
Abstract
Diarrheagenic Escherichia coli cause diarrheal diseases, which are a public health concern and affect mainly developing countries. Multidrug-resistant (MDR) pathogens have been spreading in different sources, including animals and the environment. E. coli strains were obtained from a small-scale pig farm and 33 antimicrobials were tested. All strains were classified as MDR and harbored several antimicrobial resistance genes (ARGs) [blaCMY, blaOXA-1-like, blaSHV, tet(A), tet(B), aadA, aac(6’)-Ib, aph(3′)-Ia, sul1, sul2, sul3, floR, and cmlA] and plasmids. Besides, mutations in quinolone resistance-determining region of GyrA (Ser83Leu and Asp87Asn) and ParC (Glu84Asp) were detected. Among the MDR E. coli, nine strains (52%) presented diarrheagenic virulence genes, including genes related to Shiga toxin-producing E. coli (STEC), enteroinvasive E. coli (EIEC), and enteroaggregative E. coli (EAEC). The pulsed-field gel electrophoresis results showed a high genetic diversity among the MDR E. coli strains. Multilocus sequence typing (MLST) analyses revealed different sequence types phylogenetically related to each other, including ST10 and ST56. Subtyping of MLST by fimH gene showed different fimH type. This study shows a high genetic diversity among MDR ARG-producing E. coli belonging to STEC, EIEC, and EAEC pathotypes obtained from a small-scale pig farm and contributes to the monitoring of antimicrobial-resistant pathogens worldwide, mainly in environmental samples, which are associated with One Health framework.
Introduction
Diarrheagenic Escherichia coli (DEC) cause diarrheal diseases in humans and animals. According to the virulence mechanisms, DEC are classified into different well-defined pathotypes, such as Shiga toxin-producing E. coli (EHEC/STEC), enteroinvasive E. coli (EIEC), enterotoxigenic E. coli (ETEC), enteroaggregative E. coli (EAEC), diffusely adherent E. coli (DAEC), and enteropathogenic E. coli (EPEC). 1 Diarrheal diseases are considered a public health concern and mainly affect the developing countries, where high rates of morbidity and mortality occur in young children. Diarrheal diseases are responsible for high health care costs in industrialized countries. 2
Poor living conditions and food chain are closed related to the acquisition of DEC and, consequently, diseases caused by them. 3 Multidrug-resistant (MDR) pathogens are being detected in several sources, including animals and the environment, and consequently, the One Health concept is being increasingly applied. Animals and the environment have been reported as reservoirs and disseminators of MDR pathogens, principally E. coli, which carry different antimicrobial resistance genes (ARGs), including β-lactamase-encoding genes. Besides, MDR E. coli co-producing ARGs and mutations in quinolone-resistance determining region (QRDR) have been described.4,5
MDR DEC belonging to different pathotypes, including STEC, EIEC, and EAEC, have been reported worldwide, which is striking and worrying.6–8 Therefore, this study aimed to characterize molecular MDR E. coli strains obtained from a small-scale pig farm regarding ARGs, diarrheagenic virulence genes, plasmids, and genetic relatedness, as well as phylogenetic groups, sequences types, and fimH type.
Materials and Methods
Samples and bacterial isolation
A total of 22 samples, including feces (n = 11) and soil (n = 11), were collected in a Brazilian pig farm located in Sales Oliveira City, São Paulo State, Brazil. On this farm, there are no reports of antimicrobial use. The tops of fresh feces were collected using sterile pots and spatulas on the floor where the pigs were kept. The soil samples were collected up to 10 m around the site where the pigs were kept at a depth of up to 15 cm after the surface layer was removed. For these reasons, there was no contact with the animals. The samples were mixed and one gram of each sample was added in the Brain Heart Infusion (BHI) broth (Oxoid) and incubated at 37°C for 18–24 hours. Then, the samples were seeded on MacConkey Agar (Oxoid) and incubated under the same conditions described above. One lactose fermenter colony from each sample was selected and stocked in BHI broth plus 15% glycerol for further experiments. 6
PCR conditions and sequencing
All PCRs were performed using the reaction mixture (50 μL) containing 2 μL (1.25 U) of JumpStart™ Taq DNA Polymerase (Sigma-Aldrich), 5 μL of 10 × PCR buffer, 3.5 μL (25 mM) of MgCl2, 1 μL (10 mM) of deoxynucleotide solution mix, 2 μL (25 μM) each primer, 29.5 μL of purified water, and 5 μL (100 ng) of DNA on the ProFlex™ PCR Thermocycler System (Applied Biosystems, Singapore). The BigDye Terminator Cycle Sequencing Kit (Applied Biosystems) was used for sequencing of the amplicons on the ABI 3500xL Genetic Analyzer (Applied Biosystems). The sequences were analyzed using the ChromasPro v.1.7.6 software (Technelysium Pty. Ltd.) and compared to those available in GenBank using the MegaBlast algorithm.
