
Abstract
Objective:
Antibiotic resistance of Pseudomonas aeruginosa (PA) that lowers the effectiveness of current treatments for pneumonia is a growing problem. Qi Gui Yin is a Chinese herbal medicine that has been used to improve the efficacy of antibiotic therapy against antibiotic-resistant bacteria. This study aimed to elucidate the mechanism by which Qi Gui Yin inhibits antibiotic resistance of PA.
Methods:
Active components of Qi Gui Yin were analyzed by chromatography. Isobaric Tags for Relative and Absolute Quantification (iTRAQ) technology was used to compare protein expression profiles of PA strains cultured in serum from rats that were and were not treated with Qi Gui Yin. Quantitative polymerase chain reaction (qPCR) analysis was performed to detect gene expression changes.
Results:
Proteomic analysis identified 76 differentially expressed proteins between PA strains cultured in serum from rats that were or were not treated with Qi Gui Yin. Bioinformatics analysis revealed that the largest number of differentially expressed proteins were associated with resistance mechanisms such as quorum sensing, bacterial biofilm formation, and active pumping. In addition, qPCR analysis confirmed that downregulation of iscU and arcA gene expression was associated with Qi Gui Yin treatment.
Conclusions:
Serum from Qi Gui Yin-treated rats could effectively inhibit antibiotic resistance of PA. Chlorogenic acid and astragaloside IV are the main components of Qi Gui Yin, which may mediate inhibition of antibiotic resistance. Our findings provide new insights into strategies involving Chinese herbal medicine that can be used to treat pneumonia caused by antibiotic-resistant bacteria.
Introduction
P
Medicinal plants have long been used as agents to treat infectious diseases. 6 Qi Gui Yin is a Chinese herbal medicine mixture that includes Radix Astragalus, Radix Angelicae Sinensis, Flos Lonicerae, Artemisia annua, and Rhizoma Polygoni Cuspidati, which may be able to significantly improve the efficacy of antibiotic therapy for antibiotic-resistant bacteria. 7 In a previous study, we demonstrated that Qi Gui Yin induced PA antibiotic resistance by stimulating production of protective antibodies against VIM-1, SPM-1, and TEM-1, which are involved in drug resistance. 8 In this study, we aimed to elucidate the mechanism by which Qi Gui Yin inhibits the resistance of PA to antibiotics such as imipenem (IPM).
Herbal mixtures have complex components and the in vivo metabolism of these compounds is dynamic. As such, serum isolated from animals treated with herbal medications may better mimic the in vivo situation and in turn more faithfully reflect the pharmacologic effects of herbs. In this study, we used Isobaric Tags for Relative and Absolute Quantification (iTRAQ) technology to compare protein expression profiles of PA strains cultured in serum from rats that were or were not treated with Qi Gui Yin. We identified target proteins of Qi Gui Yin that may mediate the inhibition of antibiotic resistance. Our findings provide new insights into targeted therapies involving Chinese herbal medicines to treat pneumonia caused by antibiotic-resistant bacteria.
Materials and Methods
Animals
All animal experiments were approved by the Animal Use and Care Committee of the Beijing Traditional Chinese Medicine Research Institute. Specific pathogen-free male Sprague-Dawley rats (6–8 weeks old) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. and housed under a 12-hour light/12-hour dark cycle at a constant temperature of 20°C ± 4°C, with ad libitum access to food and water. The rats were randomly divided into a control group (group C) and an experimental group (group E; n = 6 for each group).
Instruments, reagents, and chromatographic analysis
Qi Gui Yin was supplied by the College of Chinese Medicine, Beijing University of Traditional Chinese Medicine (Lot No. 216–002); Tienam was produced by Hangzhou Merck East Pharmaceutical Co., Ltd. (Lot No. 17850). Mueller-Hinton (MH) medium, MH agar plate (XWO, 1701256), and IPM antimicrobial susceptibility disks (Batch No.2154386) were purchased from OXOID (Basingstoke, Hampshire, UK). Chromatographic column parameters were determined as described previously. 9 Standard samples and Qi Gui Yin were prepared using routine methods. Standard concentrations of astragaloside IV, chlorogenic acid, emodin, and polydatin were 413, 45, 48, and 49.4 μg/mL, respectively. The amount of these four molecules in Qi Gui Yin was analyzed by chromatography.
