoglycosides, the level of resistance to streptomycin and kanamycin was equal (49.2%), whereas only one intermediately gentamicin-resistant strain was noted. Resistance to chloramphenicol was at the level of 30.1%, with an equally high number of strains from the minks and badgers (Table 2). The percentage of ciprofloxacin- and nitrofurantoin-resistant strains was the same, that is, 28.6%, whereas the resistance to nalidixic acid was higher and reached 46%. Of the 63 strains tested, 49 (77.8%) were resistant to at least three antimicrobials and were classified as MDR strains (Table 3).
Drug Susceptibility of Escherichia coli Strains
AMP, ampicillin; CHL, chloramphenicol; CIP, ciprofloxacin; CTX, cefotaxime; GEN, gentamicin; KAN, kanamycin; NAL, nalidixic acid; NFT, nitrofurantoin; STR, streptomycin; SUL, sulfamethoxazole; TET, tetracycline.
Characterization of Resistant Escherichia coli Strains
Profiles were created from the first letters of the names of antimicrobials to which given strains are resistant: A, ampicillin; C, ciprofloxacin; N, nalidixic acid; Nf, nitrofurantoin; S, streptomycin; G, gentamicin; T, tetracycline; Sx, sulfamethoxazole; Ch, chloramphenicol; K, kanamycin; E, extended-spectrum β-lactamase.
MIC, minimum inhibitory concentration; MDR, multidrug resistant.
Only two strains among the isolates from the badgers (n = 15) were non-MDR. Thirteen isolates showed resistance to aminoglycosides (mostly to kanamycin, n = 10). There was also a high number of sulfamethoxazole-resistant strains (60%, n = 9). Resistance to tetracycline was detected in 53.3% of the strains (n = 8). 43.8% (n = 7) were resistant to quinolones and 40% (n = 6) to phenicols.
Seven (50%) of the 14 isolates obtained from the raccoon dogs exhibited resistance to at least 3 antimicrobial agents. Most strains (64.5%, n = 9) were resistant to tetracycline and/or to at least one aminoglycoside. Only 28.6% (n = 4) of the strains were resistant to sulfamethoxazole.
All isolates obtained from the minks (n = 13) were classified as MDR with resistance to at least four antimicrobials. All strains were resistant to tetracycline, whereas 84.6% (n = 11) exhibited resistance to at least one aminoglycoside and/or ciprofloxacin and nalidixic acid. Seven (53.8%) strains isolated from the minks were also resistant to chloramphenicol. One ESBL-producing strain (N19/CEF) was shown in this group as well.
Nearly 70% (n = 15) of the isolates obtained from the beech martens were classified as MDR. Resistance to tetracycline was observed in 16 strains (84.2%). Twelve strains (63.2%) were resistant to aminoglycosides and nine (47.4%) to quinolones. Three strains were classified as the ESBL phenotype (K30/CEF, K29/CEF, K32/CEF): two of them were assigned to the group of TEM producers (K29/CEF, K32/CEF) and one represented the CTX-M-1 group (K30/CEF). The strains from the TEM group were classified as MDR strains, whereas the strain possessing the CTX-M-1 group gene was resistant to β-lactam antimicrobials only.
Only two isolates from the European polecat were analyzed with the microdilution method; one of them was classified as an MDR strain with resistance to four antimicrobials (Table 3).
The statistical analysis revealed that the strains isolated from the minks were significantly more resistant to streptomycin, ciprofloxacin, sulfamethoxazole, nalidixic acid, and tetracycline than strains isolated from the other host species (Fig. 2).
