
Abstract
Linezolid (LNZ) is one of the most important antimicrobial agents against infections caused by gram-positive bacteria, including enterococci. In a layer operation system, antimicrobial resistance can be transferred to commercial layers via the fecal-oral route. This study investigated the presence and distribution of LNZ-resistant Enterococcus faecalis and Enterococcus faecium in a layer operation system. Among 117 E. faecalis and 154 E. faecium, 10 (8.5%) E. faecalis and 5 (3.2%) E. faecium isolates showed resistance to LNZ and chloramphenicol, and they exhibited multidrug resistance against 5 or more classes of antimicrobial agents. Among the resistant isolates, 9 (90.0%) and 2 (20.0%) E. faecalis harbored optrA and cfr genes, respectively. The optrA and fexA genes were not detected in five LNZ-resistant E. faecium. None of the 15 LNZ-resistant isolates harbored the fexA gene, and no mutations were observed in the genes encoding domain V of 23S ribosomal RNA (rRNA) and ribosomal proteins L3 (rplC) and L4 (rplD). Transferability was identified in three of the nine optrA-positive LNZ-resistant isolates. The tetM, tetL, and ermB genes were cotransferred with the optrA gene in all optrA-positive transconjugants. The results indicate that optrA is well-distributed in E. faecalis, implying a greater level of transferability. Thus, enhanced surveillance efforts are needed to monitor the emergence and spread of optrA in enterococci in layer operation system.
Introduction
Linezolid (LNZ), which was approved for clinical use in 2000 in the United States and Korea, is one of the most important antimicrobials used to treat infections caused by multidrug-resistant gram-positive bacteria, methicillin-resistant Staphylococcus aureus, and vancomycin (VAN)-resistant enterococci.1,2 However, resistance against LNZ has raised concern regarding possible issues in the treatment of enterococcal infections in humans. 3 Although LNZ resistance is still uncommon among enterococci, it has emerged in recent years in human and animal isolates from many countries.4–6
Resistance in enterococci is mediated by point mutations in the central loop of domain V of 23S ribosomal RNA (rRNA) and ribosomal proteins L3 (rplC) and L4 (rplD); other mechanisms include the presence of the multiresistance gene cfr, which encodes a 23S rRNA methyltransferase.1,5 In addition, Wang et al. 7 reported a novel gene, optrA, which encodes an ATP-binding cassette transporter that confers resistance against oxazolidinones and phenicols. Recently, optrA has been regularly identified in Enterococcus faecalis and Enterococcus faecium isolates from food-producing animals such as chickens, ducks, and pigs from China, 5 Colombia, 8 and Korea.9,10
E. faecalis and E. faecium are present in the gastrointestinal tract of humans and animals as well as in soil, on plants, and in water. E. faecalis and E. faecium are ubiquitous in nature and resistant to unfavorable environmental conditions.11,12 In addition, E. faecalis and E. faecium have a high ability to acquire resistance genes from the same or different species via transferable plasmids or transposons, which allows them to have a role as antimicrobial resistance indicators. 13
In Korea, the poultry layer operation system has a pyramidal structure, in which the grandparent stock (GPS) is at the top, followed by the parent stock (PS) and commercial chickens (CC), which produce eggs at the bottom of the pyramid. 14 The presence of antimicrobial-resistant bacteria in fecal material from the PS can be disseminated into the environment via the intestine and transferred to chicks through hatcheries, which can serve as bacterial reservoirs. 13 Although many studies have investigated the prevalence and characteristics of antimicrobial-resistant bacteria in the poultry industry,15–17 little is known about the prevalence and characteristics of LNZ-resistant enterococci in poultry. Thus, this study aimed to investigate the presence and distribution of LNZ-resistant E. faecalis and E. faecium isolates from a layer operation system in Korea.
Materials and Methods
Sampling
Between 2018 and 2019, 230 dust and 230 feces samples were obtained from 1 GPS farm (5 flocks), 16 PS farms (80 flocks), and 6 CC farms (30 flocks) of the biggest one-layer operation system in Korea. All samples were collected as two dust and two feces samples per flock once when the chickens were 20 weeks old, per to the standards set by the National Poultry Improvement Plan. 18 For each flock, dust samples were swabbed from dusty spots by using a gauze (12 layered, 10 × 10 cm; Hans Medical, Busan, Korea) that had been moistened with sterile skim milk. Fecal samples (∼10 g fresh fecal droppings) were also collected from random locations within the flock.
