
Abstract
The objective of this study was to characterize Polish penicillin-resistant, ampicillin-susceptible Enterococcus faecalis (PRASEF), increasingly reported to the National Reference Centre for Susceptibility Testing, Poland, to elucidate the path of emergence of such strains. A total of 136 isolates were examined by antimicrobial susceptibility testing and for the β-lactamase production (cefinase test). The clonality of isolates was established by multilocus sequence typing (MLST) and the penicillin-binding protein pbp4 gene was sequenced to search for putative mutation(s). The presence of pheromone-responsive plasmids was investigated by clumping test and PCR detection of plasmid-specific genes. All Polish PRASEF were multidrug resistant and β-lactamase-negative. MLST assigned isolates mostly to high-risk enterococcal clonal complexes (HIRECCs) 6 (57.4%) and 87 (30.1%), in addition to to CC88 (12.5%). The sequencing of pbp4 revealed mutations upstream of a putative promoter region and amino acid alterations in PBP4, affecting 24 positions and resulting in 30 variants. While production of aggregation substance was observed for 17.6% of isolates, genes of pheromone plasmids were much more commonly detected. However, no conjugal transfer of penicillin resistance was observed. Penicillin resistance in E. faecalis emerges mostly in HiRECCs due to PBP4 overproduction and/or mutations. The acquisition of penicillin resistance by HiRECCs may represent the next step in the evolution of E. faecalis as human nosocomial pathogen.
Introduction
Enterococci occur commonly as harmless components of gastrointestinal microbiota of humans and animals, but these bacteria also cause serious infections in hospitalized patients with impaired immunity. Currently, enterococcal species are ranged the third most common cause of hospital-associated infections (HAIs), with the most significant proportion of Enterococcus faecalis.1,2 Enterococci are successful nosocomial pathogens since they are intrinsically resistant to multiple antimicrobial agents (EUCAST Expert Rules Version 3.1. Intrinsic Resistance and Exceptional Phenotypes Tables, and they acquire nonsusceptibility to other drugs, such as β-lactams, quinolones, glycopeptides, and high concentrations of aminoglycosides. 3
Resistance to penicillin and ampicillin is very common among clinical Enterococcus faecium, in contrast to E. faecalis. However, some studies showed increased frequency of penicillin-resistance among clinical isolates of E. faecalis (reviewed in Gawryszewska, et al. 4 ). This phenomenon is of a big concern, as synergistic combination of high doses of aminoglycosides with cell-wall active agents such as glycopeptides or penicillins is used in treatment of enterococcal infections. 5 So far, three mechanisms of penicillin resistance in E. faecalis have been described, including (1) production of β-lactamase, which hydrolyses the antibiotic 6 ; (2) overproduction of the cellular target of antibiotic, penicillin-binding protein PBP4 (also known as PBP5; 680 amino acids in length)7,8; and (3) changes in PBP4, leading to the decreased affinity of PBP4 to β-lactams.8–12
The acquisition of ampicillin resistance in the case of nosocomial E. faecium constituted an important step in the formation of hospital-adapted meroclone of this pathogen 13 contributing to its current rise in HAIs, and it is plausible that a similar scenario might occur as well for E. faecalis. The nosocomial subpopulation of E. faecalis consists in a big part of specific clonal complexes (CCs), so-called high-risk enterococcal clonal complexes (HiRECCs),14–16 adapted to hospital settings. With the use of multilocus sequence typing (MLST), three HiRECCs have been identified so far for E. faecalis (CC6, CC9, and CC87).15–17
Evolution of HiRECCs is driven by an acquisition of resistance determinants and virulence factors,18,19 with a big role played by mobile genetic elements, especially pheromone-responsive plasmids, which have unique mechanism of transfer requiring pheromone signals from plasmid-free recipient cells.20,21 This group of conjugative plasmids occurs exclusively in enterococci and have been mostly described in E. faecalis. Pheromone-responsive plasmids frequently carry genes encoding enterococcal virulence factors, such as aggregation substance (Asa1 and Asa373), cytolysin (Cyl), and pili (Bee proteins)22,23 and determinants of resistance to aminoglycosides, penicillins (β-lactamase gene), glycopeptides, and other drugs. 24 It was shown that they are also able to mobilize parts of chromosome during conjugation, for example, a pathogenicity island, vancomycin resistance transposon Tn1549, and a capsular operon. 25
Penicillin-resistant, ampicillin-susceptible E. faecalis (PRASEF) have been recently reported to the National Reference Centre for Susceptibility Testing (NRCST) with an increasing frequency. The objective of presented study was to characterize these isolates by establishing their clonal relatedness, molecular basis of penicillin resistance, and ability of isolates to disseminate resistance to elucidate the possible evolutionary origins of such strains and their further epidemic potential.
