
Abstract
The emergence of multidrug-resistant (MDR) bacterial infections poses a catastrophic threat to medicine. The development of phage-based therapy combined with antibiotics might be an advantageous weapon in the arms race between human and MDR bacteria. A cocktail composed of the MDR Acinetobacter baumannii infecting bacteriophages with high lytic activity was used in combination with antibiotics to destroy a bacterial biofilm in human urine. A. baumannii exhibited varying susceptibility to the host range of bacteriophages used in this study, ranging from 56% to 84%. This study demonstrated that bacteriophages could reduce biofilm biomass in a human urine model, and some of the antibiotics commonly used in the treatment of urinary tract infection (UTI) act synergistically with phage cocktails. Additionally, the combined treatment showed a significantly greater reduction of biofilm biomass and clearance of persister cells.
Introduction
A
UTI is one of the major hospital problems and constitutes 40% of all nosocomial infections, usually associated with catheterization and percutaneous nephrostomy tubes.5,6 The most common pathogenic agents of this infection are the bacteria from the Enterobacteriaceae family (∼70% of cases). 7 A. baumannii is rarely isolated during a UTI episode (<2%), and mostly from debilitated elderly patients or from patients with prolonged indwelling catheter-related infections. 8 Nevertheless, the epidemic potential of MDR A. baumannii makes this bacterium of special interest despite its relatively low input into hospital-acquired UTI; this is more urgent in relation to clinically critical populations prone to be affected with this pathogen, as above. MDR A. baumannii might also be isolated from animal urine (e.g., cats or dogs). 9
In the previous century, Acinetobacter infections were treatable with ampicillin, carbenicillin, gentamicin, or nalidixic, however, those antibiotics are now ineffective. Nowadays, better treatment options, including carbapenems (meropenem, imipenem), aminoglycosides (amikacin, levofloxacin), broad-spectrum cephalosporins (ceftazidime, cefotaxime, ceftriaxone, cefepime), and combinations of beta-lactamase inhibitors with an antibiotic (ampicillin/sulbactam), are used to treat A. baumannii infection. 8 Colistin or tigecycline is recommended to treat XDR A. baumannii infection. Therefore, minocycline may retain antimicrobial activity if the strain is resistant to other tetracyclines. 10
Most frequently, there is a need for the use of antibiotic combinations against XDR A. baumannii infection, for example, doripenem with colistin, sulbactam, amikacin, or tigecycline.11,12 Due to the high nephrotoxicity of colistin, a treatment approach often based on combination with another antibiotic (rifampicin, tigecycline, carbapenems) is used to decrease colistin dosage. The next combination is sulbactam and cefepime, meropenem, imipenem, or amikacin.4,10 The effect of combined therapies is diverse, sometimes equal to monotherapy, other times superior or inferior.
Due to an increasing number of isolated antibiotic-resistant A. baumannii strains, antibiotic therapeutic alternatives are being investigated. Mostly mentioned are bacteriophages (phages for short) able to lyse bacterial host cells. In 2010 two phages against A. baumannii, AB1 and AB2, were isolated, and since then interest in them has increased. Phages are very active natural enemies of bacteria. Unfortunately, their action is based on receptor specificity, and one phage may not kill all strains of specific bacterial species. Single phage therapy may be effective in the treatment of A. baumannii infection, as demonstrated by Jeon et al. 13 However, in most situations, the use of a phage cocktail is necessary.14–16
There is also a need to isolate new environmental phages to expand the repertoire of treatments to use against A. baumannii. Regeimbal et al. 17 showed that the five-phage-based cocktail of wild phages infecting A. baumannii demonstrated therapeutic efficacy in a mouse model, arresting infection spread and necrosis. Bacteriophages may also be a source of endolysins–enzymes that degrade the bacterial cell walls. Treatment with lysin, isolated by Lood et al.. 3 was able to reduce planktonic forms and biofilm of A. baumannii, in both in vitro and in vivo conditions.
Alternatively, to enhance the activity, bacteriophages can be combined with antibiotics. The synergy between these factors is called phage antibiotic synergy (PAS). Such a combination demonstrates strong control of A. baumannii and reduction in the development of resistant strains, for example, when the phage interacts with the bacterial drug efflux systems as a receptor. 18
Different modes of action of phages and antibiotics promote the possibility of combining them. Positive interactions between antimicrobial agents depend on treatment conditions, for example, dosage, frequency, time points, and order of administration. 15 Therapies based on lytic phages combined with carbapenem antibiotics (meropenem, imipenem), colistin, or ciprofloxacin, enhanced the antimicrobial activity of both agents, and resistant A. baumannii strain became sensitive to applied factors.15,18 Examination of bacteriophage lysins with an antibiotic, such as levofloxacin, ceftazidime, colistin, or polymyxin B, also demonstrated a positive effect in the treatment of A. baumannii infection.14,19
Therefore, we aimed to analyze the activity of the different antibiotics (ciprofloxacin [CIP], levofloxacin [LVX], trimethoprim/sulfamethoxazole [SXT], gentamicin [GN], amikacin [AMK]; tobramycin [TOB]; colistin [CS], imipenem [IMP]; and meropenem [MRP]) combined with an environmental bacteriophage cocktail against A. baumannii biofilm under human urine.
