
Abstract
Objectives:
To investigate the transcriptional response of blaOXA-48 and the copy number alteration of IncFrepB plasmid carrying blaOXA-48 under an antibiotic concentration gradient.
Methods:
Escherichia coli strains harboring blaOXA-48 on an IncFrepB plasmid were isolated from Silchar Medical College and Hospital, Silchar, India. Sequence type and common resistance determinants were determined by PCR assay. Plasmid copy number alteration and the transcriptional expression of blaOXA-48 under different antibiotic pressures were determined by quantitative real-time PCR, and the relative fold change was measured by the ΔΔCT method.
Results and Conclusion:
The plasmid that carried blaOXA-48 in E. coli ST448 was characterized as IncFrepB and found to be conjugatively transferable. The isolates were found to coexist with blaNDM-1 within the IncX3-type plasmid. It was observed that the copy number and transcriptional response of blaOXA-48 were directly proportional to the increasing concentration of meropenem and ertapenem, whereas in the case of imipenem, it was reversed. The identification of blaOXA-48 through IncFrepB-type plasmid in this study indicates the potential route of spread of this resistance determinant in this area and also the insights we gained from the transcriptional changes of blaOXA-48 in response to different antibiotic pressures could also facilitate the development of novel or alternative therapeutic options needed for multidrug-resistant infections.
Background
After the first report from Turkey in 2001, blaOXA-48 has been increasing in a number of reports of OXA-48-producing organisms throughout the world, and the plasmid-mediated occurrence of blaOXA-48 has led to the dissemination of this resistance determinant not only in hospitals but also in different ecosystems, imposing a serious problem. 1 In the earlier studies, it has been reported that OXA-48-producing isolates are increasingly identified from different enterobacterial isolates in many parts of the world such as North Africa, the Middle East, and the European Countries, and in the majority of the cases, dissemination of this gene was found to be associated with a self-transferable IncL/M plasmid.2–4 OXA-48 and its variants are unique carbapenemases with low-level hydrolytic activity toward carbapenems and also show very weak activity against expanded-spectrum cephalosporins; however, isolates are frequently detected as multidrug resistant harboring multiple resistance determinants. 5 In the present study, we have investigated the expression of blaOXA-48 and the alteration in the copy number of the plasmid under gradient antibiotic pressure concentration. A part of this work has been presented in the joint meeting of ISAAR and ICIC 2017 (11th International Symposium on Antimicrobial Agents and Resistance and 3rd International Interscience Conference on Infection and Chemotherapy 2017) held in Busan, Korea.
Methods
Bacterial strains
Two multidrug-resistant Escherichia coli isolates harboring blaOXA-48, that is, DP-Ec101 and Dp-Ec209, were identified in a retrospective study of carbapenem-resistant E. coli isolates from Silchar Medical College and Hospital, Silchar, India, which is the only tertiary referral center in the southern part of Assam. DP-Ec101 was collected from the urine culture of a 69-year-old lady suffering from urinary tract infection and was admitted to the medicine ward, and the second strain Dp-Ec209 was isolated from a clinical specimen of endotracheal aspirate of a hospitalized female patient of 26 years admitted to the intensive care unit of the same hospital.
Coexistence of other resistance determinants
Co-occurrence of class A (blaKPC, blaIMI/NMC, and blaSME), class B (blaNDM, blaVIM, and blaIMP), and other class D (blaOXA-23 and blaOXA-58) carbapenemases was determined by the performance of different PCR assays as described earlier.6,7 In addition, the coexistence of extended-spectrum β-lactamases (ESBLs) targeting blaTEM, blaSHV, blaCTX-M, blaOXA-2, blaOXA-10, blaPER, blaVEB, and blaGES was also determined. 8 The amplified product of the resistance determinant was purified using the MinElute PCR Purification Kit (Qiagen, Germany) and then sequenced to confirm the variant of the resistant determinants.
Sequence typing
The sequence type of the isolates was determined by multilocus sequence typing, targeting the internal fragments of seven conserved housekeeping genes of E. coli. Later, the STs were assigned based on the existing E.coli database. 7 The details of the housekeeping genes and the oligonucleotides used in this study are mentioned in Table 1.
