
Abstract
To analyze the characteristics of fosA and fosA3 in Enterobacter cloacae isolated from aspirated and catheterized urine culture specimens of companion pets in Taiwan. A total of 19 E. cloacae isolates from pets with urinary tract infection were screened for the presence of fosA, fosA3, and fosC2 and for the genetic context of them by PCR amplification and sequencing. The transferability, resistance phenotypes, plasmid replicon typing properties and genetic environments of fosA- and/or fosA3-positive strains were characterized. Five E. cloacae isolates were positive for fosA and three coharbored fosA and fosA3. No fosC determinant was detected. Transconjugants of fosA3 were successfully acquired, while the acquisition of fosA transconjugants was failed. The minimum inhibitory concentrations (MICs) of the three fosA3-positive isolates and their transconjugants were ≥256 mg/L, whereas the MICs of the five fosA-positive isolates ranged from 64 mg/L to 256 mg/L. Three plasmid replicons (InCFrepB, InCL/M, and InCHI2) were identified in fosA- and fosA3-positive E. cloacae isolates. Different genetic contexts lay in the downstream region of fosA and fosA3, respectively. Eight distinct patterns based on the similarity value of more than 80% were typed for all the 8 fosA-positive isolates. In conclusion, the fosA concomitant with fosA3 were found in E. cloacae isolates. The fosA3 not only exhibits stronger activity of inactivating fosfomycin than fosA but also possesses stronger potential to spread than fosA. Different genetic backgrounds exist in these fosA- and fosA3-positive isolates, and different mobile elements may confer the dissemination of fosA and fosA3.
Introduction
Fosfomycin, formerly called phosphonomycin or phosphomycin, is a phosphonic acid derivative (cis-1,2-epoxypropyl phosphonic acid) containing an epoxide ring and a carbon−phosphorus bond. 1 It enters susceptible bacterial cells by active transport through the L-α-glycerophosphate (α-GP) uptake system (GlpT) or the glucose-6-phosphate (G-6-P) uptake system (Uhp) and irreversibly inhibits peptidoglycan biosynthesis through the inactivation of the UDP-N-acetylglucosamine (UDP-GlcNAc) enolpyruvoyl transferase by acting as an analog of phosphoenolpyruvate, subsequently disrupting cell wall synthesis. 2 Oral fosfomycin was mainly used to treat uncomplicated urinary tract infections (UTIs) and bacterial prostatitis caused by multidrug-resistant gram-negative (MDR-GNB); besides, intravenous fosfomycin has been mostly used in combination with other agents for treating various type of complicated, severe MDR-GNB infections. 3
With the rapidly increasing of bacterial resistance in clinic recently, an increasing number of antibiotics lost their superiorities in treatment of the infections caused by resistant bacteria such as methicillin-resistant Staphylococcus aureus, carbapenem-resistant Pseudomonas aeruginosa, and extended-spectrum beta-lactamase-producing gram-negative bacteria. 4 Fosfomycin has become alternative choice in treatment of resistant microbial infections for its unique bactericidal mechanism of action, broad antimicrobial spectrum in both gram-negative and gram-positive bacteria, good tolerability, and synergistic or additive effects with other therapeutic antibiotics in vivo or in vitro.5–7 However, resistance to fosfomycin still occurred and developed in recent years.