Materials and Methods
Bacterial strains and growth conditions
In total, 25 MDR A. baumannii strains isolated from urinary tract infections at the Pomeranian Medical University in Szczecin, Poland, were used in the study. Strains were identified with the use of the VITEK®2 Compact Bacterial Identification System (bioMérieux, Inc., Durham, NC) and confirmed by MALDI-TOF mass spectrometry. Antibiotic susceptibility testing and assessment of the minimal inhibitory concentration was performed according to CLSI guidance. 20 Bacterial strains were stored at −80°C before use. Bacterial strains were grown on blood agar (bioMérieux, Inc.) for 18–24 hr at 37°C to revitalize.
Bacteriophage isolation and initial characterization
Bacteriophages were isolated from different environmental samples collected in the West Pomeranian Region, Poland. Bacteriophages were isolated according to Guzmán et al. 21 with the use of different strains of A. baumannii. Bacteriophages were initially characterized to determine their lytic development, adsorption to the host cells, stability, host range, and visualize their lytic activity in bacterial liquid culture.
One-step growth of isolated phages infecting A. baumannii cells was prepared according to Turner et al. 22 Briefly, bacteriophages at a multiplicity of infection (MOI) of 0.05 were added to the bacterial culture and allowed to adsorb for 5 min. Afterward, the sample was centrifuged at 13,000 g, the supernatant removed, and the pellet resuspended in equal to the initial volume of Luria-Bertani (LB) broth. The sample was diluted to 10−7, incubated, and titrated by triplicate overlay plaque assay to determine the single burst.
The host range was determined using the efficiency of plating (EOP) methodology, according to Mirzaei and Nilsson, 23 to avoid false-positive results. Briefly, the decimal dilutions of the phage lysates were used. For EOP testing, overlay plaque assay plates were prepared to contain 10 μL of bacteriophage dilution and 190 μL of bacterial cultures in an exponential growth phase. Following overnight incubation at 37°C, the relative EOP was expressed as the ratio between the titer in plaque-forming unit (PFU) × mL−1 for a given isolate and the titer for the propagating host from overlay plaque assays.
Bacteriophage lysis of bacterial liquid culture was prepared, according to Łubowska et al. 24 Briefly, overnight bacterial cultures were diluted 1:200 in fresh LB broth and incubated with shaking (180 rpm) at 37°C. Bacterial culture was incubated to reach the early logarithmic phase (OD600 = 0.2). Afterward, bacteriophages solution was added to reach MOIs equal to 0.1, 0.25, and 0.5. The optical density of culture was measured at 30-min intervals. Additionally, bacteriophage thermal, pH, and UV stability were performed according to Szymczak et al. 25
Evaluation of biofilm production level and phage biofilm-reducing activity
The biofilm biomass quantification was prepared with the use of the Crystal Violet staining method detailed by Chajęcka-Wierzchowska et al. 26 with minor modifications. The standardized overnight culture of A. baumannii (density of 109 cells × mL−1) was added to 180 μL of the trypticase soy broth (TSB) broth and incubated for 24 hr at 37°C. Afterward, bacterial culture was removed and the established biofilm was washed five times with phosphate-buffered saline (PBS) and fixed with ethanol for 15 min. Plates were dried at 37°C, filled with Crystal Violet solution (1%; Merck), and incubated for 5 min. After incubation, Crystal Violet was removed, and plates were washed with running water and dried. A mixture of glacial acetic acid (33%; Merck) and ethanol (2:8) was used to resolubilize Crystal Violet. The absorbance was measured at 595 nm with the use of the En-Vision multimode plate reader (PerkinElmer).
The classification proposed by Stepanović et al. 27 was used to determine the biofilm formation ability of A. baumannii strains, respectively: no biofilm producer (OD ≤ ODc), weak biofilm producer (ODc < OD ≤2 × ODc), moderate biofilm producer (2 × ODc < OD ≤4 × ODc), and strong biofilm producer (4 × ODc < OD), where ODc is the optical density of the blank sample, and OD is the optical density of the tested sample measured at 595 nm.
To establish bacteriophage biofilm-reducing activity, 24-hour-old biofilm was washed five times with PBS and treated with phage suspensions (final concentration: 1 × 107 PFU × mL−1) in TSB medium and incubated for 6 hr at 37°C. Afterward, the biofilm biomass was measured as described above. A slight modification was implemented to analyze bacteriophage biofilm-reducing activity in human urine. The matured biofilm was washed five times with PBS, followed by the addition of human urine (in equal volume to that of TSB) filtered through 0.22 μm syringe filters, then incubated for 1 hr before bacteriophage addition (for more specific information related to urine composition see Supplementary Table S1).