Materials and Methods
Bacterial strains
The study included 136 PRASEF isolates, collected by the NRCST from 32 medical centers located in Poland, from August 2006 (when the first PRASEF was received), till the end of 2016. The majority of isolates were obtained in 2015 and 2016 (41 and 48, respectively). Isolates were recovered mainly from patients with noninvasive infections such as urinary tract infections or surgical site infections (98; 72.1%), and invasive infections (29; 21.3%); eight isolates (5.9%) were obtained from carriage (rectal swab, catheter).
Antimicrobial susceptibility testing
Susceptibility of isolates to penicillin, ampicillin, imipenem, gentamicin, streptomycin, vancomycin, teicoplanin, ciprofloxacin, tetracycline, tigecycline, erythromycin, chloramphenicol, rifampicin, and linezolid was evaluated by broth microdilution method, according to the ISO-20776-1 standard, 26 and daptomycin susceptibility with the use of strips containing predefined antibiotic concentrations (bioMérieux, Marcy l'Etoile, France). The ATCC 29212 strain of E. faecalis was used as a quality control. Interpretation of observed minimal inhibitory concentrations (MICs) was conducted according to the European Committee on Antimicrobial Susceptibility Testing 27 or the CLSI clinical breakpoints, 28 when the EUCAST breakpoints were not available. The isolates were classified as multidrug resistant (MDR) when they showed nonsusceptibility to three or more classes of antimicrobials tested. 29
Clumping assay and conjugation experiments
Occurrence of pheromone-responsive plasmids was assessed in phenotypic assay evaluating ability to produce the aggregation substance (AS), manifesting itself as clumping of cells in the presence of sex pheromone 30 present in the culture supernatant of E. faecalis OG1X. The pheromone-secreting strain E. faecalis OG1X was cultured in Todd Hewitt broth (THB) at 37°C for 18 hr, centrifuged at 3,500 rpm for 20 min, supernatant sterilized by filtration, and diluted 1:5 in THB. About 200 microliter aliquots of pheromone-containing medium were inoculated with overnight cultures of tested strains (1:200) and incubated at 37°C. The clumping of cells was visually examined after 2, 4, 6, 8, and 24 hr. E. faecalis with pAM373 or pCF10 and E. faecalis OG1X were used as positive and negative controls, respectively.
Conjugational transfer of penicillin-resistance determinant was investigated following highly efficient conjugation protocol using the entire surface of agar plates 25 with the rifampicin-resistant E. faecalis OG1RF as a recipient strain. Briefly, 500 μL of overnight culture of rifampicin-susceptible donors and recipient strains in Brain Heart Infusion broth (BHI) were centrifuged for 5 min at 7,000 rpm, and washed with 1 mL of BHI. Washed cells were resuspended with 125 μL of BHI, mixed, and spread onto the BHI agar plates. After 24 hr of incubation at 37°C, cells were resuspended in 1 mL of BHI, and 100 μL were plated on BHI agar supplemented with rifampicin (32 mg/L) and penicillin (16 mg/L). Plates were screened for transconjugants after 48 hr incubation at 37°C.
DNA isolation and MLST
Bacterial DNA was isolated with the Genomic Mini kit (A&A Biotechnology, Gdańsk, Poland). Clonal relatedness of isolates was assessed by MLST 15 with the use of the MLST database. 31 Association of sequence types (STs) with the known CCs was established by the comparative goeBURST analysis (implementation of the eBURST algorithm rules proposed by Feil et al.) 32 against the whole database of E. faecalis.