Molecular identification
Genomic DNA was extracted using the GenElute™ Bacterial Genomic DNA Kit (Sigma-Aldrich) according to the manufacturer's recommendations. After that, the genomic DNA was used as template for sequencing of the 16S rDNA using the primers fD1 (5′-AGAGTTTGATCCTGGCTCAG-3′) and rP2 (5′-ACGGCTACCTTGTTACGACTT-3′). 9
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was performed using the disk diffusion method according to the Clinical and Laboratory Standards Institute (CLSI; M100, 28th ed.). A total of 33 agents from seven categories was tested, including β-lactams (ampicillin, ampicillin-sulbactam, piperacillin-tazobactam, amoxicillin-clavulanate, cefazolin, cefaclor, cefuroxime, cefotaxime, ceftriaxone, ceftazidime, cefepime, aztreonam, meropenem, imipenem, and ertapenem), aminoglycosides (streptomycin, tobramycin, gentamicin, and amikacin), quinolones (nalidixic acid, ciprofloxacin, lomefloxacin, levofloxacin, norfloxacin, and ofloxacin), tetracyclines (doxycycline, tetracycline, and minocycline), folate pathway antagonists (sulfonamides, trimethoprim, and trimethoprim-sulfamethoxazole), phenicols (chloramphenicol), and nitrofurans (nitrofurantoin). E. coli strains were classified as MDR according to the definitions established by Magiorakos et al. 10
Detection of ARGs, mutations in QRDR and plasmids
The ARGs that confer resistance to β-lactams (blaSHV, blaOXA-1-like, blaCMY, blaVEB, blaPER, blaCTX-M-Gp1, blaCTX-M-Gp2, blaCTX-M-Gp8, and blaCTX-M-Gp9), aminoglycosides [aadA, aac(3)-Ia, aac(6’)-Ib, aac(6′)-Ih, aac(3)-IIa, ant(2”)-Ia, aph(3’)-Ia, and aph(3′)-VI], quinolones (oqxA, qepA, oqxB, qnrA, qnrB, and qnrS), tetracyclines [tet(A) to tet(D), tet(G), and tet(M)], sulfonamides (sul1, sul2, and sul3), and phenicols (floR and cmlA) were researched by PCR.11–20 Detection of mutations in QRDR was performed using the primers and conditions previously described by Jia et al. 21 (Supplementary Table 1). The alignment of gyrA and parC genes was performed against the E. coli str. K-12 substr. MG1655 (GenBank accession number U00096) using the Clustal Omega EMBL-EBI multiple sequence alignment. Twenty plasmid families (IncHI2, IncHI1, IncI1, IncF, IncFIB, IncFIA, IncFIC, IncFIIS, IncL/M, IncW, IncP, IncA/C, IncK, IncN, IncY, IncT, IncX, IncU, IncR, and ColE like) were detected by PCR-based replicon typing22,23(Supplementary Table 2).
Antimicrobial Resistance Profile, Antimicrobial Resistance Genes, and Plasmids in Multidrug-Resistant Escherichia coli
Feces, A; Soil, S.
AMP, ampicillin; CFZ, cefazolin; CRX, cefuroxime; STP, streptomycin; SXT, trimethoprim-sulfamethoxazole; SUL, sulfonamides; TRI, trimethoprim; TET, tetracycline; DOX, doxycycline; GEN, gentamicin; TOB, tobramycin; MIN, minocycline; CFC, cefaclor; AMI, amikacin; CLO, chloramphenicol; APS, ampicillin-sulbactam; NIT, nitrofurantoin; CRO, cefotaxime; CTX, ceftriaxone; AMC, amoxicillin-clavulanate; PIT, piperacillin-tazobactam; CAZ, ceftazidime; NAL, nalidixic acid; CIP, ciprofloxacin; LVX, levofloxacin; NOR, norfloxacin; LMX, lomefloxacin; OFX, ofloxacin.
Incompatibility group, Inc.
Isolates that presented mutations in QRDR.
ARG, antimicrobial resistance gene; QRDR, quinolone resistance-determining region.
Detection of diarrheagenic virulence genes
Diarrheagenic virulence genes (aatA, ehxA, stx1, stx2, eaeA, bfpA, elt, est, ipaH, aaiC, AA probe, aggR, and aap) were screened by PCR following the conditions previously described24–28 (Supplementary Table 3).