Serum collection
The day on which the two groups of rats received the first intragastric gavage containing either Qi Gui Yin or placebo was considered day 1. Rats in Group E were administered 2 mL Qi Gui Yin (based on a dose of 10 g/kg body weight), and rats in group C were administered the same volume of distilled water. Each rat received the gavage once a day. On day 5, 1 hour after intragastric administration, blood samples from the two groups were collected from the abdominal aorta and centrifuged at 5,000 rpm for 10 minutes. Serum samples from groups C and E were labeled as serum C (SC) and serum E (SE), respectively, and frozen at −80°C for later use.
Drug sensitivity test
The clinically isolated PA strain number PA2038 was identified and validated. For the untreated control group (PA-M), 10 μL of bacteria (turbidity value 0.3) was inoculated into 300 μL MH medium. For the serum control group (PA-C), 10 μL of bacteria was cultured in 240 μL MH medium supplemented with 60 μL SC, and for the experimental group (PA-E), 10 μL of bacteria was cultured in 240 μL MH medium supplemented with 60 μL SE. Both PA-C and PA-E groups were treated with IPM (Tienam, 0.016 μg/μL). Bacteria for all three groups were cultured at 36°C ± 1°C and the cultures were passaged once every 24 hours when the optical density reached 0.3. A conventional IPM sensitivity test was performed at passages 3 and 6 (P3 and P6) using a disk diffusion method as described previously. 10
According to CLSI (2018 version) standards, the resistance criteria were as follows: resistant (R): ≤15; intermediate (I): 16–18; and sensitive (S) ≥19. PA from P3 and P6 of each group were collected, quantified, and frozen at −80°C.
Protein extraction and digestion
Protein was extracted from bacteria using a Total Protein Extraction Kit (Bangfei Bioscience, Beijing, China) and the protein concentration was determined using the Bradford method with bovine serum albumin as the standard. 11 The Filter Aided Sample Preparation (FASP) protocol was used for protein digestion. 12 For all samples, three biological replicates were analyzed.
iTRAQ detection
Samples were prepared and labeled with an iTRAQ kit according to the manufacturer's instructions. Briefly, 200 μg of bacterial protein was taken from each group and transferred to a centrifuge tube to which 5 μL 1 M dithiothreitol (DTT) was added. The mixture was incubated at 37°C for 1 hour before 20 μL 1 M indoleacetic acid (IAA) solution was added, mixed thoroughly, and incubated in the dark at room temperature for 1 hour.
Then, the mixture was transferred into an ultrafiltration tube and centrifuged. The filtrate was discarded and 100 μL urea (UA) was added, and the mixture was again centrifuged and the filtrate was discarded. This step was repeated twice. Then, 100 μL 0.5 M tetraethylammonium bromide (TEAB) was added before the mixture was centrifuged and the filtrate was discarded. This step was repeated thrice. Finally, the samples were digested with trypsin at 37°C for 14 hours. Around 100 μg of peptides from each group was labeled with an iTRAQ Reagent-8 plex Multiplex Kit (AB SCIEX) and used for liquid chromatography with tandem mass spectrometry. High performance liquid chromatography grading was performed using C18 columns (1.9 μm; 0.15 × 120 mm) under alkaline pH conditions, followed by mass spectrometry.
Quantitative polymerase chain reaction
Quantitative polymerase chain reaction (qPCR) was performed to detect the gene expression of arcA and iscU according to standard protocols using the following primers: arcA 5′-CCCAAGCTTTCCTGACTGTACTAACGG-3′ 5′-CGCGGATCCTTAATCTTCCAGATCACC-3′; iscU 5′-CATGCCATGGCATACAGTGAAAAGGTGATC-3′; and 5′-TGACAAGCTTGTGTGCCGCCTGGGCACT-3′.