Currently, it has been reported that the fosfomycin resistance in bacteria mainly involved four kinds of mechanisms, 8 (1) the mutation and overexpression in murA gene encoding the covalently attaching site of fosfomycin result in the fosfomycin resistance8,9; (2) the mutations in the encoding (or regulatory) genes of uptake transporters for fosfomycin can decrease the synthesis of the uptake transporters and cause the structural or functional abnormalities of the uptake transporters, leading to the impairment of fosfomycin uptake10,11; (3) various fosfomycin-modifying enzymes, including FosA, FosB, FosC, FosX, FosK, FosD(Genbank accession number, KC989517 and NC_010419), FosE (Genbank accession number, AB901041), FosI (Genbank accession number, CP003505), and FomA and FomB located on plasmid or chromosome in antibiotic-producing bacteria and clinical bacteria, including Escherichia coli, Klebsiella pneumoniae, Acinetobacter spp., Enterobacter cloacae, Enterobacter aerogenes, P. aeruginosa, Salmonella enterica, Enterococcus faecium, S. aureus, and S. epidermidis, inactivate fosfomycin by exerting their glutathione-S-transferase, cysteine-S-transferase, or other enzymatic activity12–17 ; and (4) heteroresistance to fosfomycin was probably multifactorial, and heteroresistance to fosfomycin has been described in 10 out of 11 Streptococcus pneumoniae strains, and in MDR and non-MDR P. aeruginosa clinical isolates from Australian hospitals. 8
Recently, several fosfomycin-resistant determinants, including fosA, fosA2, fosA3, fosA5, fosC2, and fosK located on plasmids, transposons, integrons, insertion sequence IS26 in E. coli, K. pneumoniae, and S. enterica isolated from human, livestocks, food-borne animals,18–23 were reported and therefore aggravated the burden of the bacterial infectious treatment. In this study, 8 E. cloacae isolates from 19 aspirated or catheterized urine culture specimens of companion pets in Taiwan were detected to harbor fosA and three of them were found carrying fosA3 simutaneously. To our knowledge, no report has ever described that fosA3 and fosA were concomitant in one isolate. So, it is necessary to explore their characteristics such as the resistance phenotypes, transferabilities, plasmid replicon typing properties, and genetic environments to supplement the resistant mechanism of fosfomycin-modifying enzymes.
Materials and Methods
Bacterial strains
A total of 19 E. cloacae strains were randomly isolated from aspirated urine cultures of companion pets with UTIs [including dogs (n = 10) and cats (n = 9)] at the Veterinary Teaching Hospital of National Taiwan University from 2010 to 2013. These strains were identified using Biomerieux VITEK® 2 Compact Automatic Microbial Analyzing system and confirmed by 16S ribosomal RNA sequencing. E. coli C600 (resistant to streptomycin, Strepr) was used as recipient in the conjugation test.
Identification of fosfomycin-resistant determinants and the coexistent-resistant genes
The fosfomycin-modifying enzyme genes fosA, fosA3 and fosC2 and their genetic environment genes, including the upstream and downstream genes of fosA and fosA3, blaTEM, and blaCTX-M (group 1, 2, 8, 9, 25), and 16S rRNA methyltransferase genes (armA and rmtB) were detected by PCR amplifying and sequencing. The primers used in this study are listed in Table 1.
Primers Used to Detect the Fosfomycin-Resistant Determinants and the Their Environment Context
Conjugation experiments and antimicrobial susceptibility test
The transferability of fosfomycin-modifying enzyme genes was determined by using liquid conjugation experiments. 24 The fosA- and fosA3-positive isolates were used as donor bacteria, and E. coli C600 (resistant to streptomycin) was used as recipient. Transconjugants were selected on tryptic soy agar plates supplemented with fosfomycin (64 mg/L), glucose-6-phosphate (25 mg/L), and streptomycin (2,560 mg/L) and confirmed by PCR and sequencing. Microdilution method and agar dilution method were used to evaluate the minimum inhibitory concentrations (MICs) of fosA- and fosA3-positive isolates and their transconjugants to amikacin, gentamicin, streptomycin, ampicillin, cefazolin, doxycycline, sulfadimethoxine, and fosfomycin, respectively. Interpretive criteria from the Clinical and Laboratory Standards Institute guidelines (CLSI) and the European Committee on Antimicrobial Susceptibility Testing (EUCAST) for E. cloacae were used to analyze the results of MICs.
Plasmid replicon typing and ERIC-PCR
Plasmid incompatibility (Inc) groups were screened using multiplex-PCR based on replicon typing (PBRT). A total of 18 pairs of primers recognizing HI1, HI2, I1-Iγ, L/M, FIA, FIB, FIC, N, P, W, T, A/C, K, B/O, X, Y, F, and FIIA in 3 multiplex-PCR reactions were used. 25 All amplicons of these plasmid replicons were visualized by gel electrophoresis on an agarose gel stained with GoldView. Simple PCR was carried out to confirm the screening result. Genetic fingerprint of the eight fosA- and fosA3-positive E. cloacae isolates was typed by using Enterobacterial repetitive intergenic consensus sequence PCR (ERIC-PCR) with primers and amplified conditions as previously. 26 Weighted pair group method using arithmetic averages dendrogram were constructed on the basis of the 80% similarity analysis using the Quantity one 4.62 (Bio-Rad).