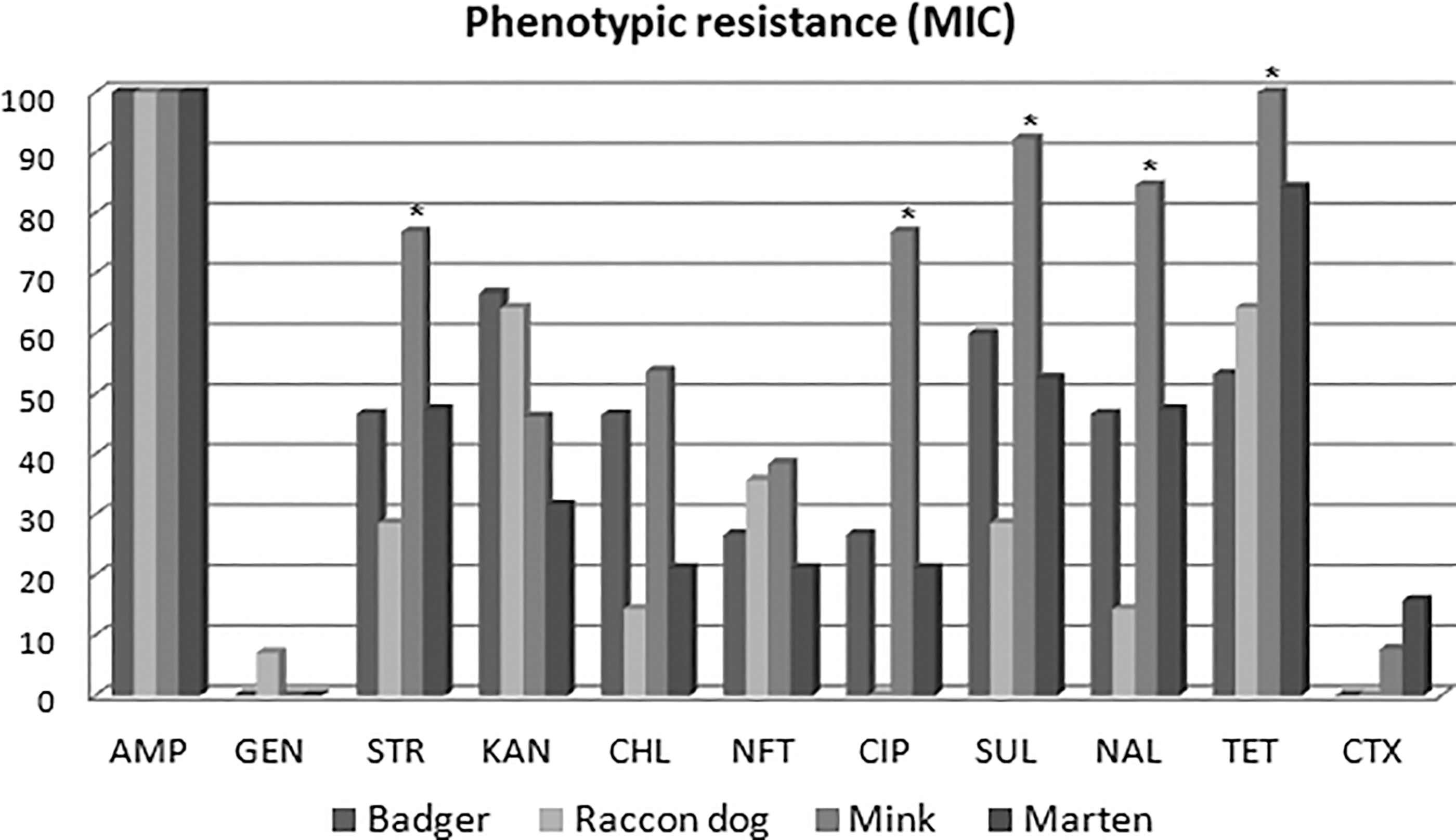
Differences in the percentage of phenotypic resistance depending on the host species. Comparison of the percentage of phenotypic resistance (evaluated with the microdilution method) among the isolates tested. AMP, ampicillin; CHL, chloramphenicol; CIP, ciprofloxacin; CTX, cefotaxime; GEN, gentamicin; KAN, kanamycin; NAL, nalidixic acid; NFT, nitrofurantoin; STR, streptomycin; SUL, sulfamethoxazole; TET, tetracycline. *Statistically significant differences. Y-axis: percentage of positive strains. X-axis: names of antimicrobials.
Relationship between the genes and phenotype of resistance
The resistance of all resistant strains to streptomycin was related to the presence of the strA gene. Among the 31 kanamycin-resistant strains, 26 had the aph(3′)-Ia gene and one strain had the aph(3′)-IIa gene. None of these genes was detected in four kanamycin-resistant isolates. Only one isolate classified as intermediately resistant to gentamicin had the aac(3)-II gene. The resistance to chloramphenicol was associated with the expression of the floR gene in eight isolates, including three strains in which this gene occurred together with the other genes tested (cmlA or cat gene). In total, seven strains had the cmlA gene, and the cat gene was only detected in six strains. The genes were not detected in two intermediately resistant strains.