Data analysis
Data are presented as mean ± standard derivation and were analyzed using SPSS version 12 (SPSS, Inc., Chicago, IL). p < 0.05 was accepted as statistically significant. The distribution of peptide length indicated the frequency distribution of proteins having different sizes in a given sample. Fold change of the differential protein expression (with 1.5-fold upregulation or downregulation) and p-values of t tests between groups were used to generate a volcano plot. Raw mass spectrometry data were searched against the Uniprot database (P. aeruginosa, 173,814 sequences) using SEQUEST software (Proteome Discoverer 1.4; Thermo Fisher Scientific, San Jose, CA). Biological function as well as molecular and cellular biology of the differentially expressed proteins were examined by Gene Ontology (GO) analysis using the tool “WEGO”. 13 Protein signaling pathways were analyzed by Kyoto Encyclopedia of Genes and Genomes GENES (including protein information) and KEGG PATHWAY.
Results
Chromatography analysis of Qi Gui Yin
The four ingredients of Qi Gui Yin, astragaloside IV, chlorogenic acid, emodin, and polydatin, were analyzed by chromatography (Fig. 1). The concentration of astragaloside IV and chlorogenic acid was higher than those in the standard samples, consistent with previous reports that chlorogenic acid and astragaloside IV derivatives are associated with inhibition of PA drug resistance.14,15

Chromatographic analysis of Qi Gui Yin components. Chromatographic analysis of
Sensitivity analysis of IPM
The third generation (P3) of the PA strain was resistant to IPM in conventional serum-free broth medium (Fig. 2). Upon addition of rat serum to the broth medium, resistance to IPM was not observed until the sixth generation (P6). Serum from rats treated with Qi Gui Yin was more effective in delaying resistance of PA to IPM, such that the sixth generation of the PA strain showed no IPM resistance.