Determination of phylogenetic groups
Five target genes (yjaA, chuA, arpA, TspE4.C2, and trpA) were used for determining the phylogenetic groups [A, B (B1 and B2) C, D, E, and F] according to Clermont et al. 29 (Supplementary Table 4).
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) was performed to determine the genetic relatedness of the MDR E. coli strains. Genomic DNA was digested using 50 U of the restriction enzyme XbaI (Thermo Fisher Scientific) and electrophoresis was performed on the CHEF Mapper™ System (Bio-Rad) (Electric field: 6 V/cm; Temperature: 14°C; Included angle: 120°; Initial switch time: 6.76 seconds; Final switch time: 35.38 seconds; and Run time: 19 hours). Salmonella Braenderup H9812 was used as molecular weight standard and analyses were performed using the BioNumerics v. 7.6 (Applied Maths, Belgium). UPGMA method and DICE similarity coefficient were used for constructing the similarity dendrogram (band position tolerance: 1.5% and optimization: 1.5%). Simpson's diversity index was used to evaluate the DI. 30
Multilocus sequence typing and fimH type
Multilocus sequence typing (MLST) analyzes were performed according to Achtman scheme using the housekeeping genes (recA, fumC, adk, gyrB, mdh, icd, and purA). The primers and conditions were available in Escherichia MLST website sited at the University of Oxford. 31 Phylogenetic analyses were performed using the PHYLOViZ 2.0 software. 32 The characterization of the fimH gene was performed using the primers and the web tool (FimTyper 1.0) 33 (Supplementary Table 4).
Results
E. coli strains
In this study, 17 E. coli strains were obtained from feces (n = 11) and soil samples (n = 6) from a Brazilian swine farm. All strains were resistant to more than one antimicrobial in more than three categories, being classified as MDR. The 16S rDNA sequences of E. coli strains were deposited in the GenBank with accession numbers MK951723-MK951739.
Antimicrobial resistance profile
All strains were resistant to ampicillin, cefuroxime, cefazolin, and streptomycin; 16 (94.1%) to trimethoprim-sulfamethoxazole, sulfonamide, and trimethoprim; 15 (88.2%) to tetracycline, doxycycline, gentamicin, and tobramycin; 13 (74.4%) to minocycline, 10 (58.8%) to cefaclor, and 8 (47%) to amikacin and chloramphenicol; 5 (29.4%) to ampicillin-sulbactam; 4 (23.5%) to nitrofurantoin; 3 (17.6%) to cefotaxime and ceftriaxone; and 2 (11.7%) to amoxicillin-clavulanate, piperacillin-tazobactam, ceftazidime, nalidixic acid, levofloxacin, ciprofloxacin, lomefloxacin, norfloxacin, and ofloxacin. All strains were susceptible to cefepime, aztreonam, meropenem, imipenem, and ertapenem (Table 1).
Antimicrobial resistance genotypes, plasmids, and virulence genes
Thirteen different ARGs were detected in MDR E. coli strains. All MDR E. coli strains harbored at least four ARGs investigated. All strains presented blaCMY, blaOXA-1-like, and tet(A), followed by tet(B) (n = 16), blaSHV (n = 15), sul2 (n = 11), aadA (n = 10), aac(6’)-Ib (n = 8), sul1 (n = 6), floR (n = 5), sul3 (n = 5), aph(3′)-Ia (n = 4), and cmlA (n = 4) (GenBank accession numbers MK959341-MK959353) (Table 1). For quinolones, no ARGs were detected; however, mutations in QRDR of GyrA (Ser83Leu; Asp87Asn) and ParC (Glu84Asp) were detected (i.e., A201 and A205) (GenBank accession numbers MK959320-MK959323). All MDR E. coli strains presented plasmids belonging to F, Y, and ColE-like, followed by FIB (n = 13), HI1 (n = 9), N (n = 8), FIA (n = 2), and FIC (n = 2) (Table 1). Three diarrheagenic virulence genes were detected in nine strains, such as ipaH (n = 6), stx2 (n = 2), and aaiC (n = 1), and thus, these MDR E. coli strains were classified as EIEC, STEC, or EAEC, respectively (Fig. 1).