Phage cocktail preparation
For cocktail preparation, bacteriophages were mixed in equal proportions to obtain a final concentration of 1 × 109 PFU × mL−1. The cocktail contained Aba-1, Aba-2, Aba-3, Aba-4, and Aba-6. Bacteriophage Aba-5 was excluded from further analysis due to its low activity. The bacteriophage cocktail antibiofilm and lysis activity were tested against A. baumannii strains as described above.
Assessment of phage cocktail activity in combination with antibiotics
The A. baumannii AB20 [minimal inhibitory concentration (MIC) (mg × L−1): CIP = 8; LVX = 16; SXT = 64; AMK = 32; TOB = 2; GN = 64; CS = 1; IMP = 16; MRP = 32] strain was used as a representative to demonstrate these experiments. The 24-hr biofilm was washed with PBS and submerged in 180 μL of filtered urine. The antibiotics were added at ¼, and ½ of the MIC and the five bacteriophage-based cocktail was added in final phage concentration approx. 1 × 107 PFU × mL−1 and incubated at 37°C for 6 hr. Biofilm biomass was analyzed with the Crystal Violet staining method as described above.
Effect of synergistic action on persister cell regrowth
Following 6 hr of combined treatment, biofilms in urine were rinsed five times with PBS, and wells were refilled with TSB medium and incubated for 24 hr at 37°C. After incubation, and PBS washing, the cell viability was measured using the resazurin assay according to Skogman et al. 28 with minor modifications. Following PBS rinsing, the wells were filled with 200 μL of fresh TSB medium containing 20 mM of resazurin and incubated for 60 min at 37°C with shaking (200 rpm). After incubation, the medium was transferred to the black 96-well plate, and fluorescence (λex = 520 nm; λem = 590 nm) was measured with the use of the En-Vision 2105 multimode plate reader (PerkinElmer). When statistically significant reduction (p < 0.05) of biofilm occurred in the combined treatment group compared with phage treated, results were marked as synergistic.
Statistical analysis
All experiments were conducted with a minimum of five biological replicates. Technical replicates were averaged to produce replicate means that were used for analysis. Mean values were compared with the use of one-way analysis of variance followed by Tukey's post hoc test for individual comparisons. Differences were considered statistically significant if p < 0.05 (GraphPad Prism 8.3.0).
Results
The A. baumannii strains used in this study were selected due to the source of isolation (UTI) and their multidrug resistance. All strains used in this study were resistant to ciprofloxacin (CIP), levofloxacin (LVX), trimethoprim/sulfamethoxazole (SXT), gentamicin (GN), and netilmicin (NET). Additionally, 8% (2 of 25) were resistant to amikacin (AMK); 28% (7 of 25) to tobramycin (TOB); 36% (9 of 25) were resistant to colistin (CS) and imipenem; and 40% of selected strains were resistant to meropenem (MRP). Only 3 of 25 (12%) A. baumannii strains were weak biofilm producers, while 28% of isolated strains produced biofilm moderately, and the major part (60%) produced high biofilm biomass (Table 1).
Antibiotic Resistance and Biofilm Formation of Acinetobacter baumannii Strains Isolated from Urinary Tract Infections Used in This Study and Bacteriophage Host Range
The propagation host.
AMK, amikacin; CIP, ciprofloxacin; CS, colistin; EOP, efficiency of plating; GN, gentamicin; IMP, imipenem; LVX, levofloxacin; MRP, meropenem; NET, netilmicin; SXT, trimethoprim/sulfamethoxazole; TOB, tobramycin.
The host range of bacteriophages indicates the susceptibility of A. baumannii strains to bacteriophage infection varying from 56% to 84%. The broadest host range was shown by Aba-3 phage (84% of susceptible strains), and the narrowest host range is 56% of susceptible strains achieved by Aba-5 (Table 1). The host range analysis also included the use of other Gram-negative bacterial strains of Acinetobacter spp., Klebsiella spp., Escherichia coli, Enterobacter spp., Morganella spp., and Pseudomonas spp. Apart from A. baumannii, bacteriophage plaque formation was not observed on other bacterial species (except Aba-4 that produce plaques on Acinetobacter lwoffii, EOP = 0.2). All of the bacteriophages used in this study were isolated between 2017 and 2019 from different isolation samples with the use of standard enrichment technique.29,30
All isolated phages formed clear, 1–4 mm plaques surrounded by a halo zone. The one-step growth was performed with the use of propagation strain. The latent period of isolated bacteriophages varied from 21 to 51 min, and there were differences in the number of progeny phage produced in one cycle of bacteriophage replication that varied from 46 (Aba-2) to 226 PFU (Aba-3) per infected cell. (for more information see Supplementary Tables S2 and S3 for bacteriophage stability data).