Detection and analysis of penicillin resistance determinants
β-Lactamase production was investigated with the use of cefinase test (Cefinase™, Becton Dickinson, Sparks, MD) according to the CLSI recommendation. 26 The pbp4 gene was amplified and sequenced using specific primers (Ono et al. 9 and this study; Supplementary Table S1). Obtained sequences were compared to the wild-type pbp4 of penicillin-susceptible JH2–2 strain of E. faecalis (AJ290435.1) to detect possible mutation(s) within the promoter region and the pbp4 ORF using the Lasergene package (DNASTAR, Madison, WI). The deduced amino acid sequences of PBP4 were compared to the wild-type PBP4 and used as a query in blastp searches in GenBank.
Detection of vancomycin resistance determinants genes and genes specific for pheromone-responsive plasmids
The detection of the vanA and vanB genes was performed by PCR,33,34 with E. faecium BM4147 and E. faecalis V583 as positive controls, respectively. Occurrence of genes specific for pheromone-responsive plasmids was investigated by PCR, including genes associated with replication (rep8pAM373 and rep9pAD1/pTEF2/pCF10), 35 stabilization (par), 36 re1pAD1, re1pCF10, 37 and virulence (asa1, asa373, cylA, bee1, and bee23)38–40 (Supplementary Table S1). DNA of enterococcal isolates from our laboratory collection served as positive controls,41,42 except re1pCF10 for which a few randomly chosen PCR products were sequenced to confirm PCR specificity.
Nucleotide accession numbers
The nucleotide sequences of pbp4 were deposited in GenBank database under the following accession numbers: JQ307813-JQ307816, JQ307818-JQ307829, MN176166-MN176179, and MN275190.
Results
Antimicrobial susceptibility
Analyzed PRASEF isolates displayed MICs of penicillin and ampicillin in the range of 16–32 and 0.5–4 mg/L, respectively (Table 1). All isolates were resistant to ciprofloxacin and frequently displayed nonsusceptibility to erythromycin, high-level gentamicin resistance (HLGR), and high-level streptomycin resistance (HLSR). Vancomycin resistance was detected in 60 isolates, predominantly associated with the presence of vanB (57 isolates; three remaining isolates carried vanA). Investigated isolates were less frequently nonsusceptible to chloramphenicol and rifampicin, sporadically displayed resistance to tigecycline (four isolates) and linezolid (one isolate, previously analyzed in Gawryszewska, et al. 43 ), and were all susceptible to imipenem and daptomycin. All PRASEF displayed the MDR phenotype.
Antimicrobial Susceptibility of 136 PRASEF Isolates Collected in Poland from 2006 to 2016, Including the Range of MIC, MIC50, and MIC90 Values, and Distribution of Isolates Among Susceptibility Categories for Each Antimicrobial Agent Tested
CLSI breakpoints: S, susceptible; I, intermediate; R, resistant.
MIC50 and MIC90, concentration of antimicrobial at which 50% and 90% of isolates were inhibited, respectively; S, susceptible, standard dosing regimen; I, susceptible, increased exposure; R, resistant; -, susceptible, increased exposure category not defined for this compound; HLGR, high-level gentamicin resistance; HLSR, high-level streptomycin resistance; MIC, minimum inhibitory concentration; PRASEF, penicillin-resistant, ampicillin-susceptible Enterococcus faecalis.
Clonal relatedness of isolates
MLST assigned the investigated isolates to eight STs, belonging to HIRECCs 6 (exclusively ST6, 78 isolates) and 87 (5 STs, 41 isolates). The remaining 17 isolates belonged to two STs in CC88 (Table 2).