Bacterial isolation
Microbiological analyses were performed according to the National Antimicrobial Resistance Monitoring Program as described by the Animal and Plant Quarantine Agency 19 in Korea. The samples were individually inoculated into 225 mL buffered peptone water (BD Biosciences, Sparks, MD) and incubated for 18–24 h at 37°C. Preenriched buffered peptone water was inoculated into Enterococcosel broth (BD Biosciences) at 1:10 ratio and incubated at 37°C for 18 to 24 h. After enrichment, the Enterococcosel broth that had turned black was streaked onto Enterococcosel agar (BD Biosciences) plates and incubated at 37°C for 18–24 h. Three to five presumptive-positive colonies were inoculated onto blood agar, and the resulting clones were identified as E. faecalis or E. faecium by using polymerase chain reaction (PCR) method previously described.20,21 If some isolates of the same origin showed the same antimicrobial susceptibility patterns, only one isolate was randomly chosen and included in this study.
Antimicrobial susceptibility test
All isolates were investigated for resistance by determining minimum inhibitory concentrations (MICs) of 16 antimicrobial agents, including ampicillin (≥16 μg/mL), chloramphenicol (CHL, ≥32 μg/mL), ciprofloxacin (CIP, ≥1 μg/mL), daptomycin (DAP, ≥8 μg/mL), erythromycin (ERY, ≥8 μg/mL), florfenicol (FFN, ≥16 μg/mL), gentamicin (GEN, ≥1,024 μg/mL), kanamycin (KAN, ≥1,024 μg/mL), LNZ (≥8 μg/mL), salinomycin (≥16 μg/mL), quinupristin/dalfopristin (SYN, ≥4 μg/mL), streptomycin (STR, ≥1,024 μg/mL), tetracycline (TET, ≥16 μg/mL), tigecycline (TGC, ≥0.25 μg/mL), tylosin tartrate (TYLT, ≥32 μg/mL), and VAN (≥32 μg/mL). MICs were measured by applying the broth microdilution method using the commercially available Sensititre® panel KRVP2F (TREK Diagnostic Systems, West Sussex, United Kingdom) according to the manufacturer's instructions. Results were interpreted according to Clinical and Laboratory Standards Institute (CLSI) guidelines. 22 When CLSI breakpoints were unavailable, MIC results were interpreted according to the National Antimicrobial Resistance Monitoring Program. 19 E. faecalis ATCC 29212 was used as the control strain for the MIC tests. Multidrug resistance (MDR) was defined as acquired nonsusceptibility to three or more antimicrobial classes. 13
Detection of antimicrobial resistance genes
The novel oxazolidinone and phenicol resistance gene, optrA, was amplified using primers as described by Wang et al. 7 The presence of the MDR gene, cfr, and the FFN resistance gene, fexA, was determined using primers as previously described. 23 All isolates were tested for the presence of resistance genes against macrolides (ermA, ermB, and mef), tetracyclines (tetL, tetM, and tetO), and aminoglycosides [aac(6′)-Ie–aph(2″)-Ia, aph(2″)-Ib, aph(2″)-Ic, aph(2″)-Id, ant(3″)-Ia, and ant(6)-Ia]. The primers and conditions used for detecting resistance genes were as previously described,24–27 and the primers are listed in Supplementary Table S1.
Screening for mutations
The presence of mutations in the genes encoding domain V of 23S rRNA, and ribosomal proteins L3 (rplC) and L4 (rplD) was investigated as previously described.4,28 The nucleotide and amino acid sequences of rplC, rplD, and domain V of 23S rRNA gene were compared with those of the wild-type LNZ-susceptible E. faecalis ATCC 29212 strain (GenBank accession No. CP008816.1) and the E. faecium DO strain (GenBank accession No. CP003583.1). The sequences were confirmed with those in the GenBank nucleotide database by using the Basic Local Alignment Search Tool program available through the National Center for Biotechnology Information website (www.ncbi.nlm.nih.gov/BLAST).