Results
Identification of fosfomycin-resistant determinants and the coexistent-resistant genes
As a result, 42.1% (8/19) specimens from three dogs (3/19, 15.8%) and five cats (5/19, 26.3%) were identified positive for fosA, 15.8% (3/19) specimens from three cats (3/19, 15.8%) were identified positive for fosA3 (Table 2). No fosC-positive isolates were detected. The three fosA3-positive isolates were found to be concomitant with fosA genes. Meanwhile, 16S rRNA methyltransferase gene armA were determined in one fosA-positive isolate and one fosA concomitance with fosA3-positive isolate, respectively. The β-lactamase determinant genes blaTEM and blaCTX-M-1 were detected in 88.9% (8/9) and 33.3% (3/9) fosA-positive isolates, respectively. Three fosA3 concomitant with fosA-positive isolates were found harboring blaTEM, while no fosA3 concomitant with fosA-positive isolates were found harboring blaCTX-M-1. Of the three fosA3 transconjugants, the β-lactamase determinant gene blaTEM was detected, while fosA gene and 16S rRNA methyltransferase gene armA, which were found in the parental isolates, were not detected.
Genetic Characteristics of the Eight fosA-Positive Enterobacter cloacae Isolates and Their Transconjugants
Conjugation experiments and antimicrobial susceptibility test result
For the three fosA3 coharboring with fosA-positive isolates, transconjugants with fosA3 determinants but without fosA gene were obtained successfully. Conjugative transfer frequencies of the three fosA3-positive isolates EC6, EC8, and EC18 were 2.3 × 10−6, 3.7 × 10−5, and 4.9 × 10−6, respectively. Moreover, no fosA transconjugants were acquired even though the transconjugation experiments were carried out repeatedly.
The MICs of eight antimicrobial agents for the eight E. cloacae isolates and the fosA3 transconjugants are listed on Table 3. The MICs of the three fosA3-positive isolates and their transconjugants to fosfomycin were ≥256 mg/L. While the MICs of EC2, EC3, EC4, EC5, and EC19 which carried the fosA to fosfomycin were 64 mg/L, 128 mg/L, 64 mg/L, ≥256 mg/L, and 128 mg/L, respectively. In the meanwhile, the fosA- and fosA3-positive isolates showed different resistance to amikacin, gentamicin, streptomycin, ampicillin, cefazolin, doxycycline, and sulfadiazine. All the fosA- and fosA3-positive isolates demonstrated resistance to ampicillin and sulfadiazine, and the MICs to them ranged from 128 mg/L to ≥256 mg/L and ≥512 mg/L, respectively. One fosA-positive isolate EC5 carried the armA gene showed high resistance to amikacin and gentamicin. Three fosA-positive isolates carried the blaCTX-M-1 genes displayed resistance to cefazolin, their MICs were 64 mg/L, 128 mg/L, and ≥256 mg/L, respectively. No fosA- and fosA3-positive isolates showed resistance to doxycycline. The fosA3 transconjugants showed high resistance to fosfomycin and ampicillin, MICs of them to fosfomycin and ampicillin were all ≥256 mg/L.
MICs (mg/L) of Eight Antimicrobial Agents for the Eight fosA-Positive Isolates and the Three fosA3 Transconjugants
The Transconjugants of EC06, EC08 and EC18.
AMK, amikacin; GEN, gentamicin; STRE, streptomycin; AMP, ampicillin; CFZ, cefazolin; DOX, doxycycline; SD, sulfadiazine; FOS, fosfomycin; CLSI, Clinical and Laboratory Standards Institute; EUCAST, European Committee on Antimicrobial Susceptibility Testing.
Plasmid replicon typing and ERIC-PCR
Three different plasmid replicons, including incompatibility (InC) FrepB, L/M, and HI2, were typed and verified in fosA- and fosA3-positive E. cloacae isolates (Table 2). Among the five fosA-harbored isolates, the Inc HI2 replicon was detected in four isolates, while the Inc L/M replicon type was determined in another isolate. Of the three fosA coharboring with fosA3 E. cloacae isolates, the IncFrepB replicon was found in two isolates, the Inc HI2 replicon was detected in another isolate. Meanwhile, two Inc FrepB replicons and one Inc HI2 replicon were detected in fosA3 transconjugants, respectively.
