
Abstract
The genetic mechanisms associated with acquisition of linezolid (LZD) resistance are diverse, including point mutations in the V domain of the 23S rRNA and the 50S ribosomal proteins as well as cfr, optrA, and/or poxtA genes, which may be plasmid- or chromosomally encoded. The aim of this study was to investigate through Whole Genome Sequencing (WGS)-based typing the presence and location of genes and point mutations associated with LZD resistance in two Enterococcus faecalis isolates from Upper Austrian patients. The isolates were retrieved during screening by LZD disk diffusion test of a total of 911 clinical E. faecalis isolates in 2017. The two E. faecalis isolates had LZD minimum inhibitory concentrations of 8 and 32 mg/L and were optrA-positive (ST476 and ST585). Bioinformatic analysis revealed the presence of optrA located in the chromosome of both isolates. One isolate carried the optrA gene in the transposon 6674, previously reported as chromosomally encoded, and the second isolate in fragments originating from the integrative plasmid pEF10748. Additional mechanisms of LZD resistance on the 23S rRNA and the 50S ribosomal proteins were detected. None of the patients reported travels to geographical areas with high LZD resistance or previous LZD treatments. This is the first report of optrA carrying E. faecalis, including characterization by WGS from Austria. LZD resistance in a low-prevalence setting is of concern and should be further monitored.
Introduction
E
Although LZD resistance in Austria is low, 15 nosocomial outbreaks due to LZD resistant enterococci (LRE) occur. 16 In 2017, we detected two LZD-resistant E. faecalis isolates during routine testing at the National Reference Center for Antimicrobial Resistance and Nosocomial Infections (NRZ). The aim of this study was to analyze the resistance mechanisms and genetic environment of the LRE isolates by Whole Genome Sequencing (WGS).
Materials and Methods
Bacterial isolates and antimicrobial susceptibility testing
In 2017, 911 clinical E. faecalis isolates underwent routine susceptibility testing at our institution. The clinical samples had been taken at four different hospitals in Upper Austria as well as at several practitioners. They also included surveillance stool cultures for hemato-oncological patients, where one stool sample per week was screened for multidrug-resistant bacteria (Gram negatives, VRE) by using chromogenic screening media. In addition, a selective medium for enterococci was included and each phenotypically distinguishable strain was tested with our antimicrobial susceptibility testing routine panel, including LZD. Two of the 911 isolates (0.2%) (isolates A and B) were found to be LZD resistant by the disk diffusion test according to EUCAST criteria. 17
Minimum inhibitory concentrations (MICs) were determined for LZD and tedizolid (TZD) by using Liofilchem test strips (Liofilchem, Roseto degli Abruzzi, Italy) and E-test (Biomérieux, Marcy L'Étoile, France) methodologies. Species identification was confirmed by matrix assisted laser desorption ionization-time of flight mass spectrometry (Bruker Daltonics, Hilden, Germany).
Molecular characterization
Testing of the two LZD-resistant E. faecalis isolates for the presence of optrA and cfr was performed by using previously published primer sets. 11 In addition, high-molecular-weight DNA from E. faecalis cultures was isolated by using a MagAttract HMW DNA kit (Qiagen, Hilden, Germany), followed by DNA quantification with a DropSense 16 (Trinean NV/SA, Gentbrugge, Belgium) device. Library preparation was done with a NexteraXT kit (Illumina, Inc., San Diego, CA) following the manufacturer's instructions, and a 300-bp paired-end sequencing run was carried out by using an Illumina MiSeq device (Illumina, Inc.). The raw reads were de novo assembled with SPAdes version 3.11.1 into contigs. 18 For each isolate, we used SeqSphere+ software (Ridom GmbH, Münster, Germany) to extract the optrA gene sequences and the Sequence Types (STs) based on classical Multi Locus Sequence Typing (MLST). 19 In addition, the genetic relatedness between the two isolates was assessed by core genome MLST. 20
The optrA gene sequences were aligned with the optrA sequence of the reference genome E. faecalis E394 (NG_048023.1) by using BLAST and the LRE identifier tool. 21 To find out whether the optrA gene was located in a plasmid or in the chromosome, we extracted plasmids by using Plasmid Finder and compared the contigs containing a plasmid with the contig carrying the optrA gene. 22 In addition, isolates were screened for the presence or absence of transposons Tn554, Tn558, and Tn6674, described as being associated with the chromosomal carriage of optrA.23,24
To further confirm the location of optrA, both strains were sequenced by using an Oxford Nanopore MinION device following the standard protocol provided by the manufacturer for library preparation. Long reads (MinIon) and short reads (Illumina) were used to create hybrid assemblies with Unicycler v.0.4.8 by using default parameters. The annotation was performed by using BLAST and Prokka (v.1.14.5), and “DNA Features Viewer” was used for visualization of the genetic context. 25
We looked for known point mutations conferring LZD resistance located in any of the four 23s rRNA genes (such as G2576T and G2505A) or in the ribosomal protein genes rplC (L3), rplD (L4), and rplV (L22). To do so we simply extracted with Seqsphere+ the sequence of the genes mentioned earlier from the WGS data and aligned them with those of the E. faecalis V583 reference genome (NC_004668.1). We assessed the presence of other resistance genes not linked to LZD resistance and the main virulence genes by using CARD and VirulenceFinder databases.26,27
Patient data and ethics approval
Patient data were retrospectively obtained by chart review. The study was approved by the local ethics committee (vote ECS 1053/2018).