Antibiotic resistance characteristics of PA in different culture mediums with or without serum from rats treated with Qi Gui Yin. −, Resistant to imipenem; +, sensitive to imipenem. PA, Pseudomonas aeruginosa. Color images are available online.
Differentially expressed proteins between group PA-C and PA-E
To elucidate the mechanism by which Qi Gui Yin could inhibit IPM resistance, protein expression profiles of PA strains cultured in serum from rats with (PA-E) or without (PA-C) Qi Gui Yin treatment were examined. After digestion with trypsin, the length of peptides from both PA-C and PA-E samples was mainly between 6 and 26 amino acids, with prominent peaks for peptides having between 9 and 12 amino acids (Fig. 3A).
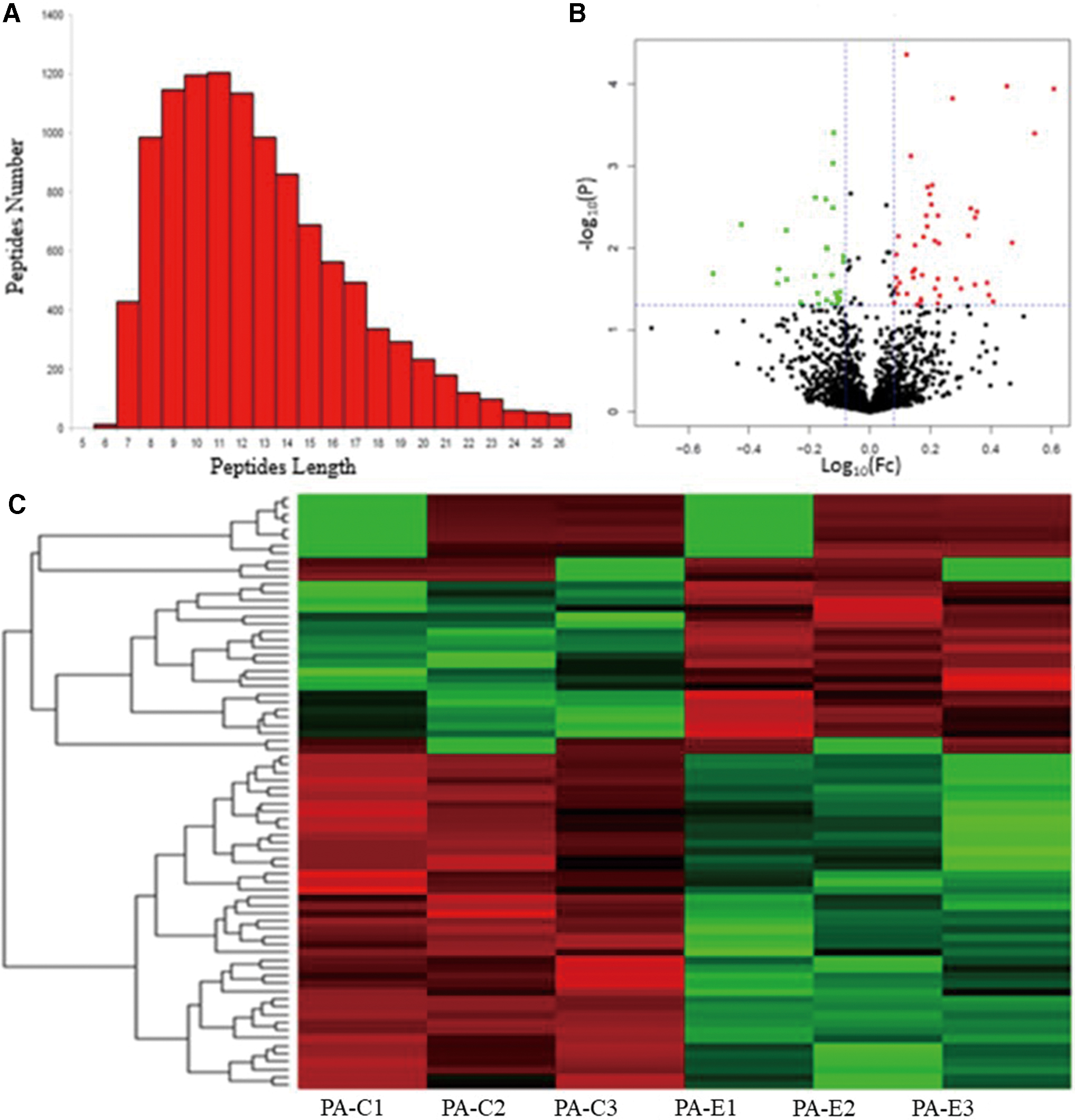
Differential protein expression between PA-C and PA-E groups.
The overall differential expression of the proteins between the PA-C and PA-E groups was examined using a volcano plot (Fig. 3B) and a heat map displaying results of cluster analysis of differentially expressed proteins (Fig. 3C). Among the differentially expressed proteins, 47 and 29 were downregulated and upregulated, respectively (Tables 1 and 2).
Proteins Showing Downregulated Expression in PA-E Relative to PA-C
Proteins Showing Upregulated Expression in PA-E Relative to PA-C
GO analysis and verification by qPCR
GO analysis showed that the differentially expressed proteins were mainly involved in biological processes such as biological adhesion, carbon utilization, and response to stimuli. In addition, some proteins were associated with cell constitution and cell membranes, and others carry out molecular functions such as molecular transduction and transport. Several proteins had roles in cell structure. Expression of both ArcA, an important protein for PA biofilm formation, and IscU, which is involved in aerobic growth of PA, was downregulated by serum from Qi Gui Yin-treated rats (Fig. 4), consistent with the results for protein expression and PA drug resistance status.