Oligonucleotides Used in This Study
Susceptibility testing
Minimum inhibitory concentrations of the isolates harboring blaOXA-48 were determined by the agar dilution method to determine the susceptibility profile of these strains against imipenem (Merck, France), meropenem (AstraZeneca, United Kingdom), ertapenem (MSD France), amikacin (Zuche pharmaceuticals, India), cefepime, aztreonam (Aristo, India), piperacillin/tazobactam (Alkem, India), and polymyxin-B (Samarth, India). Each stock solution for the corresponding antibiotic was made at 1 mg/mL concentration in nuclease-free water and stored at −80°C. The quality control for the stock solution was checked each time against E. coli ATCC 25922. The result of the susceptibility testing was interpreted as per the Clinical and Laboratory Standards Institute (CLSI) guidelines. 9
Conjugation assay
The experiment was carried out using clinical strains of E. coli coharboring blaOXA-48 and blaNDM-1 as a donor and azide-resistant E. coli J53 as the recipient. Both the donor and recipient cells were cultured in Luria Bertani broth (Hi-Media) until it reaches an optical density of 0.8–0.9 at 600 nm (OD600). Cells were mixed at a ratio of 1:5 donor-to-recipient, and the inoculated plates were incubated overnight at 37°C. Transconjugants were selected on Luria Bertani medium containing imipenem (1 mg/L) and sodium azide (1,000 mg/L) as described earlier. Plasmids were extracted from the selected transconjugants using the QIAprep Spin Miniprep Kit (Qiagen), and PCR assay was performed using the primer sequences of blaOXA-48 and blaNDM-1 (Table 1) to identify the specific resistance determinant carried within the transconjugants.
Plasmid incompatibility typing
The incompatibility type of the conjugatively transferable plasmid carrying blaOXA-48 or blaNDM-1 was determined by PCR-based replicon typing targeting 18 different replicon types, namely, FIA, FIB, FIC, HI1, HI2, I1/Iγ, L/M, N, P, W, T, A/C, K, B/O, X, Y, F, and FIIA. In addition, different groups of IncX-types such as IncX1, IncX2, IncX3, and IncX4 were also targeted. 10 The plasmid encoding the transconjugant was extracted using the QIAprep Spin Miniprep Kit (Qiagen) and was used as a template for the reaction, and the oligonucleotides that were used to identify these replicon types are mentioned in Table 1.
Plasmid stability analysis
Stability of the plasmid harboring blaOXA-48 in wild strains and transconjugants was performed by the serial passage method consecutively for 100 days (wild strains) and 40 days (transconjugants), respectively, at 1:1,000 dilutions without any antibiotic pressure. The bacterial suspension was inoculated into 10 mL LB broth and incubated at 37°C in a shaker incubator at 200 rpm for 12 hours and the steps repeated with a freshly prepared broth at 1:1,000 dilutions until the harboring carbapenemase gene got lost. After each passage, 1 mL of the culture was diluted in physiologic saline (1:1,000), and 40 μL of the diluted sample was spread on to the LB agar plate. Following overnight incubation, 50 colonies from the agar plates were randomly picked and subjected to PCR assay to confirm the presence of blaOXA-48. The loss of the resistance determinants was confirmed by performing the experiment in triplicate. 11
Plasmid copy number determination
Copy number alteration of transconjugant plasmid carrying blaOXA-48 was determined by real-time quantitative PCR using the Step One Plus Real-Time detection system (Applied Biosystem, Warrington, UK). 11 A single colony was inoculated into the LB broth containing 0.5, 1, 2, and 4 μg/mL of imipenem, meropenem, ertapenem, and oxacillin, and also without any antibiotic (considered a reference for the reaction), and incubated at 37°C for 4–7 hours until the OD reaches 0.9 at A600. The plasmid was extracted from each condition using a QIAprep Spin Miniprep Kit (Qiagen) and was used as a template. The reaction was carried out using 10 mL of SYBR® Green PCR Master Mix (Applied Biosystem), 4ng plasmid DNA as a template, and 3 mL of each primer (10picomol) in a 20 mL reaction under a reaction condition of initial denaturation at 94°C for 5 minutes followed by 40 cycles of denaturation at 94°C for 20 seconds, annealing at 52°C for 40 seconds, and extension at 72°C for 30 seconds. The relative fold change was measured by the ΔΔCT method, and the CT value of each sample was normalized against the housekeeping gene rpsLE of E. coli. Each set of reactions was run in triplicate, and the experiment was repeated thrice.