Characteristics of 136 Polish PRASEF Isolates Collected in Poland from 2006 to 2016, Including Their Epidemiological Data, β-Lactam Susceptibility Profiles, ST/CC Assignment, and Molecular Determinants of Penicillin and Vancomycin Resistance
Number of isolates provided within the brackets. PEN, penicillin; AMP, ampicillin; IMI, imipenem; wt, wild-type; clinical breakpoints used: penicillin S ≤ 8, R ≥ 16; ampicillin S ≤ 4, R > 8; imipenem S ≤ 4, R > 8. Medical centers: BYD, Bydgoszcz; CC, clonal complexe; CZE, Czestochowa; DZI, Dzialdowo; GLO, Glogow; GRO, Grodzisk Mazowiecki; GRY, Gryfice; KAT, Katowice; KRE, Krakow; KRP, Krapkowice; LUB, Lublin; NOW, Nowa Sol; OPO, Opole; PBD, penicillin-binding domain; POZ, Poznan; RAD, Radom; SLU, Slupsk; ST, sequence type; SUW, Suwalki; TRZ, Trzcianka; WAR, Warsaw; ZAB, Zabrze.
Analysis of penicillin resistance determinants
All isolates were negative for the β-lactamase production. Sequencing of pbp4 revealed nucleotide mutations within both the promoter region and the coding sequence (Table 2 and Fig. 1). All investigated isolates had nucleotide alteration(s) in the region upstream of the pbp4 ORF. A single base pair deletion of an A residue located eight bases upstream of a putative −35 region (“1” in Fig. 1A) was characteristic for all isolates of CC6 and CC87. ST832 isolates from CC87 additionally carried A/T mutation 13 bases upstream of a putative −35 region (“1” in Fig. 1A). All isolates of CC88 had C/T mutation two bases upstream of a putative −10 region (“1” in Fig. 1A).
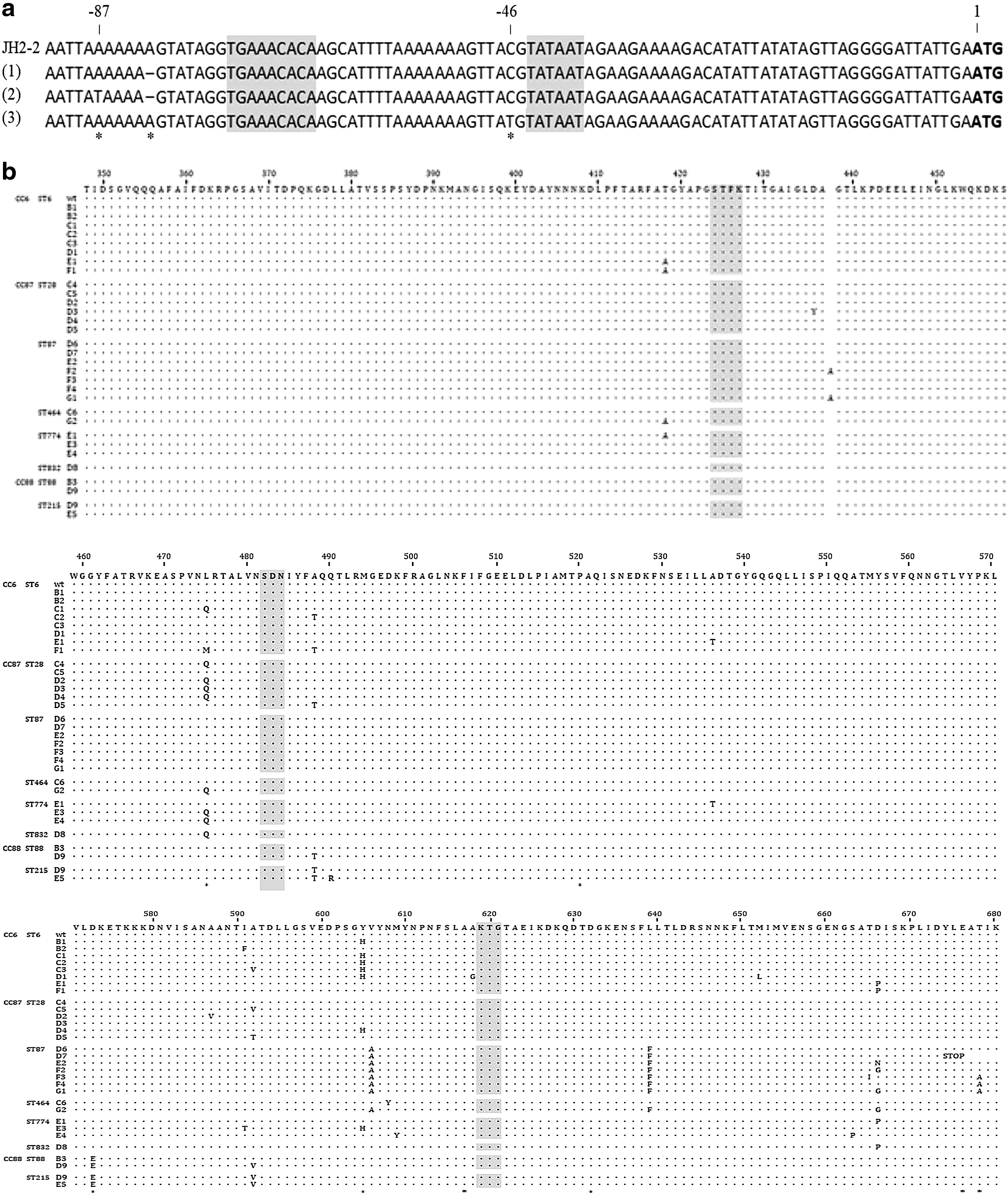
Sequence analysis of the pbp4 gene among Polish PRASEF isolates. (a) The upstream regions of pbp4. Mutations marked by an asterisk; the putative −35 and −10 regions shaded; ATG of pbp4 in bold. (b) Sequence variation of the PBP4 penicillin-binding domain. Catalytic motifs shaded; positions of amino acid substitutions previously identified as associated with β-lactam resistance marked by asterisks. PRASEF, penicillin-resistant, ampicillin-susceptible Enterococcus faecalis.