Five to nine discernible amplification bands with size ranging from 100 bp to 4,500 bp were obtained from eight fosA-positive E. cloacae isolates (Fig. 3). Also, eight distinct patterns based on the similarity value of more than 80% were typed. Four groups were artificially clustered based on the distance of genetic relationship (Fig. 3).
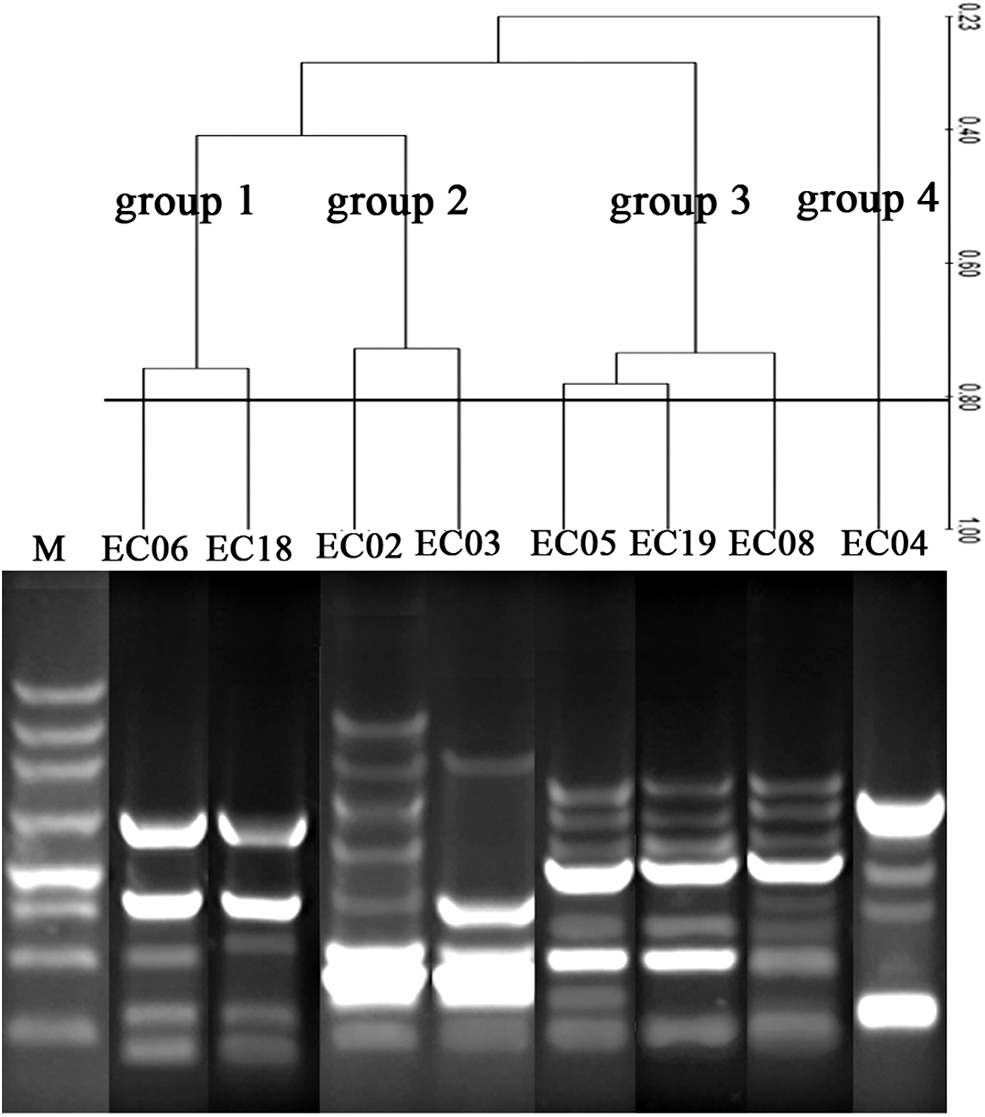
Phylogenic analysis of 8 fosA-positive E. cloacae isolates by using ERIC-PCR.
Genetic background of fosA and fosA3 in E. cloacae
Three types of genetic contexts existed in the three fosA3-positive E. cloacae isolates (Fig. 1). In the upstream region of fosA3, the IS26 transposase determinant was identified in all the fosA3 isolates. However, variable genetic elements were identified in the downstream region of the fosA3. In EC08, the length of the space region between the 3′ end of fosA3 and the IS26 transposase gene is 1,331 bp that contains a 519 bp length of orf1 encoding a hypothetical protein, a 552 bp length of orf2 encoding a CadC-like protein, and a 441 bp length of orf3 encoding a truncated TetR family transcriptional regulator. In EC6, the length of the space region between the 3′ end of fosA3 and the IS26 transposase gene is 590 bp, which only contains the 519 bp length of orf1. In EC18, the genetic context in the upstream region was not been amplified using the designed primers.