Transcriptional expression analysis of blaOXA-48
Transcriptional expression of blaOXA-48 responds to four different concentrations of imipenem, meropenem, ertapenem, and oxacillin stress (0.5, 1, 2, and 4 μg/mL) and was determined by inoculating the organisms in Luria Bertani broth (Hi-media, Mumbai, India). After 12 hours of incubation, total RNA was isolated using the Qiagen RNeasy Mini Kit (Qiagen), immediately reverse transcribed into cDNA by using the QuantiTect® Reverse Transcription Kit (Qiagen), and the cDNA was quantified by Picodrop (Pico 200; Cambridge, United Kingdom). Quantitative real-time PCR was performed using the Power Sybr Green Master Mix (Applied Biosystem) in the Step One Plus Real-Time detection system (Applied Biosystem). The housekeeping gene rpslE of E. coli was used as an internal standard. The relative expression of blaOXA-48 and the antibiotic pressure were determined by the ΔΔCt method. 12
Results
Both the E. coli strains carrying blaOXA-48 belonged to the same sequence type ST448, and these strains were also found to coharbor blaNDM-1. Susceptibility testing revealed that these isolates showed resistance toward amikacin, cefepime, piperacillin/tazobactam, imipenem, meropenem, and ertapenem, leading to a multidrug-resistant phenotype, but were found susceptible only to tigecycline and polymyxin (Table 2). Like parent strains, transconjugants carrying blaOXA-48 were also found to have a similar pattern of susceptibility profile showing resistance to all the tested antibiotics except polymyxin and tigecycline (Table 2). The blaOXA-48 resistance determinant was carried through a conjugatively transferable IncFrepB-type plasmid of ∼70 kb in size, but interestingly, blaNDM-1 was found to be located in a different 90 kb plasmid of IncX3 type. Stability analysis of the parental strains revealed that the resistant plasmids were stable until the 90th serial passage, but interestingly, the blaOXA-48 gene was found lost after the 65th serial passage, whereas blaNDM-1 carrying plasmid was found stable until the 90th serial passage. Significantly, in the case of transconjugants carrying blaOXA-48, the stability was drastically reduced, and the plasmid was maintained only for 30th serial passage, while the transconjugant carrying blaNDM-1 was stable until the 40th passage (Supplementary Fig. S1).
Minimum Inhibitory Concentrations of Clinical Isolates of Escherichia coli and Transconjugants Carrying blaOXA-48
Real-time PCR analysis revealed that the copy number and transcriptional expression of blaOXA-48 were directly proportional to the concentration of meropenem and ertapenem, that is, with the higher concentration of these antibiotics, the relative copy number and the transcriptional response of blaOXA-48 also increased, whereas in the case of imipenem, it was inversely proportional and was found to be reduced with the increasing concentration of imipenem. However, under the oxacillin pressure, the expression and copy number of OXA-48 were found to be variable with the concentration of antibiotic (Fig. 1). Transcriptional analysis revealed that the expression of blaOXA-48 was induced under the increasing concentration of meropenem, ertapenem, and oxacillin, whereas in the case of imipenem pressure, the expression level was found to be downregulated. The expressional pattern of blaOXA-48 was found specific for each concentration of the antibiotics.
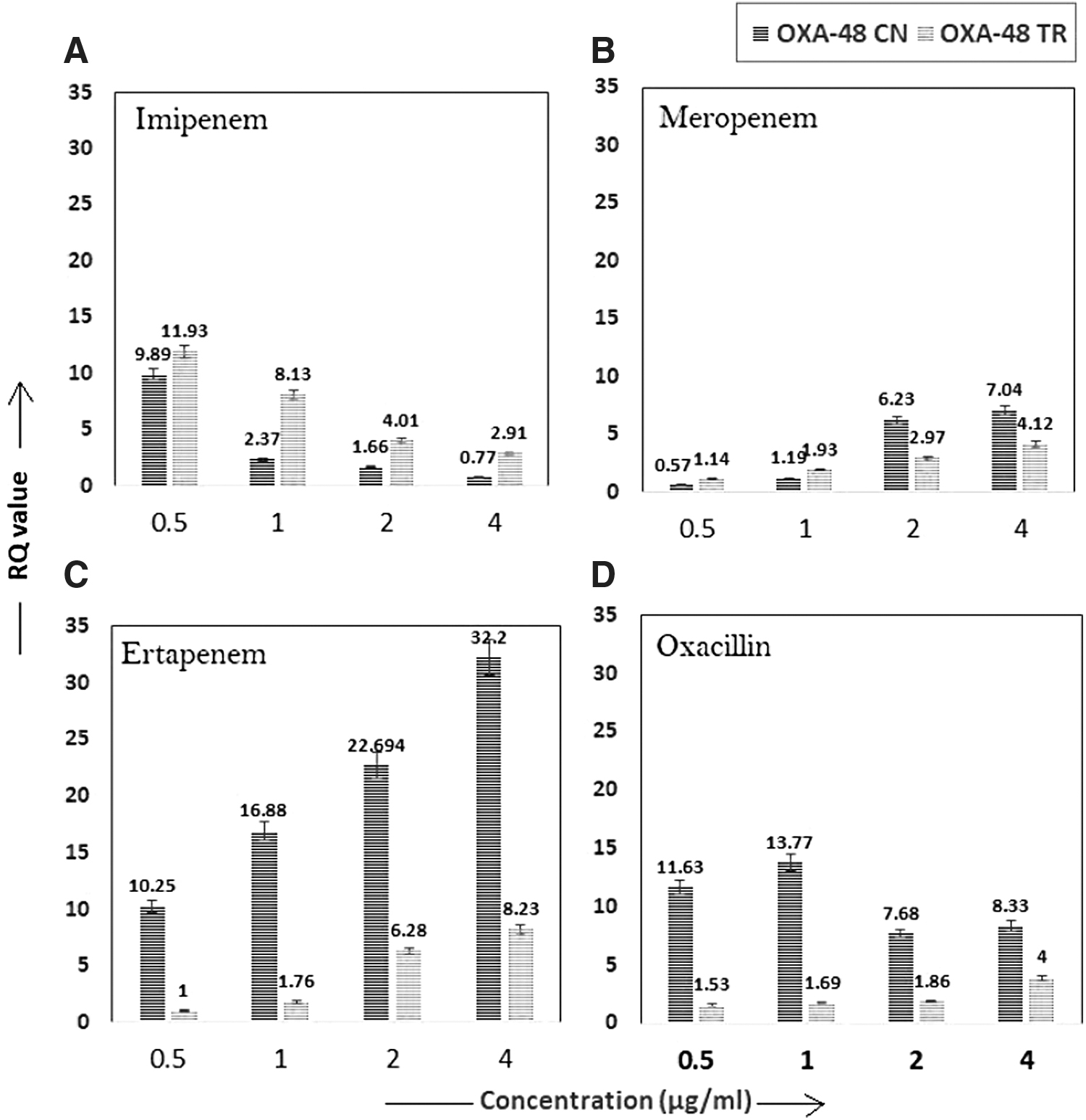
Discussion