Analysis of the pbp4 coding sequence revealed amino acid changes in 24 positions, mostly located in the penicillin-binding domain (350–680 amino acids) and resulting in 30 PBP4 variants named with letters according to the number of amino acid changes in comparison to the wild-type protein from zero (A) to six (G). PBP4 variants with the same letter and different numbers represent protein variants with the same number of amino acid changes but located in different positions.
Among identified PBP4 variants, only the wild-type A protein (seven isolates) and three PBP4 variants, including B1 (Efs EnGen0231 E0I27410.1, Efs EnGen0203 EOM54637.1), B3 (Efs UFTM 20–06 AGW23922.1), 11 and E1 (Efs RefSeq WP_048943053.1) were known previously. Two novel PBP4 variants, F2 and G1, found in CC87 had a duplicated alanine residue at position 437. The D7 variant, also detected in CC87, was truncated by seven amino acid residues due to the presence of C2022A mutation resulting in a stop codon. The most prevalent PBP4 variant, F1 (58 isolates of ST6), was mainly identified in 55 isolates obtained from a single hospital setting, POZ1 in 2015 and 2016. All these isolates were resistant to vancomycin and carried the vanB gene, which strongly suggested a VRE-PRASEF outbreak in POZ1.
Clumping assay, detection of pheromone-responsive plasmids associated sequences and conjugation experiments
In the investigated group, 24 isolates (17.6%) were positive in clumping assay. Detection of sequences specific for pheromone-responsive plasmids showed, however, that these plasmids likely are more prevalent, considering the observed frequency of the rep9pAD1/pTEF2/pCF10 (98.5%), relpAD1 (99.5%), rep8pAM373 (11.0%), relpCF10 (11.0%), par (35.3%), asa1 (98.5%), cylA (32.3%), bee1 (4.4%), and bee23 (2.2%). None of the investigated isolates carried asa373. Conjugational transfer of penicillin-resistance determinant was attempted for 33 isolates either positive in clumping assay and/or carrying the highest number of all investigated pheromone-responsive-associated genes, that is, 6 of 10; however, penicillin-resistant E. faecalis transconjugants were not obtained.