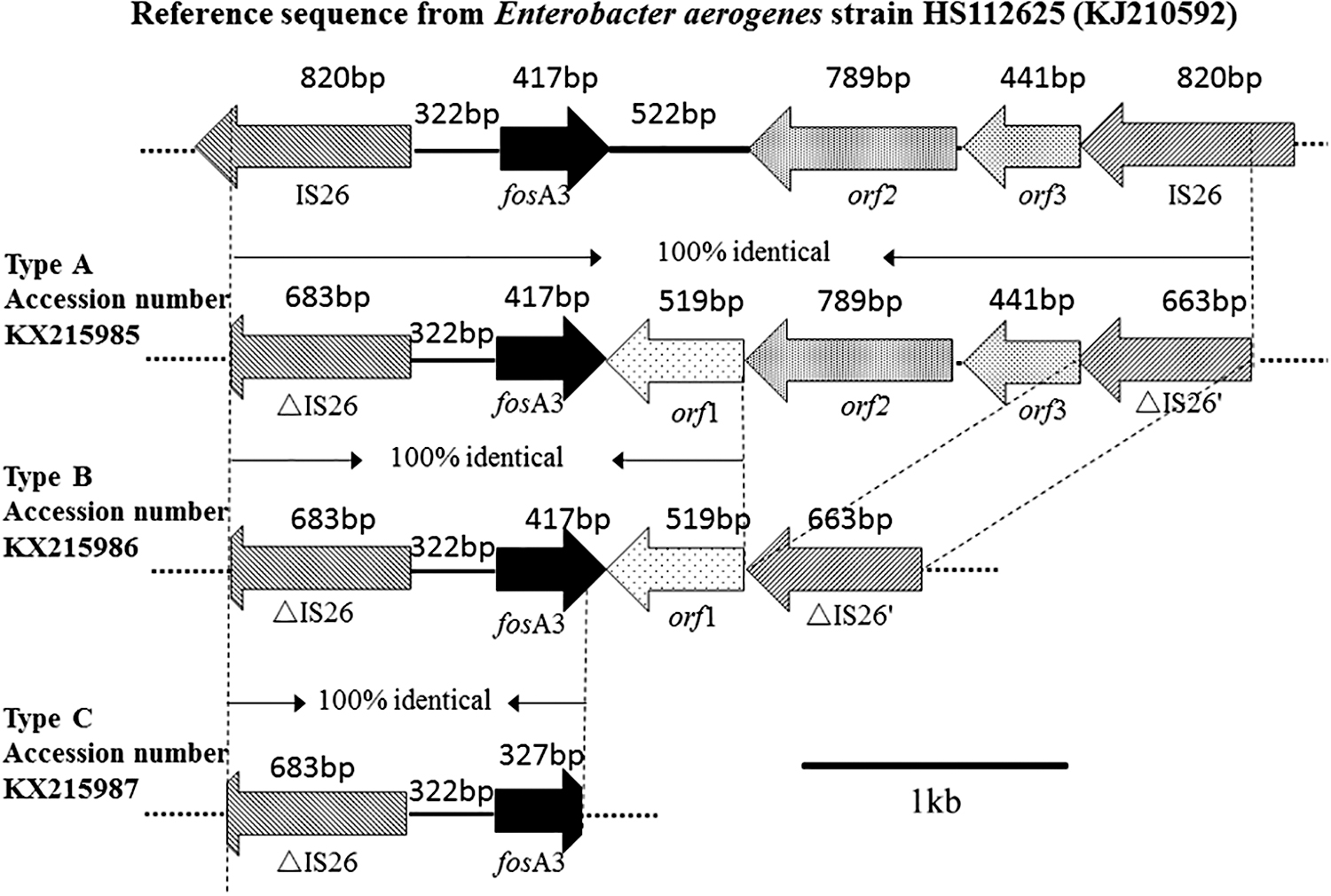
Schematic representation of the fosA3 genetic contexts in Enterobacter cloacae IS26, encoding mobile element IS26; △IS26, encoding part of IS26; orf1, encoding a hyperthetical protein but no conputative domain; orf2, encoding a CadC-like protein; orf3, encoding a truncated TetR family transcriptional regulator; △IS26′, encoding part of IS26.