Bacteriophages, Aba-1 (Fig. 1A), Aba-2 (Fig. 1B), Aba-3 (Fig. 1C), Aba-4 (Fig. 1D), and Aba-6 (Fig. 1F), exhibit high lytic activity against A. baumannii. The complete reduction of bacterial cultures was observed at all used MOIs. The phage Aba-5 does not completely lyse bacterial culture, and fast regrowth of the optical density of bacterial culture was observed (Fig. 1E)
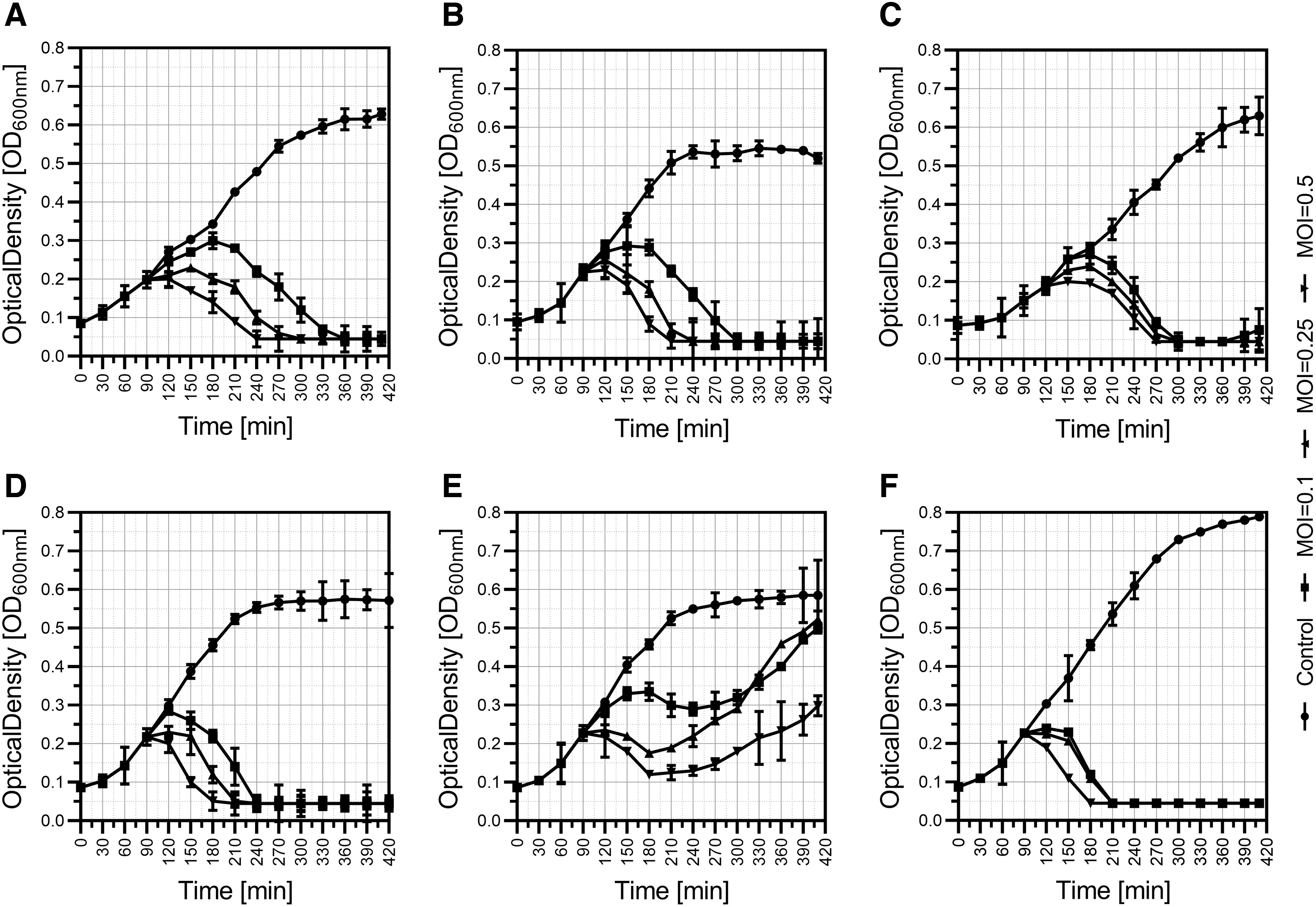
Bacteriophage-mediated lysis of Acinetobacter baumannii liquid culture. The experiment was conducted with the use of LB medium and incubated with shaking (180 rpm) at 37°C. The phages were added at the early logarithmic phase of growth (OD600 ∼0.2) at different MOI equal to 0.1, 0.25, and 0.5, and the OD was measured every 30 min.