Results
Antimicrobial susceptibility testing and PCR
Disk diffusion testing yielded two LZD resistant E. faecalis isolates. Gradient testing revealed MICs of 8 and 32 mg/L, respectively, for isolates A and B. MICs for TDZ were 2 and 4 mg/L, respectively. The presence of optrA and the absence of cfr genes were shown by conventional PCR.
Epidemiological data
Isolates A and B had originated from two patients treated in two different hospitals in Linz, Upper Austria. Isolate A was cultured from a wound swab of a 34-year-old male patient in October 2017. He had been treated at a trauma ward in hospital 1 for an occupational foot injury. His antimicrobial therapy during the previous 4 weeks included clindamycin, moxifloxacin, vancomycin, and amoxicillin-clavulanate, but not LZD. After multiple surgeries, his foot had to be amputated because of the severity of his trauma, and no more LZD-resistant E. faecalis were detected.
Isolate B was identified in a routine surveillance stool culture of a 68-year-old female stem cell transplant recipient treated for acute graft versus host disease in hospital 2 in November 2017. She had not received LZD during the past 4 weeks and weekly screening for LRE during previous inpatient stays during the past year had been negative. In the following weeks, the optrA positive E. faecalis could not be detected anymore in multiple stool surveillance cultures nor in other clinical materials. The patient died from non-infectious causes 3 months later.
Infection control measures, including isolation of the patients in single rooms, were initiated in both cases. No additional cases were detected in specimens received from other patients treated at these wards.
WGS-based typing
The raw sequencing coverage of isolate A was 47 × with Illumina Miseq reads and 160 × with Oxford Nanopore MinION reads. The coverage of Isolate B was 73 × with Illumina MiSeq reads and 159x with Oxford Nanopore MinION reads.
The hybrid assemblies resulted in close-to-complete genome drafts for isolate A and isolate B.
Isolate A was assembled into one large chromosome of 2,728,657 bp and three circular contigs (62,940, 38,725, and 10,667 bp), which were predicted to be plasmids. The chromosome of isolate B was assembled with 2,835,786 bp, together with two circular contigs (75,364 and 5,437 bp), which were predicted to be plasmids.
We confirmed that isolate A was ST476 and isolate B was ST585. They differed by 2027 alleles in their core genomes when using the short reads assembly and by 2035 alleles when using the hybrid assemblies, and both carried the optrA gene, PlasmidFinder identified in isolate A one rep_trans (repUS43) and one repA_N (rep9a) plasmid. For isolate B, two rep_trans (rep7a and repUS43) and one repA_N (rep9a) plasmids were identified. None of the contigs containing those plasmids harbored the optrA gene. Also, independently of the type of assembly (short reads vs. hybrid assembly), plasmid types found in the isolates were the same.
Isolates A and B carried the optrA variants 5 and 3, respectively. Neither poxtA gene nor cfr was detected but other resistance genes not linked to LZD resistance as well as virulence genes were detected (Table 1). When using Seqsphere and BLASTn, we detected in both isolates a point mutation located at nucleotide 301 of the rplD gene (encoding for the L4 50S protein), leading to an aminoacidic change at codon 101 (F101L). Isolate B carried a silent mutation at codon 40 (C120T) of the rplV gene encoding for L22. No other mutations were detected in any of the two isolates at rplC gene, encoding for L3 protein. Also, three different point mutations in at least one of the four copies of the 23s RNA genes were detected in each isolate. LRE-Finder detected no point mutations at all linked to LZD resistance. Again, no differences in the results were seen when using different assembly types.
Point Mutations, Virulence Genes and Additional Resistance Genes Found in the Sequenced Enterococcus faecalis Isolates
Compared with the reference sequence Enterococcus faecalis E394 (NG_048023.1).
New mutation. Numbers in between parentheses indicate the mutated 23s RNA copies out of the four copies present in E. faecalis.