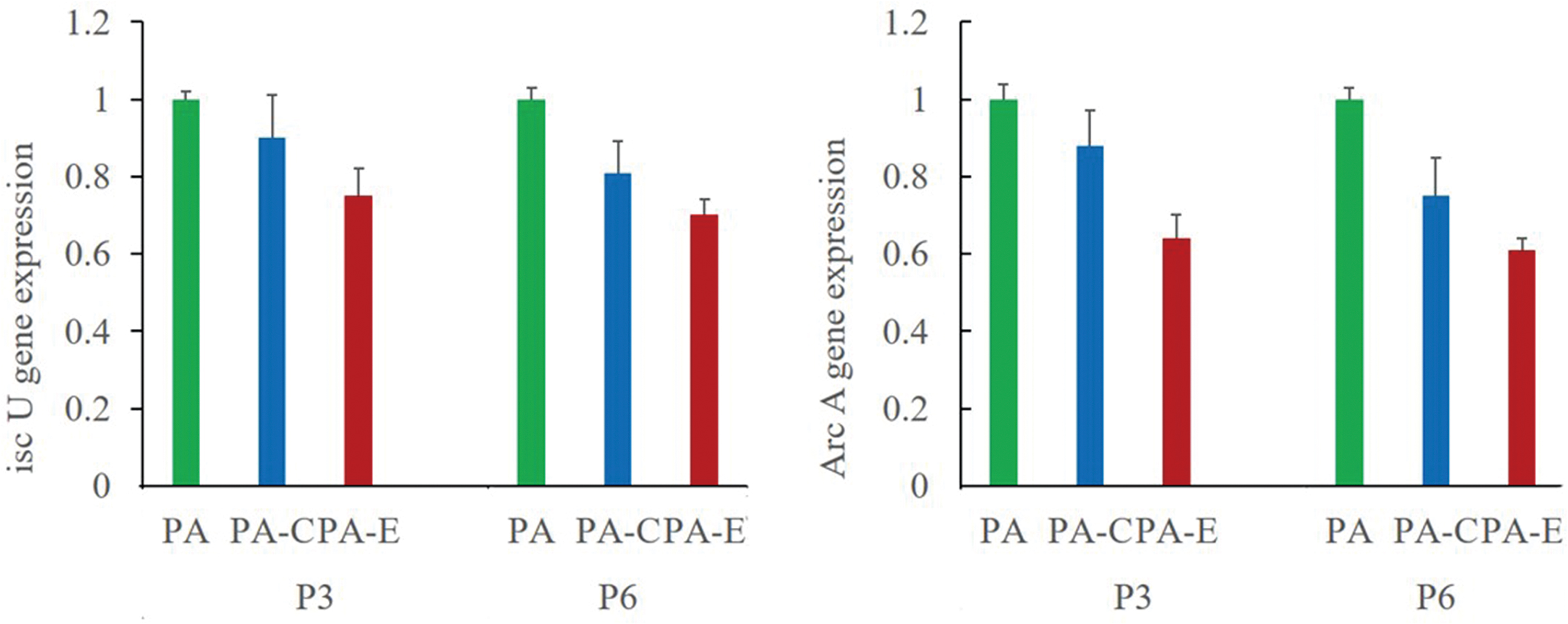
ArcA and IscU gene expression levels in different groups of PA. Data are presented as mean ± standard derivation (n = 3). Color images are available online.
Enrichment and KEGG analysis
Further enrichment analysis revealed that the differentially expressed proteins are mainly involved in the biological processes transmembrane transport activity, structural constituents of ribosomes, and antibiotic metabolism (Fig. 5A). Together, the differentially expressed proteins could affect these biological processes to contribute to drug resistance of PA.