The phenotypic resistance to tetracycline was confirmed by the presence of the tet gene or genes in all strains and was mainly encoded by the tetA gene in 58.3% (n = 28) or tetB in 31.3% (n = 15) of the strains, whereas six isolates had both these genes. Resistance to sulfamethoxazole was encoded by at least 1 sul gene in all strains; 16 sulfamethoxazole-resistant strains exhibited a combination of 2 genes, that is, mainly sul1 and sul2 (13 strains). The sul3 gene occurred only in three strains always in combination with another sul gene. The other 21 strains had a single sul1 or sul2 gene, that is, 19 and 2 strains, respectively.
The sequencing reactions for the blaTEM and blaCTX-M-1 genes confirmed that these genes represented blaCTX-M-15 in strains isolated from the mink and marten (N19/CEF and K30/CEF), whereas genes encoding enzymes from the TEM groups confirmed blaTEM-1 and blaTEM-135 genes (strains K29/CEF and K32/CEF, respectively).
The pairs of strains with 100% similarity of the ADSRRS profile had the same resistance gene profiles although two strains originated from two different minks (No. 13 and No. 20).
Owing to the high degree of genomic similarity, but not identity, of strains grouped in the same ADSRRS-fingerprinting profiles, we tested the whole panel of genes used in this study for the paired strains. In the case of the strain pair isolated from a badger (No. 100), we found three genes more in strain No. B100/KAN, whereas in the strain pair derived from a mink (No. 13) No. N13/KAN possessed only one more gene encoding sulfamethoxazole (Table 3). The paired strains isolated from two different raccoon dogs (J85/KAN and J86/TET) differed significantly in the presence or absence of phenotypic and genomic resistance to tetracycline and kanamycin, despite the high level of their genotypic similarity (Table 3).
Discussion
Of the 53 free-living animals of 5 different host species, 71.6% were positive for at least one resistant E. coli strains, and most strains defined as distinct were MDR (77.8%). This is a relatively high percentage of resistance compared with the results of studies obtained by other authors in Poland and worldwide.8,9,11,27 These discrepancies probably result from the substantial differences in the procedure for isolation of resistant strains, 12 the range of antibacterial substances accepted for use in a given country, and the large diversity of species in the studied groups of animals. As in the scheme followed previously, four selected antibacterial substances were used based on the resistance profiles found most commonly among isolates of commensal bacteria from livestock and human infections, 6 the level and range of antimicrobial sales in Poland, 28 and their importance in the treatment of human and animals.29,30
Despite the use of selective media with as many as 4 different antimicrobials, we obtained a negative result in 15 animals, which may indicate that resistance in free-living animals does not yet occur in the entire population; however, it is necessary to implement constant monitoring of this group of animals to assess the progression of the phenomenon, as in the case of farm animals and humans. 6 On the contrary, this may also suggest the need to expand the panel of selective substances used for isolation. The use of selective media has some limitations that should be taken into account when interpreting the results,12,31 for example, the risk of failure to detect resistance to substances that are not included in the tested drug panel in selective media or type of resistance that has so far been very rare or even completely new. On the contrary, the panel of antibacterials used can be flexibly adapted to the changing situation, for example, the recently increasing problem with acquired resistance to colistin and fluoroquinolones among commensal E. coli isolates derived from livestock animals.5,32–34
We also applied the strategy of isolation and analysis of more than one strain from the same sample, which in combination with the use of selective isolation of resistant strains facilitated demonstration of the presence of more than one multiresistant strain in the same individual. A similar isolation strategy is used relatively rarely.8,12,35 However, it yields an increased percentage of isolation of resistant and MDR strains from tested samples, but only if a reliable and repeatable method is used to eliminate clones of the same strain simultaneously. In this study, we used two-stage elimination, that is, the disk diffusion method and followed by the ADSRRS-fingerprinting method.