The production of carbapenemases severely compromises treatment options, and the increased spread of these resistance determinants is considered a major public health threat. 13 Earlier, Turkey, the Middle East, and the North African countries were considered a reservoir of blaOXA-48. 14 However, the present study reporting this resistant determinant's occurrence within nosocomial isolates of E. coli from an Indian hospital underscores its wide dissemination in this subcontinent. In India, blaOXA-48 was earlier detected in Klebsiella pneumoniae and Enterobacter cloacae,15,16 and this indicates that the gene is being maintained in a diverse range of hosts. The coexistence of multiple resistance genes within an isolate is exceptionally problematic as it minimizes therapeutic alternatives, and this may depend on their transfer by mobile genetic elements and stable plasmids with and without selective pressure in clinical settings. The high-stability nature of the plasmid encoding blaOXA-48 indicates the potential ability to be maintained within the host for several generations, even after the withdrawal of antibiotic pressure. 17 In earlier studies, it was recorded that the blaOXA-48 gene was customarily linked to the IncL/M-type plasmid. In contrast, the present study reports the IncFrepB type as a carrier of blaOXA-48, which warrants further investigation to track the origin and acquisition. The antibiotic susceptibility profile of the parent strains and the transconjugants carrying blaOXA-48 was found to be in a similar pattern in this study, which is in contrast to the previous reports where the weak activity of blaOXA-48 toward carbapenems and cephalosporins was observed and indicates the growing resistance pattern over a period of time. 5
This study also shows that antibiotic pressure increases the copy number of the plasmid. In the case of ertapenem pressure, the copy number alteration was found unique compared with other carbapenems. So, maintaining this high copy number of the plasmid within the cell under some selection pressure may create an impact on the fitness and survival of the harboring organism. However, no previous study is found to support our data.
The current study shows that the spread of the blaOXA-48 gene is propagated through IncFrepB-type plasmid in India, which underscores the necessity of systematic screening of hospitalized patients and early detection of these multidrug-resistant bacteria. Also, the insights that we have gained from the transcriptional changes of blaOXA-48 in response to different antibiotic pressures can facilitate the development of novel or alternative therapeutic options that are needed for multidrug-resistant infections.
Ethical Approval
The work was approved by Institutional Ethics committee of Assam University, Silchar vide Reference Number: IEC/AUS/C/2014–001.
Availability of Data and Materials
All the relevant data and information are presented in the article.
Footnotes
Acknowledgment
The authors sincerely acknowledge the financial support provided by the Council of Scientific and Industrial Research (CSIR) to carry out the work.
Authors' Contributions
D.P.: Performed the experimental work, data collection and analysis, and prepared the article. D.D. and A.C.: Participated in experiment designing and article correction. A.B. Supervised the research work and participated in designing the study and drafting the article. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
The Council of Scientific and Industrial Research (CSIR Grant number 37(1632)/14/EMR-II), and Dr. Deepjyoti Paul was a Senior Research Fellow in the Department of Microbiology, Assam University, Silchar, and received the CSIR Senior Research Fellowship under grant number 37(1632)/14/EMR-II.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.