ST, sequence type.
Regarding the location of optrA, it was contained in mobile elements within the chromosome of both isolates. The chromosome of isolate A contained 98.5% of Tn6674 (accession no.: MK737778.1) and shared an identity of 99.8% with the transposon. Isolate B contained five fragments (sizes varying from 3,067 to 17,211 bp) of the recently described plasmid pEF10748 (accession no.: MK993385.1) integrated into the chromosome. 28 This represented 97.6% of the plasmid pEF10748 and an average identity of 99.3% (Fig. 1).
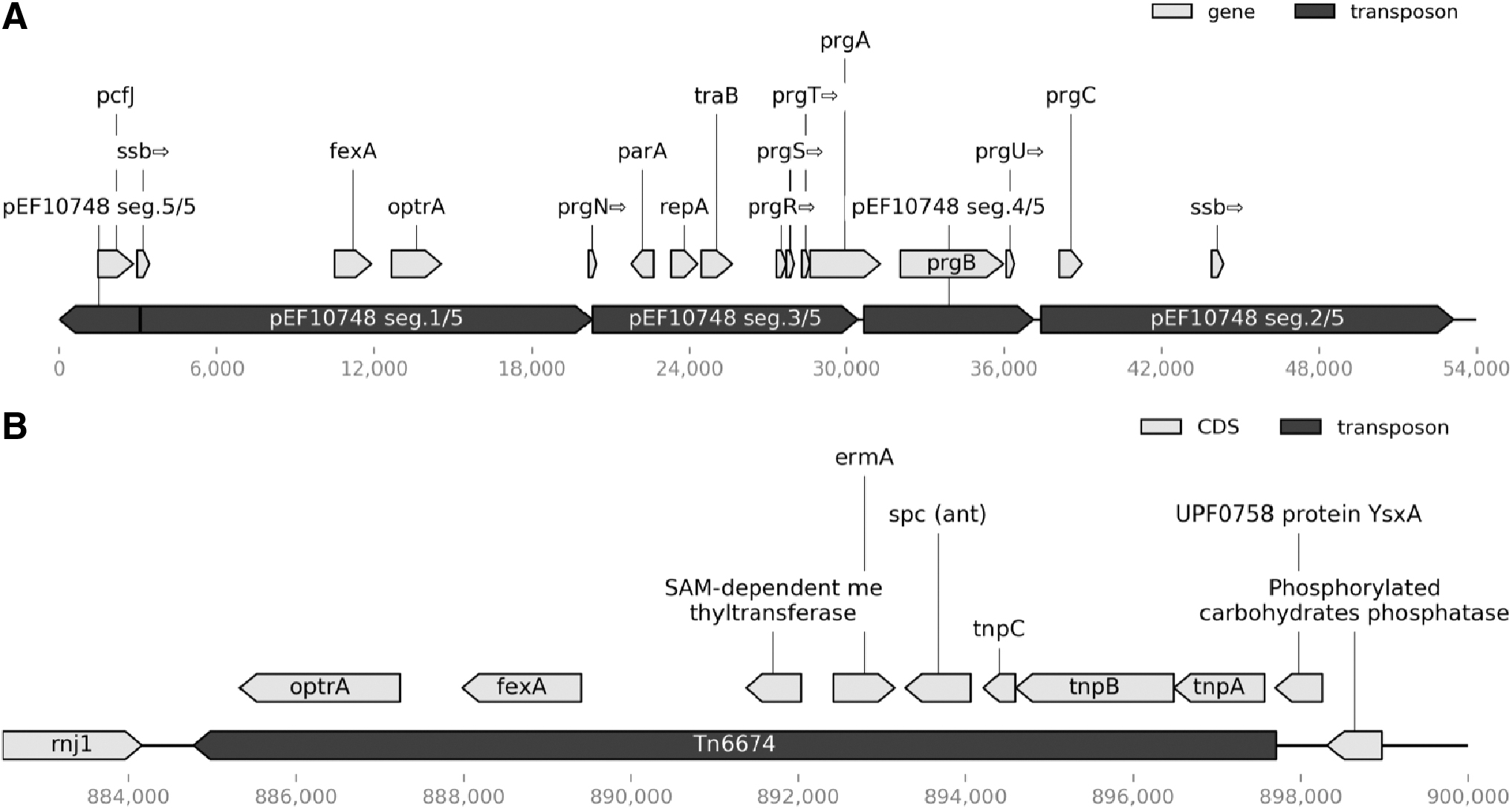
Genetic context of optrA genes detected in both isolates. For better visibility, hypothetical genes were hidden.