In the second stage, despite the use of a method with high discriminatory potential, the diversity of strains isolated from the same individual turned out to be quite large, as evidenced by the number of ADSRRS profiles comparable with the number of strains, whereas only two strain pairs achieved complete similarity. In the case of the other two pairs of strains classified into the same profiles, the similarity was at the level of 90–91%, which was reflected in the differences in both the phenotypic and genomic resistance profile in individual pairs of strains isolated from the same mink (No. 13) and the same badger (No. 100). Because of the high similarity of the ADSRRS profiles, these small variations are probably a result of horizontal gene exchange between strains inhabiting the same habitat (the same animal).
However, high similarity of genomic profiles (90%) was also observed in strains isolated from different raccoon dogs (strains J85/KAN and J86/TET). These strains not only clearly differed in their resistance profiles but also in the presence or absence of only one gene, which may also suggest that both strains probably differ only in the acquisition or loss of a mobile genome element. On the contrary, we also isolated two identical strains from two different minks, which confirms the circulation in the environment and further spread of the same bacterial clones among animals inhabiting the same niche or potentially closely related to each other. The results obtained with the use of the ADSRRS-fingerprinting method indicate a large diversity of multiresistant strains found among wildlife carnivores, and to some extent confirm the possibility of exchanging strains between individuals, mainly representing the same species. Nevertheless, we did not observe a phenomenon of grouping the strains isolated from the same host species based on a similarity coefficient, indicating that individual clones are not likely to be associated with a particular host species.
High genetic diversity also reflects rich and diverse phenotypic and gene-resistance profiles that determine multidrug resistance in most strains. This phenomenon among the isolated strains may be related to the diet of the tested animal groups. 13 Because of the wide range of food intake (small mammals, birds, fish, amphibians, and carrion), carnivorous animals can cumulate, spread, and carry resistant strains, which was also confirmed by other authors. 13 Nhung et al. 36 showed that carnivorous animals are more frequently carriers of resistance strains than omnivorous animals. Isolation of E. coli strains from birds confirmed the higher resistance of strains derived from raptors.19,37 Smith et al. 38 reported that more antibiotic-resistant strains were obtained from carnivores than from herbivorous animals.
Moreover, an additional factor increasing the level of MDR may also be the high (beech marten) or increasing degree of synanthropization of the other studied animal species. A high level of environmental contamination, especially in agricultural regions such as the Lubelska Upland, can play an important role as a resistance-increasing factor.14,39 In urbanized regions, the level of contamination with strains and/or resistance genes has been shown to be substantially higher than in areas with no sources of wastes containing anthropogenic antimicrobials.13,14 Owing to the relatively high resistance to environmental conditions, E. coli clones can survive outside the host for up to 1 year. 1
However, watercourses are an additional element facilitating spread of resistance. Studies conducted by other authors have shown that fresh waters are an important reservoir of MDR bacteria. 40 Therefore, we believe that the highest percentage of statistically resistant strains isolated from the martens may be associated with the specific habitat of this animal species (lifestyle associated with areas along watercourses).40–42
According to the World Health Organization list, E. coli strains resistant to the ESBL antibiotics are considered as a critical group of bacteria that pose a particular threat, 43 as their wide occurrence significantly limits therapeutic options. 15 The low percentage of strains with the ESBL phenotype (6.3%) shown in this study is similar to the results obtained in other groups/species of wildlife animals.8,9,27 Of importance, results of investigations conducted over the years do not indicate an increasing tendency for occurrence of ESBL-producing strains in this group of animals, regardless of the type of methodology used, for example, the use of media with cefotaxime to increase the recovery of resistant strains.8,12,27
In the case of wildlife animals, there is a large diversity of genes encoding particular groups of enzymes; however, most of them represent groups found also among isolates from farm animals and humans.27,29,30 Genes encoding beta lactamases from the TEM and CTX-M groups were detected most frequently in E. coli strains isolated from free-living animals. 44 Similar results were obtained in this study. Moreover, we have shown that both strains with blaCTX-M genes represent the blaCTX-M-15 group, which is most frequently reported in livestock, wildlife animals, and humans in European countries. 45 The same situation is observed in Poland,27,33,46 which may suggest interspecies transmission of ESBL-producing isolates. In the case of the blaTEM genes, both blaTEM-1 and blaTEM-135 genes have been detected in strains isolated from animals in Poland and worldwide.47–49 As many as three of four E. coli strains producing ESBLs were isolated from the beech marten, which may be related to the fact that this extremely synanthropic species inhabiting house buildings remains in close contact with the urban environment. Therefore, these animals are more exposed to transfer of strains with this phenotype.