Discussion
Enterococci, especially E. faecalis and E. faecium, nowadays represent important nosocomial pathogens of humans. Acquisition of antimicrobial resistance constitutes a significant step in the adaptation of enterococci to persistence in hospitals. 44 The emergence of novel resistance phenotypes in these bacteria, especially to important anti-enterococcal drugs, is of great concern. The NRCST collects PRASEF isolates from all clinical materials and while such isolates are as yet sporadically reported, and in our study were obtained mostly from noninvasive infections, considering the importance of β-lactams in therapy of enterococcal infections this phenomenon deserves to be thoroughly investigated.
Hospital-associated isolates of E. faecalis and E. faecium significantly differ from commensal strains and predominantly belong to distinct subpopulations of both species, that is, they represent so-called HiRECCs. One of the steps in the emergence of hospital-adapted E. faecium was the development of resistance to ampicillin, followed by the acquisition of vancomycin resistance, which positioned these bacteria among the most challenging nosocomial pathogens. It is possible that the similar scenario might be repeated by E. faecalis, with an acquisition of penicillin resistance as one of important adaptive features.
In the investigated group of isolates, penicillin-resistance emerged in a limited number of STs, which predominantly belonged to HiRECCs 6 and 87, two important CCs circulating in Polish hospitals. 17 The diversity of mutations conferring this phenotype and the fact that almost all STs were typically associated with more than a single variant of PBP4 clearly indicates several independent events of resistance acquisition by these HiRECCs. At the same time, we did not find PRASEF among representatives of other CCs, such as 16, 21, 30, and 40, which also occur among hospitalized patients. 17 Considering that CCs 6 and 87 are almost limited to the hospital settings, in contrast to other clones circulating also in the community, the selective pressure exerted on strains representing CCs 6 and 87 by use of β-lactams in hospitals may be the strongest. Similar scenario may be also true for CC88, which was not previously identified as HiRECC, but its isolates appear to be limited to hospital settings in Poland. 45 Not much information is available about the clonality of PRASEF isolates in other countries; penicillin resistance was observed among ST6 isolates in Denmark 46 and in an invasive isolate of ST28 (CC87) in Malaysia 47 while in Brazil this phenotype occurred in another hospital-associated CC9. 11
Penicillins and other β-lactams interfere with the synthesis of cell-wall peptidoglycan by inhibiting PBPs (D,D-transpeptidases) that catalyze cross-linking of peptidoglycan peptide side chain. Enterococci exhibit an intrinsic low susceptibility to β-lactam antibiotics due to the production of PBPs with low affinity for the drug, PBP5 in E. faecium and PBP4 in E. faecalis. 48
In the present study, further increase in MIC values of penicillin probably resulted from the PBP4 overproduction and/or mutations lowering PBP4 affinity to penicillin. All investigated isolates had nucleotide alteration(s) within the pbp4 promoter region. The most prevalent change, a single nucleotide deletion of the A residue 8 bases upstream of a putative −35 region (119 isolates, 87.5%), was present in all isolates of CC6 and CC87. This deletion was previously shown to contribute to PBP4 overproduction and penicillin resistance. 8 The importance of two additional observed mutations, A/T 13 bases upstream of a putative −35 region in ST832 (CC87) and C/T two bases upstream of a putative −10 region in CC88 for the pbp4 expression remains to be established.
Most reports on β-lactam resistance in E. faecalis associate the lack of susceptibility to these antimicrobial agents with amino acid changes in PBP4. In the present study, 136 investigated isolates carried 30 different PBP4 variants. Only seven isolates of CC6 contained the wild-type A variant, these isolates, however, as mentioned above presumably overproduce PBP4.
Amino acid changes of PBP4 were almost exclusively observed in the C-terminal penicillin binding domain, which contains three conserved motifs, 424SXXK427, 482SDN484, and 619KTG621, defining the transpeptidase active-site. 49 The B3 variant with a single substitution D573E as a possible determinant of penicillin resistance had been previously identified in PRASEF isolates of HiRECC 9. 11 Besides the B3 variant, the D573E change was also present in D9 and E5 variants in CC88. None of additional amino acid substitutions characteristic for these variants did not, however, influence penicillin and ampicillin MIC values, thus they may represent sequence polymorphisms.