Discussion
We present here the first characterization by WGS of optrA-mediated LZD-resistant E. faecalis isolates in Austria. To the best of our knowledge, this resistance mechanism appears to be rare in Austria, with only one previous conference report. 29 Compared with reports from the United States and a multicenter study screening a large sample of enterococcal isolates for LZD resistance, the prevalence of 0.2% in our sample is similar.30,31 In contrast, in other countries the detection of LRE is frequently reported both in humans and livestock as well as in meat products,9,32 drawing a link between optrA-carrying bacterial isolates and an animal reservoir. In our study, no contact of the patients with livestock could be established and oxazolidones are not used in veterinary medicine in Austria. However, other antimicrobials such as florfenicol are applied to treat livestock and pets, hence optrA may be responsible for florfenicol resistance too. 6 Previous LZD treatments are a known risk factor for selection of LRE, 33 however no treatment with LZD had been administered in our patients. Indeed, LZD consumption is stable and relatively low in Austria, with a reported use of about 0.5 defined daily doses per 100 bed-days between 2011 and 2017. 15
Although each isolate contained at least two plasmids, none of them carried the resistance gene optrA. The hybrid assembly constructed with Oxford Nanopore reads using Unicycler confirmed the chromosomal origin of optrA in both isolates.
Moreover, isolate A was of the same ST, optrA variant, and rplD mutation (ST476, optrA4, T301C) as an LRE isolate recovered from chicken meat in Tunisia (isolate 697T), in which optrA was also chromosomally located, and which differed by 102 alleles from isolate A. 34 Similarly, studies in China reported chromosomally located ST476 E. faecalis obtained from human samples, with one of them being optrA4-positive too.9,35 In addition, the transposon Tn6674 found in isolate A has been detected in E. faecalis isolates carrying optrA gene in the chromosome. 35
Isolate B was of ST585 and optrA_3, the same ST as three E. faecalis strains found in a Spanish study 31 and the same variant as in previous studies. 35 In contrast, our ST585 LZD-resistant E. faecalis seems to carry optrA chromosome-encoded, whereas other studies have mostly reported this gene within plasmids.23,28,34,36 However, in the study of Freitas et al., these ST585 isolates were also optrA_3 and among the plasmids identified, one was from the family RepA_N as well. 35 Although previous works reported on rep9a plasmids carrying optrA, 28 bioinformatics analysis here evidenced the opposite. In Europe, E. faecalis of other STs such as ST16 have been also reported as carriers of optrA in the chromosome. 37 However, information on the epidemiology of E. faecalis is still scarce.
The identified point mutation at rplD gene (F101L) was already described in countries such as China and Mexico in E. faecalis and Staphylococcus epidermidis.13,38 Regarding its possible role in LZD resistance, there were some discrepancies between authors, since some claimed that it may contribute to the resistance,9,35 others that it is not strongly associated to LZD resistance 13 or even found in LZD-susceptible isolates only. 39 Nevertheless, when this mutation was the only explanation for LRE resistance, the MIC concentrations were always over the break-point of 8 mg/L. 9 In the case of the C120T mutation at the rplV gene, due to its silent effect no contribution to the LZD resistance was expected.
Only one non-previously identified 23S RNA mutation was detected (T2802 C/I934M), which due to the aminoacid change might have some impact on LZD resistance. The other two (G388A/D130N and T2838C/-) were previously reported in Tunisia in both chicken meat and feces and they were associated with LZD resistance. 34
Isolates A and B carried mostly the same resistance genes as the ST476 and ST585 E. faecalis from the study of Freitas et al. 35 The efrA gene, associated to macrolide resistance, was present in both of our isolates and absent in the study of Freitas et al. whereas the ermB gene, associated with erythromycin resistance, was absent in our ST585 isolate but present in the ST585 isolates from Freitas et al. Similarly, the virulence gene cbh, which is part of the Pathogenicity Island of E. faecalis, was not detected in our ST585 isolate.
We can conclude then that the optrA gene was most probably the only genetic element contributing to the LZD resistance detected in both of the isolates, and that the optrA genetic variant may play a role in the different MICs detected. Our results contribute to a better understanding of the molecular mechanisms behind LZD resistance in Austria, which can have an influence on the efficacy of the antimicrobial treatment. Therefore, continuous monitoring of LRE strains together with their genetic resistance mechanism is advised.
Footnotes
Acknowledgments
The authors thank Anna Blaimschein, Bernadette Hones, and Sarah Widhalm for excellent technical work and Markus Sinkovics (Unfallkrankenhaus Linz) for his help in obtaining clinical data. Tedizolid E-Tests were generously donated by Merck Sharp & Dome, Vienna.
Disclosure Statement
No competing financial interests exist.
Funding Information
No external funding was received for this study.