To assess the activity of bacteriophages in human urine, we replaced the growth medium with human filtered urine. Exchanging the medium for urine does not result in changes in biofilm biomass (6 hr incubation, data not shown). The usage of all bacteriophages significantly decreased the A. baumannii biofilm biomass. The antibiofilm activity observed in the urine conditions was either significantly lower or did not differ when compared with antibiofilm bacteriophage assay in the TSB medium (Fig. 2). Bacteriophage Aba-1 posed the highest antibiofilm activity and reduced bacterial biofilm biomass by 77.69% and 67.45% in TSB medium and urine, respectively. The lowest antibiofilm properties were exhibited by Aba-5, decreasing biofilm biomass by ∼35%, regardless of the liquid used. Other phages: Aba-2 reduces 55% and 34%, Aba-3 (53%, 44%), Aba-4 (61% and 52%), and Aba-6 (62% and 91%) of biofilm biomass in TSB and urine, respectively (Fig. 2).
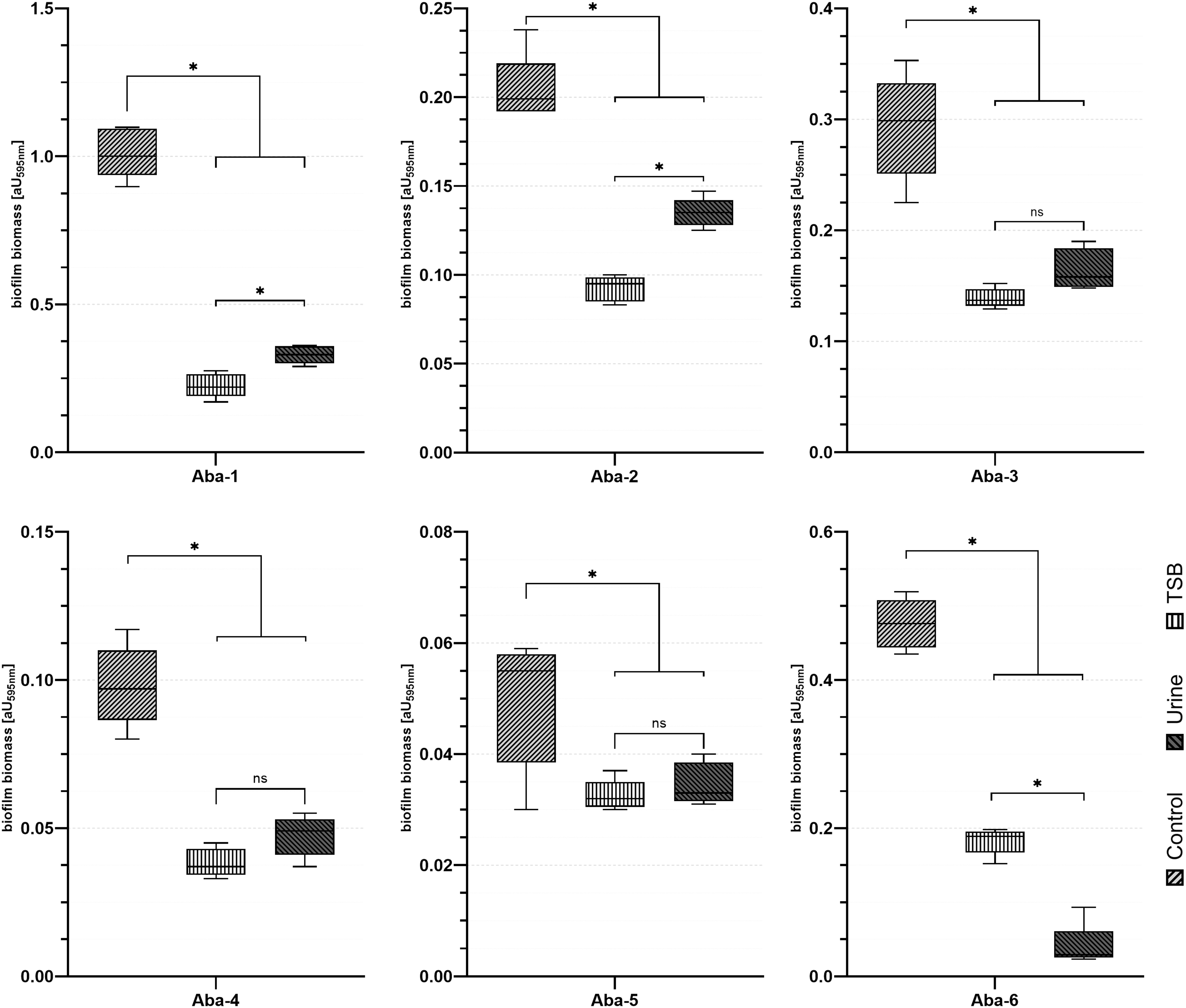
Influence of bacteriophages addition on 24-hr cultured biofilm of the A. baumannii strains in culture medium and urine after 6 hr of incubation. Bacteriophage treatment significantly reduce A. baumannii biofilm under TSB broth and urine conditions. To assess phage activity, medium was removed and wells were refiled with TSB medium or urine. Bacteriophages in final concentration: 1 × 107 PFU × mL−1 were added and incubated for 6 hr. Following incubation, biofilm was stained with the use of Crystal Violet staining method. Each experiment was performed in triplicate and results are presented as the mean ± maximum/minimum. Asterisks indicate results significant at the level of 0.05. TSB, trypticase soy broth.