For the eight fosA-positive isolates, two types of genetic contexts were identified (Fig. 2). In the upstream region of fosA, a 288 bp length of amplicon, which was identical to part of the truncated tryptophanyl tRNA synthetase gene that existed in E. cloacae strain ECNIH5 (CP009854) and Serratia marcescens transposon Tn2921 (FJ829469), was identified in all fosA-positive isolates. In the downstream region of fosA, a 1,008 bp length of amplicon encoding the lacI family transcriptional regulator and a 942 bp length of amplicon encoding a sugar-binding cellulose-like protein, which were similar to the 1,4-beta-xylanase in E. cloacae strain ECNIH5 and S. marcescens transposon Tn2921, were identified in EC5. In other seven fosA-positive isolates, only a 634 bp length of amplicon encoding part of lacI family transcriptional regulator were confirmed.
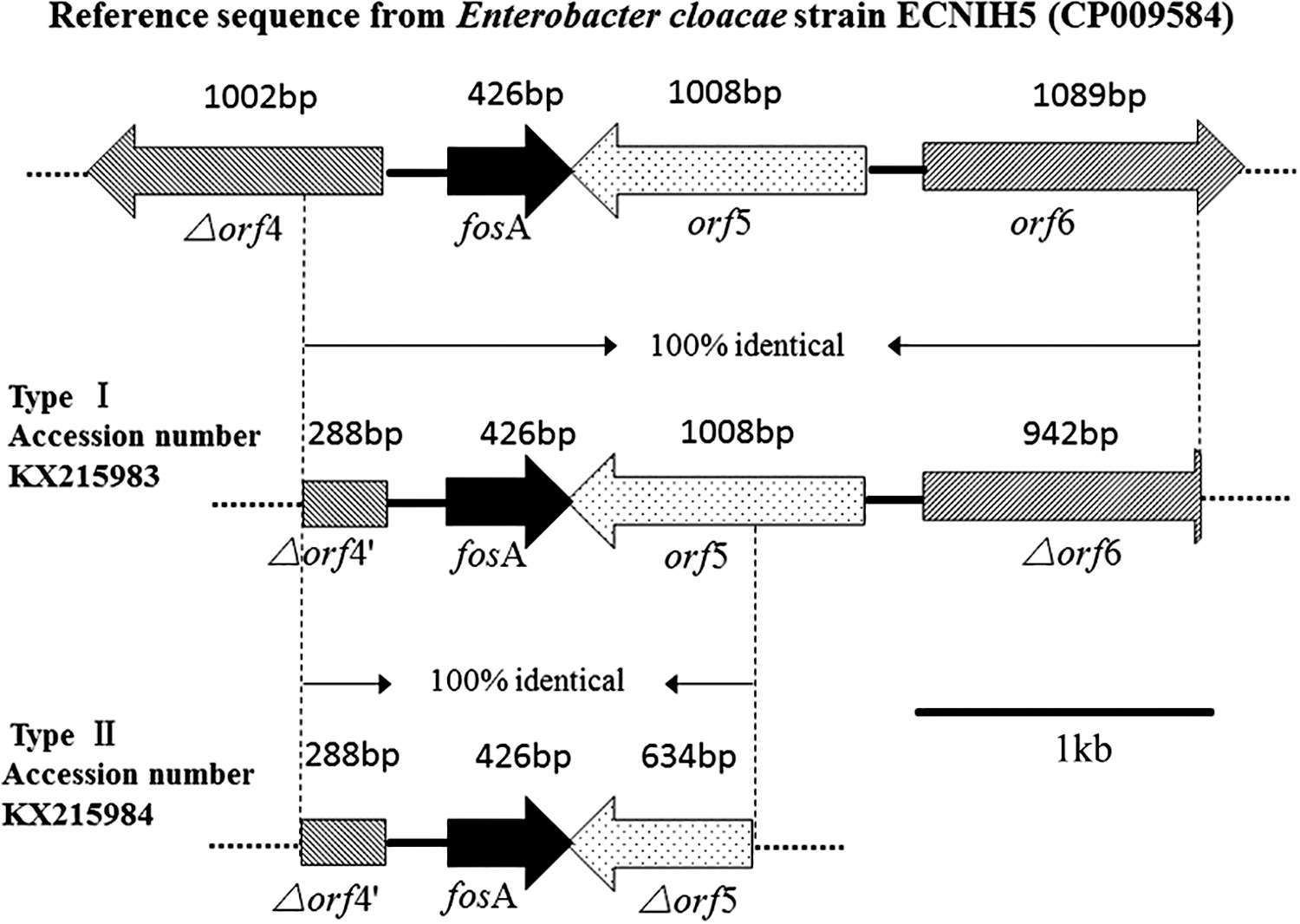
Schematic representation of the fosA genetic contexts in E. cloacae △orf4, encoding truncated tryptophanyl tRNA synthetase; △orf4′, encoding a nucleotidyl transferase conserved domain which was identical to part of the truncated tryptophanyl tRNA synthetase; orf5, encoding lacI family transcriptional regulator; △orf5, encoding part of the lacI family transcriptional regulator; orf6, encoding 1,4-beta-xylanase; △orf6, encoding glycosyl hydrolases family 2, TIM barrel domain which was identical to part of the 1,4-beta-xylanase.