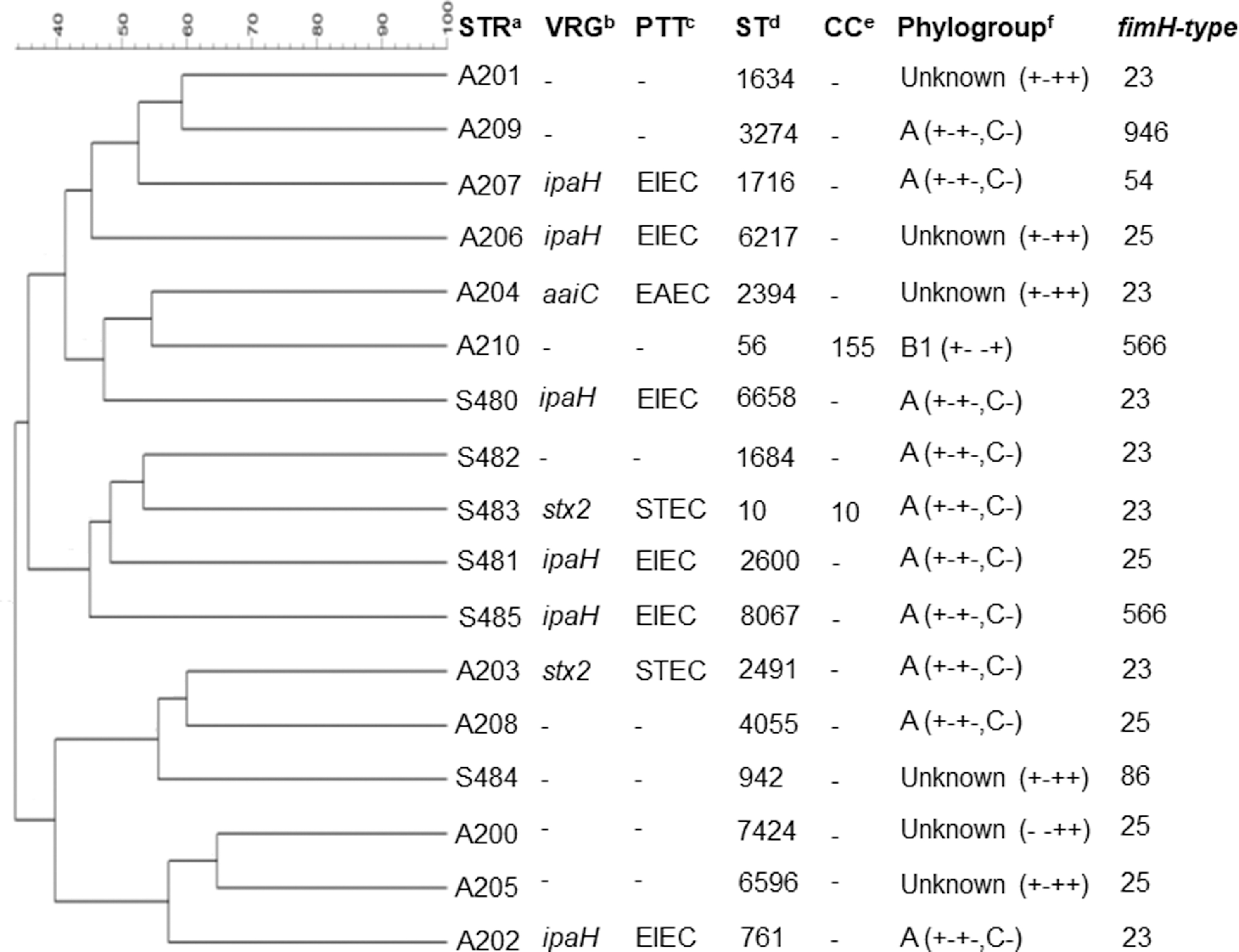
Dendrogram represents the genetic relatedness among the MDR Escherichia coli strains based on PFGE XbaI fingerprints.
E. coli typing and subtyping
PFGE showed a high genetic diversity among the MDR E. coli strains, which presented a similarity of 33.6% to 64.7%. For each strain, a different sequence type (ST) and clonal complex (CC) were determined, which were classified as ST10/CC10, ST56/CC155, ST761, ST942, ST1634, ST1684, ST1716, ST2394, ST2491, ST2600, ST3274, ST4055, ST6217, ST6396, ST7424, ST6658, and ST8067 (Figs. 1 and 2). Minimum-spanning tree was used for comparing the STs phylogenetically. The ST10/CC10 (S483) obtained from soil differed from ST1634 (A201), ST761 (A202), ST2491 (A203), and ST4055 (A208) obtained from feces and ST6658 (S480) obtained from soil by a single-locus variant. Other STs differed by two- to five-locus variants (Fig. 2). Regarding the determination of phylogroups, the majority was classified as A, followed by unknown and B1. Subtyping of the fimH gene showed six fimH type (23, 25, 54, 86, 566, and 946) (Fig. 1) (GenBank accession numbers MK959324-MK959340). The phylogroups A and Unknown, STEC and EIEC pathotypes, and fimH type (23, 25 and 566) were detected in both soil and feces samples.