We analyzed the activity of the bacteriophage cocktail (containing Aba-1, Aba-2, Aba-3, Aba-4, and Aba-6 in equal proportions) combined with antibiotics against formed biofilm biomass in the human urine model. Bacteriophage Aba-5 was excluded from further analysis due to its low activity. The AB20 strain was selected as representative for these experiments due to susceptibility to all phages used in phages cocktail, high antimicrobial resistance, and strong biofilm production (for phage/antibiotic combination activity regarding all main host please see Supplementary Table S4).
The use of bacteriophage cocktail in combination with CIP, SXT, GN, TOB, IMP, and MRP resulted in a significant reduction of biofilm biomass (when compared with only phage-treated control) in a dose-dependent manner. The highest reduction of biofilm biomass was observed when the phage cocktail was combined with SXT and achieved 94.3% and 98.6% when ¼ and ½ of MIC values were implemented, respectively (Fig. 3C). Results are significant (p < 0.05) if compared with phage cocktail treatment resulting in 77% of biofilm biomass reduction. The use of CIP combined with the phage cocktail resulted in 87 and 93.3% reduction for ¼ and ½ of MIC, respectively (Fig. 3A). The use of GN, TOB, IMP, and MRP similarly resulted in a significantly higher reduction of A. baumannii biofilm biomass (Fig. 3E, F, H, J).
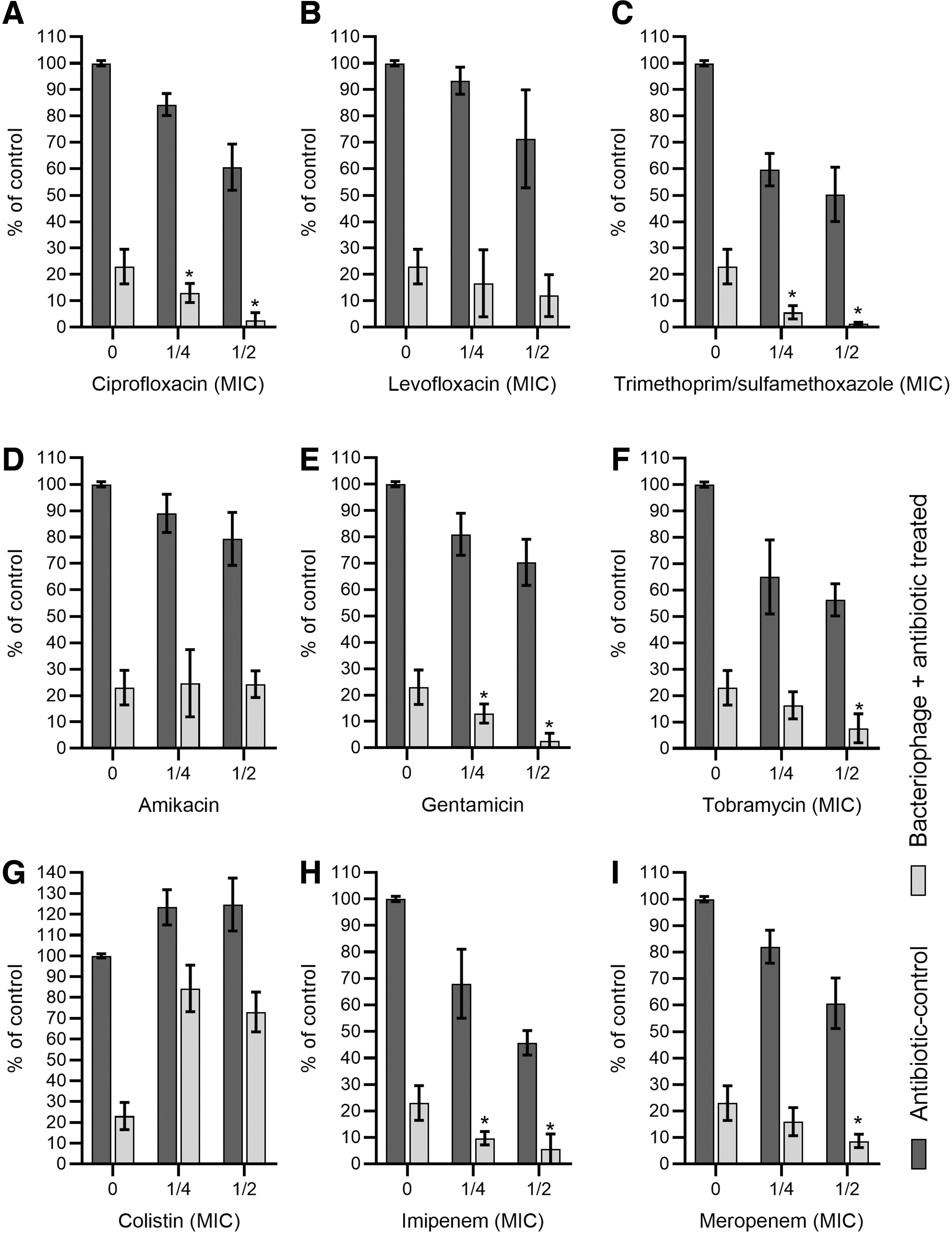
Effect of biofilm disruption in a urine environment with the use of a combination of bacteriophage cocktail and antibiotics. Twenty-four-hour biofilm was washed with PBS buffer and submerged with filtered urine. The antibiotics [ciprofloxacin
The use of PC (phage cocktail) combination with LVX and AMK did not act synergistically. Results indicate no interaction between these components. The lowest outcome was observed for the combination of CS with PC. The use of the sub-MIC of CS exclusively increased the biofilm production abilities of the tested A. baumannii strain. The biofilm biomass increased (p < 0.05) to ∼120% in both concentrations used when compared with the control (p < 0.05). Furthermore, the antagonism between CS and PC was observed with regard to the bacteriophage's antibiofilm properties (Fig. 3G).