Conjugation assay
To determine the transferability of optrA, a conjugation assay was performed with broth mating using rifampicin- and fusidic acid-resistant E. faecalis FA2-2 recipient strain as previously described. 29 Transconjugants were selected on a brain heart infusion agar plate (BD Biosciences) with 50 μg/mL rifampin (Merck, St. Louis, MO), 100 μg/mL fusidic acid (Merck), and 10 μg/mL LNZ (Merck). Mating experiments were repeated a minimum of three times. All transconjugants selected were confirmed by the presence of optrA by performing PCR, and MICs were investigated by using antimicrobial susceptibility tests.
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) of optrA-positive E. faecalis and E. faecium isolates was performed by digesting genomic DNA with SmaI restriction enzyme (Takara, Shiga, Japan) according to a standard protocol of the Center for Disease Control and Prevention using a CHEF-MAPPER apparatus (Bio-Rad Laboratories, Hercules, CA), as previously described. 10 Gel images were analyzed by using BioNumerics software, version 4.0 (Applied Maths, Sint-Martens-Latem, Belgium).
Statistical analysis
Statistical analysis was performed using the Statistical Package for Social Science version 25 (IBM, Seoul, Korea). The chi-squared tests were used to compare the prevalence of drug-resistant isolates between production stages as well as between E. faecalis and E. faecium. A p-value of <0.05 was considered statistically significant.
Results
Prevalence of E. faecalis and E. faecium
The prevalence of E. faecalis and E. faecium in the layer operation system is shown in Table 1. A total of 117 E. faecalis and 154 E. faecium isolates were obtained from the GPS (8 and 10 isolates, respectively), PS (74 and 103 isolates, respectively), and CC (35 and 41 isolates, respectively). Among them, 10 (8.5%) E. faecalis and 5 (3.2%) E. faecium isolates showed resistance against LNZ and CHL. Although no enterococci isolated from GPS showed resistance against LNZ, 5.6% and 6.6% of enterococci from PS and CC, respectively, showed resistance to LNZ. There were no significant differences in the prevalences of LNZ and CHL resistance isolates among GPS, PS, and CC. Of note, E. faecalis (8.5%) showed higher resistance against LNZ than did E. faecium (3.2%), although there was no significant difference between the E. faecalis and E. faecium resistances.
Resistance to Linezolid of Enterococcus faecalis and Enterococcus faecium Isolated from Layer Operation System
There were no significant differences between E. faecalis and E. faecium.
All isolates showed the resistance to LNZ and CHL, simultaneously. There were no significant differences between GPS, PS, and CC.
CC, commercial chicken; CHL, chloramphenicol; GPS, grandparent stock; LNZ, Linezolid; PS, parent stock.
Multidrug resistance phenotypes of LNZ-resistant enterococci
Resistance patterns of 10 E. faecalis and 5 E. faecium LNZ-resistant isolates from the layer operation system are shown in Table 2. A total of 15 enterococci showed MDR against over 5 classes of antimicrobial agents. In particular, LNZ-resistant isolates also were highly resistant to CHL (100%), CIP (100%), DAP (100%), SYN (93.3%), TYLT (93.3%), ERY (80.0%), and FFN (80.0%). The commonest pattern detected was CHL-CIP-DAP-ERY-FFN-GEN-KAN-LNZ-STR-SYN-TET-TGC-TYLT.
Distribution of Multidrug Resistance Patterns of 15 Linezolid-Resistant Enterococcus faecalis and Enterococcus faecium Isolated from Layer Operation System
n = No. of Enterococcus spp. isolated in this study.
No. of isolates shown resistance (%).
CIP, ciprofloxacin; DAP, daptomycin; ERY, erythromycin; FFN, florfenicol; GEN, gentamicin; KAN, kanamycin; SAL, salinomycin; STR, streptomycin; SYN, quinupristin/dalfopristin; TET, tetracycline; TGC, tigecycline; TYLT, tylosin tartrate.