Discussion
Emergence of fosA coharboring with fosA3 in the E. cloacae isolates
The prevalence of fosA3 in E. coli, K. pneumoniae, E. aerogenes, and Salmonella has been frequently reported recently, while it is rarely reported for fosA.18,20–23,27–31 To our surprise, our research data showed that fosA is more prevalent than fosA3 in E. cloacae isolates (42.1% vs. 15.8%). Interestingly, three fosA3-positive isolates were found to be concomitant with fosA genes. Although the fact that the fosfomycin-resistant gene fosA3 usually links with β-lactamase determinants such as blaCTX-M, blaSOF-1, blaNDM-1, and 16S ribosomal RNA methyltransferase determinants armA and rmtB in E. coli, K. pneumoniae, E. aerogenes, and Salmonella from clinical and nonclinical origins in Asia region was frequently reported recently,20–23,28–30,32–37 after checking all the available information, no fosA3-positive isolates coharboring with fosA were reported in clinical isolates so far, therefore, this is the first report of the fosfomycin-modifying enzyme gene fosA coexisted with fosA3 in one E. cloacae strain isolated from pets with UTI.
fosA3 possesses the stronger potential to spread than fosA
The fosA gene previously reported in S. marcescens could be transferred to the recipients E. coli by using liquid mating conjugation method with other resistance markers. 12 However, the hardship of the acquisition of fosA transconjugants in our study suggested that fosA gene from E. cloacae likely locates on the chromosome, which was similar to the fosA2 chromosomally located on E. cloacae from a water sample. 38 Moreover, comparing to the fosA isolates, transconjugants of fosA3 without fosA were obtained successfully. Also, the fosfomycin resistance phenotype combined with ampicillin-resistant determinants and phenotype was easily cotransferred to the recipient E. coli C600. This suggests that fosA3 locates in mobile genetic element (or transposon) and possesses stronger potential to spread than that of fosA.
fosA3 exhibits stronger ability of inactivating the fosfomycin than fosA
Different fosfomycin resistance enzymes displayed variable phosphorylation ability to fosfomycin. FosA inactivates fosfomycin by the addition of glutathione to the oxirane ring with the participation of Mn2+or the K+. 39 FosB inactivates fosfomycin with the performance of Mg2+ and L-cysteine. 13 FosC inactivates fosfomycin through the introduction of a phosphate group in the molecule, with ATP as a donor. 15 FosX in different pathogens displays different inactivation efficiency of fosfomycin. For example, robust resistance to fosfomycin was performed when fosX expressed in Listeria monocytogenes, while no significant resistance to fosfomycin could be observed when fosX expressed in Mesorhizobium loti. 14 The MICs data in our research showed that fosA3 may possess stronger ability of inactivating the fosfomycin than fosA. The MICs of the fosA3 parental isolates and their transconjugants to fosfomycin were all ≥256 mg/L. However, MICs of the fosA-positive isolates were a little different from that of fosA3-positive strains, although all fosA-positive E. cloacae isolates were resistant to fosfomycin according to the EUCAST interpretive criteria, the MICs of them were various. Apart from the MIC of EC5 was ≥256 mg/L, the MICs of the rest four fosA-positive isolates, EC2, EC3, EC4, and EC19, were 64, 128, 64, 128 mg/L, respectively.