The synergistic effect on biofilm destruction, in most cases, also resulted in the limitation of persister cell regrowth. The most significant effect was observed for SXT combined with a PC. The SXT-PC combination limited bacterial regrowth to 18% and 6% for ¼ and ½ of MIC values, respectively. A similar, positive effect was also observed for TOB, IMP, and MRP in all used concentrations, whereas the combinations with CIP and LVX were efficient only in ½ of the MIC value. No significant effect was observed for gentamicin at any concentration (Fig. 4).

Effect of combined treatment on 24 hr A. baumannii biofilm persister cells regrowth. Summarized results show the ratio of the resazurin-based biofilm metabolic activity compared with control. After 6 hr of the combined treatment, biofilm was rinsed five times with PBS, and wells were refilled with filtered human urine. Following 24 hr incubation, metabolic activity with the use of a resazurin assay was performed. Each experiment was performed in triplicate, and results are presented as the mean ± standard deviation. Asterisks indicate results significant at the level of 0.05.
Discussion
Ever-increasing A. baumannii antibiotic resistance puts these bacteria in first place on the WHO global priority list for research and development of new antibiotics and treatment options. 31 The development of new treatment options for MDR bacterial infection has become a major challenge for medicine. 32 Bacteriophages seem to be a promising tool to combat MDR strains. Successful treatment of the A. baumannii infection with the use of the bacteriophages was presented in literature.33–35
The bacteriophage cocktail is the best choice of phage application. Cocktail preparation enlarges the host range, which is important in the case of narrow host range infecting phages. 36 Bacteriophages selected for this study represented a relatively wide host range and were efficient in lysis of the A. baumannii cells. Additionally, all of the phages produced a halo zone, which can potentially be correlated with enzymatic biofilm exopolysaccharide matrix degradation.37,38
Synergistic action between antibiotics and bacteriophages might be extremely beneficial for infection treatment outcomes. Unfortunately, the actual knowledge of this phenomenon is limited and mostly speculative. 39 In this study, we tested carbapenems—modified β-lactam antibiotics (imipenem and meropenem), fluoroquinolones (ciprofloxacin, and levofloxacin), aminoglycoside antibiotics (gentamicin, amikacin, tobramycin), and sulfonamide with dihydrofolate reductase inhibitor (sulfamethoxazole/trimethoprim).
The β-lactams and quinolones stimulate virulent phage growth primarily as a consequence of cellular filamentation. 40 The synergistic effect can be explained by the antibiotics mode of action. In this manner, the subinhibitory concentration activates the stress response pathways. Antibiotic-related changes in the bacterial cell morphology enabled rapid phage maturation and accelerated cell lysis.40–42 More giant and upregulated cells permit faster phage assembly, possibly by much faster phage assembly, probably due to more extensive availability of necessary substances and improvement of processes related to the rate-limiting speed of assembly. The cell filamentation was observed for β-lactams and fluoroquinolones.40–42
Kirby 43 showed that aminoglycoside antibiotics (gentamicin) in a continuous culture of Staphylococcus aureus act synergistically with phages. Gentamicin exposure induces the aggregate formation of S. aureus, and these aggregators were more susceptible than the parental cells to the bacteriophage infection. The use of CIP, MRP, and minocycline significantly reduces mortality of Galleria mellonella larvae infected with Burkholderia cenocepacia due to stimulation of the phage lytic activity. 41
In this study, we did not observe synergy between phage and amikacin, but bacteriophage PEV2 infecting exhibit synergy with CIP, AMK, and CS when P. aeruginosa strain FADD1-PA001 was used. Change of the strain to IJP865 resulted in synergy with CIP, AMK, and tobramycin (TOB). However, in the case of strain 20844n, no synergistic action was observed for AMK, CIP, TOB, CS, and aztreonam combinations.