Resistance to tetracycline and sulfamethoxazole is one of the most common types of resistance recorded among strains derived from wildlife animals.9,11,27 Our studies also confirmed this observation; however, the percentage of strains resistant to these antimicrobials was slightly higher compared with the results obtained by those authors. This may be associated with the use of tetracycline for selective isolation on the one hand and the presence of various genes transferred on a common mobile determinant (plasmids, transposons, integrons) on the other hand, which causes co-selection of strains resistant to other antimicrobials, that is, to sulfamethoxazole. 50
The most common configuration of sulfamethoxazole resistance genes (sul1 and sul2) in this study is also common in strains isolated from other groups of livestock and wildlife animals.7–9,12,51 The sul3 gene was first detected in E. coli isolated from pigs, 52 but it also appeared sporadically among strains isolated from wildlife. 8 The tetracycline resistance in all strains was determined by two genes dominant among tetracycline-resistant E. coli strains, either co-occurring or singly, with dominance of tetA, as in previous studies.7,9,12
We observed resistance to streptomycin and kanamycin in almost half of the tested strains (49.2%), which indicates a high level of resistance to aminoglycosides among E. coli strains isolated from wildlife animals and is comparable with results obtained for strains isolated from livestock animals6,53 and humans. 34 Resistance to high concentrations of streptomycin (MIC ≥64 μg) is common among E. coli isolates and is generally associated with the presence of the strA and/or strB gene,54,55 as in our study. Kanamycin resistance was most often determined by the aph(3′)-Ia gene, which is also responsible for neomycin resistance. This gene is widely distributed in Enterobacteriaceae within plasmids and transpozons.50,51 The kanamycin/neomycin resistance among E. coli strains isolated from animals in Poland seems to be common; a similarly high percentage was also observed among isolates from bats, 12 and the percentage observed among isolates from companion animals was equally high. 10 The high resistance to kanamycin reported in this study may also result from the use of selective media with kanamycin that were not used by other authors. 46 It may also be associated with the fact that neomycin is the most common aminoglycoside used in animals, 28 and can therefore induce cross resistance to kanamycin.
In this study, resistance to gentamicin was detected in only one strain isolated from the raccoon dog. This contrast to our previous study, 12 where >30% of strains isolated from bats in Poland were resistant to gentamicin, indicates that the level of resistance depends on the type of free-living animals studied (host species, type of diet, habitat).
The results regarding chloramphenicol, nalidixic acid, and ciprofloxacin resistance obtained in this study were comparable with previous results of investigations of bats. 12 In this study, the high (>30%) chloramphenicol resistance was determined mainly by the floR gene, which is responsible for resistance to both chloramphenicol and florfenicol. The latter antimicrobial is widely used in livestock animals, which may cause the appearance of cross-resistance and explain the high percentage of chloramphenicol-resistant strains in wildlife animals. In turn, other genes may appear not only as a result of selective pressure from phenicols, but their presence is often associated with the simultaneous occurrence with genes determining resistance to other antimicrobials, for example, aminoglycosides and sulfonamides, on the same mobile element.56,57
Conclusion
The high percentage of multiresistant isolates shown in this study confirms that wildlife animals are a significant reservoir of MDR strains. Moreover, the presence of resistance phenotypes and genes commonly found in E. coli strains isolated from livestock animals and humans suggests a strong link between wildlife and the growing anthropogenic antimicrobial contamination of the environment. Equally worrying is the fact that the animals tested in this study, likewise livestock animals, may be carriers of more than one MDR clone,58,59 suggesting that the level of resistance may be still underestimated.
This study included analysis of the level of resistance of indicator E. coli isolated from carnivorous mammals. However, our previous study involving another species of commensal bacteria Enterococcus faecalis 31 in the same group of animals also confirmed the high level of resistant bacteria, indicating that this is not an accidental result but rather a certain regularity that should still be intensively studied.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.