Characterization of LNZ-resistant E. faecalis and E. faecium
Characteristics of the 15 LNZ-resistant Enterococcus spp. are shown in Table 3. Among the 10 E. faecalis and 5 E. faecium LNZ-resistant isolates, 9 (90.0%) and 2 (20.0%) E. faecalis isolates harbored optrA and cfr genes, respectively. The fexA gene was not detected in any of the LNZ-resistant isolates. Furthermore, all 15 LNZ-resistant isolates carried no mutation in the genes encoding domain V of 23S rRNA and ribosomal proteins L3 (rplC) and L4 (rplD). The MIC of LNZ was between 8 and ≥16 μg/mL, while that of CHL was ≥32 μg/mL. Two E. faecalis and one E. faecium LNZ-resistant isolates were susceptible to FFN (≤8 μg/mL). Other antimicrobial resistance genes were also identified: aac(6″)Ie-aph(2″)-la+ant(6)+ermB+tetL+tetM (two E. faecalis); aac(6″)Ie-aph(2″)-la+ant(6)+ermB (three E. faecalis); aac(6″)Ie-aph(2″)-la+ermB+tetL+tetM (three E. faecalis); mef+tetL+tetM (one E. faecium); and tetL+tetM (two E. faecium). Moreover, two E. faecalis and two E. faecium isolates carried no other resistance genes.
Characteristics of 15 Linezolid-Resistant Enterococcus faecalis and Enterococcus faecium Isolates from Layer Operation System
—, Not detected; FFN, florfenicol; MICs, minimum inhibitory concentrations; rRNA, ribosomal RNA.
Characteristics of optrA–positive E. faecalis
The genetic relatedness and transferability of nine optrA-positive and LNZ-resistant E. faecalis isolates are shown in Fig. 1. Based on the similarity value of 0.85, the nine optrA-positive E. faecalis isolates represented five arbitrary (designated as I through V) pulsotypes. Three optrA-positive transconjugants with cotransferability of the tetM, tetL, and ermB genes were confirmed.
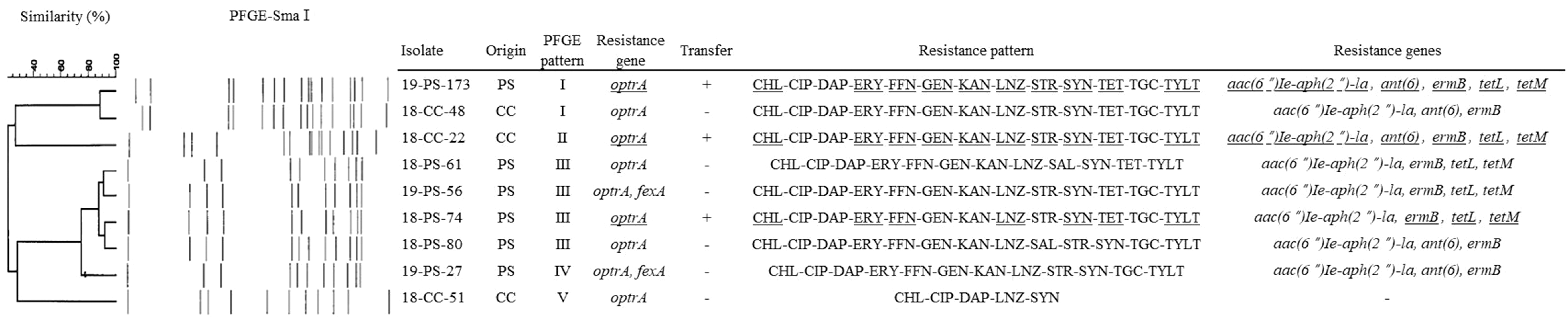
Dendrogram of SmaI-PFGE patterns of nine Enterococcus faecalis with optrA gene. E. faecalis isolates showing similarities of <85% were considered to be unrelated. Underline indicated that was found in the transconjugant strains. +, positive; −, negative; CC, commercial chicken; CHL, chloramphenicol; CIP, ciprofloxacin; DAP, daptomycin; ERY, erythromycin; FFN, florfenicol; GEN, gentamicin; KAN, kanamycin; LNZ, linezolid; PFGE, pulsed-field gel electrophoresis; PS, parent stock; SAL, salinomycin; STR, streptomycin; SYN, quinupristin/dalfopristin; TET, tetracycline; TGC, tigecycline; TYLT, tylosin tartrate.