Conversely, the synergistic action of antibiotics, combined with phage on 86.6% of P. aeruginosa strains analyzed, was previously observed by Pletzer et al. 43 This result suggests that the PAS effect can also be strain specific, which confirms results presented in Supplementary Table S4. Additionally, Pletzer et al. 44 observed that the combination of antibiotics with host defense peptides (HDP) increases the efficacy of treatment. They suggest that HDP enhance antibiotic penetration and potential disruption of the stringent stress response. The set of phage lytic enzymes, especially those responsible for local peptidoglycan degradation, could enhance antibiotic penetration through cell wall permeabilization. 45
The synergy between the phage cocktail and minocycline was observed when this combination was successfully used to treat A. baumannii infection. The authors suggest that phage predation imposed selective pressure on infectious agents. This phage pressure favored regrowth of bacteria with limited capsule production and lower virulence.33,39
The use of phage potential to disperse and penetrate bacterial biofilm combined with the antibiotic treatment was presented by Chaudhry et al. 4 2 Combined treatment was particularly effective in P. aeruginosa biofilm removal from layers of epithelial cells. Bedi et al. 46 research showed that phage-mediated biofilm disruption increases antibiotic penetration and activity against Klebsiella pneumoniae. The study suggests that high concentrations of antibiotics is not recommended to investigate bacteriophage/antibiotic synergy. The high concentration of antibiotics could kill bacteria in a surface layer of biofilm, which makes it impossible for bacteriophage penetration and biofilm dispersion.39,42,47
It is worth mentioning that during the therapy, the antibiotic concentration in tissues is lower than the MIC of drug used. 48 Furthermore, patient exposure to a high dose of antibiotics (to reach MIC concentration), may result in adverse events, for example, due to nephro- and neurotoxicity of polymyxins. 49
Among the tested antibiotics, sulfamethoxazole/trimethoprim showed the most significant activity when combined with a phage cocktail. Interestingly, sulfamethoxazole, when combined with phages, does not lead to notable synergy, but in some Gram-negative bacterial increase of plaque diameter and bacteriophage, production was observed without observation of morphological changes of the bacterial cell. Otherwise, trimethoprim demonstrates the PAS effect exhibited by the increase of plaque diameter and bacteriophage production with observed filamentation of Gram-negative bacteria. 50
Polymyxins are currently used as a last-resort antibiotic for Gram-negative bacterial infections. The mode of action of these membrane-active peptides is based on the disorganization of bacterial outer membrane. 51 Biofilm reduction activity of colistin and phage cocktail combination have no synergistic effect. This phenomenon could be explained by the fact that colistin destabilizes the bacterial cell membrane. Danis-Wlodarczyk et al. 52 pointed out that colistin can limit bacteriophage propagation by destabilizing the cell membrane. Apart from that, Jansen et al. observed an increase of the antibacterial effect of the vB_AbaM_KARL-1 bacteriophage at very low MOI (10−7) in combination with colistin in liquid culture. 53
Additionally, we observed a similar effect to Sato et al., who described the increase of the A. baumannii biofilm formation when exposed to a sub-MIC concentration of colistin. The increase of biofilm formation was correlated with overexpression of the AdeFGH efflux pump and autoinducer synthase AbaI. 54 He et al. pointed out that these genes' overexpression accelerate the synthesis and transport of acylated homoserine lactones. 55
The reduction of the biofilm regrowth from persister cells in case of the use of combined treatment could be connected with the enhancement of bacteriophage activity, enzyme-mediated permeabilization of membranes, and digestion of the biofilm matrix. These elements can expose bacteria released from the biofilm matrix to antibiotics and increase antibiotic diffusion through the biofilm matrix. Saggu et al. 56 indicated that enzyme-mediated degradation of biofilm increases the concentration of antibiotics inside the biofilm. Regarding this, the degradation of the biofilm component allows antibiotics to penetrate the biofilm. Therefore, bacteriophages represent unique properties to degrade biofilm and make bacterial cells inside biofilm more susceptible to antibiotics. 57 Additionally, bacteriophages can target persister cells induced by antibiotic treatment. 58
Worrying increases in antimicrobial resistance among UTI substantially increases treatment costs. Even though bacteriophage therapy effectiveness was demonstrated before,59,60 to the best of our knowledge, there is a lack of ex vivo studies showing phage activity in human urine. Our study demonstrated that bacteriophages could reduce biofilm biomass under urine conditions, and some of the antibiotics commonly used in the treatment of UTI act synergistically with used phage cocktail. These results show that combined treatment of the UTI may result in greater efficiency than using them separately. This seems to be especially important for the treatment of catheter-associated urinary tract infection; foreign body in the urinary tract predisposes to microbial colonization and biofilm formation.
Hard-to-eradicate catheter biofilm 61 may be limited with the use of the phage/antibiotic combined treatment. Although the role of bacteriophages in shaping urobiome composition remains unclear, further studies are required to understand the role of lytic phages combined with antibiotics in UTI treatment outcomes. 62
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.