Variable genetic mobile elements are responsible for the dissemination of fosA and fosA3
Genetic mobile elements, especially plasmid and transposon, play an important role in the dissemination of fosA. Previous literatures reported that fosA from S. marcescens was located on conjugative plasmid or the transposable element 2921.12,40 Another report of fosA characterized that fosA from Enterobacteriaceae was located on InC M plasmid. 41 The plasmid replicons of InC HI2 and InC L/M were detected in the 5 fosA-positive isolates in our study, but the acquisition of transconjugants of fosA was failed, implying that the fosA gene is supposed to be chromosomal-located in these E. cloacae strains. So, it is difficult to confirm that the plasmid InC HI2 and InC L/M may be the vehicle of fosA gene in the 5 E. cloacae strains, While mobile sequence which was similar to part of transposon 2,921 harbored fosA were detected in fosA-positive E. cloacae isolates, which suggested that mobile element transposon may cause dissemination of fosA in E. cloacae under certain circumstance. Moreover, the high similarity profiles of the four groups (Fig. 3) showed that they had a closer relationship and the vertical transmission might happen between each other, while the phylogenic distance of the above four groups indicated that they came from different sources, which suggests that the horizontal gene transfer might happen among them. Different from fosA, insertion sequence IS26 involved in facilitating the dissemination of fosA3 across different plasmids.20,28 In our study, incompatibility (Inc) FrepB and HI2 were typed and verified in the fosA3-positive isolates and their transconjugants, respectively. Genetic context analysis showed that fosA3 were flanked by mobile element IS26, which was identical to that existed in E. coli plasmid pHK23a and pC0206A.19,28 ERIC-PCR confirmed that EC06 (from cat) and EC18 (from cat) had similar relationship and they may arise from the common ancestor, while the low similarity between group 1(EC06 and EC18) and group 3(EC08) revealed that they may come from different population and we supposed that genetic drift horizontally might take place between them.
Nucleotide sequence accession numbers
The nucleotide sequences of fosA and fosA3 in this study has been deposited in GenBank under the accession number JN232517, KX215983, KX215984, KX215985, KX 215986, and KX215987.
Conclusions
In conclusion, fosA and fosA3 coexist in E. cloacae from companion animal in this study. Comparing to the fosA determinant, fosA3 possessed stronger potential to be spread, and different inactivation efficiency of fosfomycin between fosA and fosA3 was observed after analyzing the data of MICs. Three plasmid replicons and mobile element IS26 were found existed in fosA- and fosA3-positive isolates, which indicated that fosA and fosA3 can be facilitated to transfer across different plasmids. 28 Different genetic backgrounds exist in these fosA- and fosA3-positive isolates, and different mobile elements may confer the dissemination of fosA and fosA3. With the aid of mobile elements, the transmission of resistant determinants from pet pathogens to human pathogens by close contact would be accelerated. 20 So, it is necessary to ascertain the resistance mechanism of fosA, and the interaction between fosA and fosA3 should be explored in the future.
Footnotes
Acknowledgments
We are grateful to Doctor Chia-Lan Wang from National Taiwan University Zoonoses Research Center for technical assistance. We thank Professor Wu from National Chengkung University for providing the recipient strain E. coli C600 (streptomycin resistant).
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from Chinese National Science Foundation (Grant No. 30571374, 30771603, 31072136, 31270171), the Genetically Modified Organisms Technology Major Project of China (2014ZX08006-001B), the Project from Jiangsu Overseas Research &Training Program for University Prominent Young and Middle–aged Teachers and Presidents, the China Postdoctoral Science Foundation (2013M541740), Natural Science Fund of Colleges in Jiangsu Province(17KJD230001), Research Project of Practice Teaching Reform in 2018 of Zhaoqing University (sjjx201810), Young Innovative Talents Projects of Colleges in Guangdong Province in 2018 (Natural Sciences) (2018KQNCX289), Scientific Research Start-up Fund of Zhaoqing University (611/180160), Youth Fund Project of Regional Joint Fund of Basic and Application Basic Research Fund in Guangdong Province (2019A1515111186), and Social Public Welfare Science and Technology Research Project of Zhongshan City (2019B2022).