Discussion
Although LNZ is not used in the treatment of food-producing animals, LNZ-resistant enterococci have been recently observed in duck feces and carcasses in Korea.9,10 Enterococci from the poultry industry generally show high resistance to variety antimicrobials,30,31 and this report is the first report on the characteristics of LNZ-resistant enterococci in a layer operation system in Korea. In this study, 5.5% (15/271) of the enterococci isolates were resistant to LNZ and CHL. Although no enterococci isolated from GPS showed resistance against LNZ, the rate was higher than the previously reported prevalence of 0.16% of enterococci in food-producing animals in Korea. 9 In particular, 5.6% of enterococci from PS showed resistance against LNZ; therefore, the presence of LNZ-resistant enterococci in PS is problematic as commercial layers can be adversely affected through the fecal-oral route from hatcheries to hatcheries, due to the pyramidal nature of the layer operation system.
Miller et al. 32 demonstrated that LNZ resistance in enterococci was mainly due to the G2576T mutation in the 23S rRNA gene and mutations in rplC and rplD. But in this study, all LNZ-resistant enterococci isolates lacked mutations in the domain V of 23S rRNA gene and in rplC and rplD. Tamang et al. 9 and Zhang et al. 33 reported that LNZ-resistance among enterococci commonly develops as a result of genetic transfer. The novel optrA gene, which codes for an ABC transporter that confers transferable resistance against oxazolidinones and phenicols, was first detected in enterococci isolates of animal origin in China. 7 Moreover, optrA-positive isolates cocarried cfr, which codes for a 23S rRNA methyltransferase that confers resistance against oxazolidinones, phenicols, lincosamides, pleuromutilins, and streptogramin A. 34 In this study, 9 (90.0%) and 2 (20.0%) E. faecalis among 15 LNZ-resistant isolates harbored the optrA and cfr genes, respectively. In particular, these genes were more frequently detected in E. faecalis than in E. faecium. Previous studies also reported that the optrA gene is more frequently seen in E. faecalis than in E. faecium, and more commonly observed in food-producing animals than in humans.7,33 These surveillance data suggested that optrA is commonly disseminating in E. faecalis, implying a high level of transferability.35,36
The optrA gene has been detected in the chromosome and plasmids of enterococci of human and animal origin.5,7 In this study, 3 among the 15 LNZ-resistant enterococci also transferred with macrolides (ermB), tetracyclines (tetL and tetM), and aminoglycosides [aac(6′)-Ie–aph(2″)-Ia and ant(6)-Ia] resistance genes. Previous studies reported that LNZ-resistant enterococci are transferred with macrolide, streptogramin, tetracyclines, and phenicols resistance, along with LNZ resistance.5,10 Therefore, the mobile genetic element, carrying not only LNZ resistance genes but also other additional resistance genes, is a matter of concern, since they might facilitate coselection of the optrA gene.10,33
The results of the PFGE analysis showed that nine optrA-positive and LNZ-resistant E. faecalis isolates were divided into five pulsotypes. According to Tamang et al., 9 PFGE results demonstrated that enterococci carrying the optrA gene isolated from animal origin were diverse, but some enterococci were genetically homogeneous, similar to the present results. Noteworthy, in the present study, two different isolates from PS and CC each, showed the same PFGE pattern.
The emergence detection of LNZ-resistant enterococci carrying optrA gene in a transferable plasmid, linked to resistance genes against antimicrobials commonly used in poultry layer operation system suggest that this gene has a role in the coselection of MDR bacteria. Also, LNZ-resistant enterococci carrying the optrA gene can lead to dissemination via horizontal or vertical transfer within the layer operation system, which has a pyramidal structure. In addition, the results suggest that the dissemination of optrA gene can lead to layer hens that can contribute to the transmission of this gene to humans through the food chain. The transfer of this gene may significantly limit the treatment of infections caused by multidrug-resistant gram-positive cocci. Thus, an enhanced surveillance effort is needed to monitor the emergence and spread of optrA in enterococci in the layer operation system.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.