While none of amino acid alterations observed in our study directly affected three PBP4 conserved motifs, several amino acid changes such as T418A, L475Q/M, A488T and A618G were located in their proximity. The A617T alteration next to the 619KTG621 motif was shown to destabilize the PBP4 transpeptidase domain, which most likely led to a reduced β-lactam affinity of this variant and resistance to penicillin, ampicillin, and imipenem of E. faecalis isolate 8 and the A618G change observed in our study may have a similar effect. Similarly, the L475M amino acid substitution, located next to the 482SDN484 motif presumably affects penicillin resistance, since it was associated with faropenem insusceptibility. 10 Several other identified amino acid changes in the PBP4 transpeptidase domain, which were not adjacent to PBP4 conserved motifs might also affect β-lactam-resistance, as it was demonstrated for P520S and Y605H amino acid substitutions, resulting in ampicillin and imipenem resistance. 10 In our study, Y605H was identified in several PBP4 variants.
Apart from substitutions, for the first time we observed more profound PBP4 changes, such as duplication of an amino acid residue and truncation at the C-terminus. Duplications involving single amino acid residues (466′S, 466′D, and 568′L) and a dipeptide (433′GL) were described in PBP5 of penicillin-resistant E. faecium. 50 The PBP4 truncation affects the C-terminal helix of the protein (aa 657–680), which together with the “lid” (445–473 amino acids) enclose the PBP4 active site in a deep cleft. 51 Taken together, penicillin-resistance of Polish PREASEF is the most likely the effect of amino acid changes in PBP4 combined with increased quantities of this protein, as it was previously proposed by Rice et al. 8
Pheromone-responsive plasmids play important role in the evolution of E. faecalis toward nosocomial pathogen contributing to the spread of antimicrobial resistance genes and putative virulence determinants. In this study, only limited number of investigated isolates (17.6%) phenotypically manifested the presence of pheromone-responsive plasmids, yielding positive results in clumping assay. Nevertheless, sequences specific for such plasmids were detected in nearly all isolates, which implies that pheromone-responsive plasmids were much more prevalent in the investigated group.
Pheromone-responsive plasmids were shown to be able to mobilize chromosomal genes during conjugation 25 and conjugational transfer of chromosomally located pbp5 between strains was reported in E. faecium. 52 Thus, we attempted to achieve such transfer in E. faecalis, using selected donors, active in clumping assay and/or carrying several genes specific for pheromone-responsive plasmids. Although under the laboratory conditions this attempt was not successful, this does not preclude the possibility of such events in natural environment of these bacteria.
Apart from penicillin resistance, investigated isolates commonly displayed resistance to several other drugs, a typical feature of HiRECCs. 17 Vancomycin resistance was predominantly associated with the presence of vanB and all these isolates carried F1 variant of PBP4, belonged to ST6 and were mostly identified in a single hospital setting, consistent with an outbreak situation. Thus, the observed increase of PRASEF was in a big part due to the presumable outbreak of vanB-PRASEF of CC6 in the POZ1 hospital setting.
The recent study showed that penicillin resistance in E. faecalis is acquired by mutations in pbp4 and its promoter, and that this phenomenon is specific for HiRECCs. Although PRASEF are still infrequently isolated both in Polish hospitals and worldwide, the emergence of penicillin-resistance in hospital-associated E. faecalis clones is a dangerous phenomenon, especially when combined with the acquisition of resistance to vancomycin and other drugs.
Footnotes
Acknowledgment
We thank all Polish microbiologists for providing isolates used in this study.
Disclosure Statement
No competing financial interests exist.
Funding Information
The study was financed by the grant UMO-2014/13/N/NZ7/00843 from the Polish National Science Centre (NCN). It was also supported by the SPUB MIKROBANK from the Polish Ministry of Science and Higher Education, and Narodowy Program Ochrony Antybiotyków (NPOA) from the Polish Ministry of Health.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.