MST based on PHYLOViZ software for comparing the STs phylogenetically. Genetic relationships are shown in MDR E. coli strains obtained from feces (dark gray) and soil (light gray). Representative STs are shown in bold. The numbers between circles show the number of allelic mismatches between STs. MST, minimum-spanning tree; ST, sequence type.
Discussion
MDR ARG-producing E. coli have been reported in several studies involving animal and environment samples; however, the studies performed on environmental samples mostly analyze water sources. Resistance to ampicillin, cephalosporins, streptomycin, and sulfonamides is a phenotype increasingly detected in E. coli obtained from different sources, including animals, which was also detected in this study. Soils present a great diversity of ARGs, being considered reservoir of these genes, and MDR ARG-producing E. coli have been previously detected in soil samples in Brazilian farms.34,35
The MDR profiles detected in this study are strongly related to the presence of encoding genes for β-lactamases (CMY, SHV, and OXA-1), which have penicillins and cephalosporins as preferred substrates; aminoglycoside-modifying enzymes [phosphotransferases (APH), nucleotidyltransferases (ANT), and acetyltransferases (AAC)]; and changed dihydropteroate synthase (Sul).36,37 Besides the MDR profile, many other E. coli strains are classified as MDR due to acquired efflux pumps (TetA/TetB and ClmA/FloR), which are associated with resistance to tetracyclines and phenicols, respectively, as well as by the presence of mutation in QRDR, leading to resistance to quinolones and fluroquinolones.38,39
CMY-type β-lactamase is the most common plasmid-mediated ampC gene in E. coli, which is able to hydrolyze extended-spectrum cephalosporins. 40 CMY-producing E. coli were recently reported in different sources, including humans, animals, and food. 41 The majority of the ARGs found in this study are commonly carried by different plasmids, mainly IncF. 42 DEC can be classified in some pathotypes, which are related to diarrheagenic diseases in humans and animals. STEC, EIEC, and EAEC E. coli produce Shiga toxins (stx1/stx2), invasion plasmid antigen H (ipaH), and aggR-activated island C (aaiC), respectively, and have been reported causing infections and outbreaks worldwide. 43 Kusumoto et al. 44 reported MDR ARGs-producing STEC in swine samples, corroborating with the results found in this study.
Genetic relatedness by PFGE demonstrated a high genetic diversity in swine and environmental MDR E. coli strains, a profile also observed in other studies.44,45 Regarding the phylogenetic groups, there was a high prevalence of phylogroup A and low prevalence of B1, corroborating with the phylogenetic groups detected by Asai et al. 46 According to Enterobase Database, some STs have already been detected in swine (ST56, ST761, ST1716, and ST3274) and soil (ST10 and ST942); however, it is the first report of ST1634, ST2394, ST2491, ST4055, ST6217, ST6396, and ST7424 in swine, and ST1684, ST2600, ST6658, and ST8067 in soil. Interestingly, no ST detected in fecal samples was detected in the soil, showing that E. coli are able to survive in environmental samples. 47
Eight STs differed from ST10/CC10, an ST worldwide disseminated, by one- to three-locus variants, showing a genetic evolution of these strains. MDR E. coli lineages belonging to CC10 and CC155 have been reported worldwide, mainly in infections in humans. 48 The FimH protein, codified by the fimH gene, is an adhesin responsible for immobilizing E. coli on different surfaces. As the fimH gene has a low mutation rate, it is being used for high-resolution subtyping of MLST of E. coli. Thus, it is possible to differentiate E. coli strains within the same ST/CC and the alleles of the fimH gene detected in this study can be used to compare E. coli strains from several sources.33,49
Conclusion
This study shows a high genetic diversity among MDR ARG-producing E. coli belonging to STEC, EIEC, and EAEC pathotypes obtained from a small-scale pig farm, which is worrying since it can disseminate to other sources and, consequently, cause diseases and spread ARGs for other pathogens. In addition, these results also contribute to the monitoring of antimicrobial-resistant pathogens worldwide, mainly in environmental samples, which are associated with One Health framework.
Footnotes
Acknowledgment
The authors thank the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) [grant number 2018/01890-3] and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) [grant numbers 88882.180855/2018-01 and Finance Code 001] for fellowships.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by São Paulo Research Foundation - FAPESP [grant number 2018/19539-0].
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.