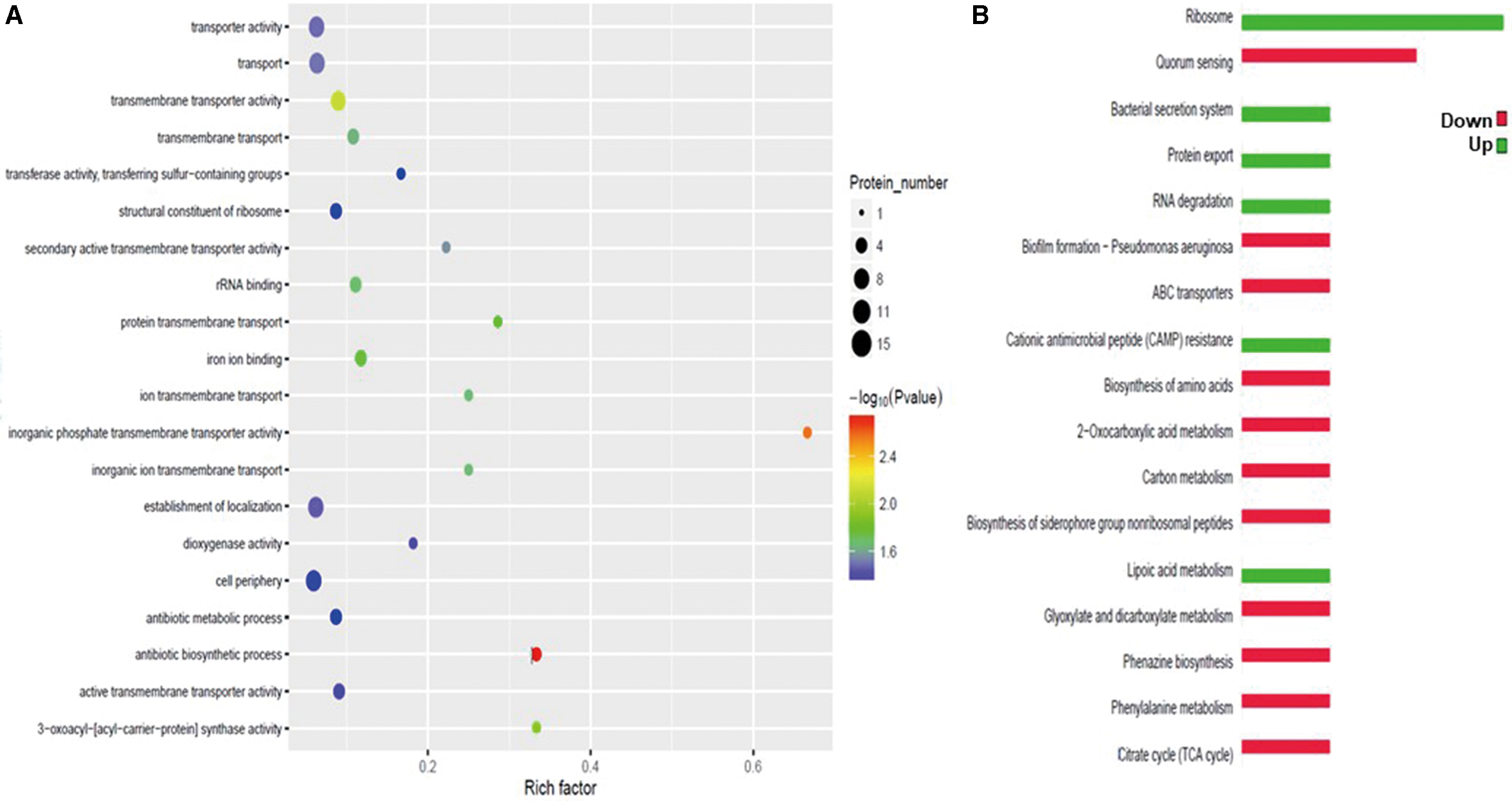
Enrichment and KEGG pathway analysis of proteins showing differential expression between PA-C and PA-E.
KEGG pathway annotation analysis revealed that the differentially expressed proteins mainly participated in biological functions associated with ribosomes, quorum sensing, bacterial secretion system, and PA biofilm formation. Expression of proteins related to ribosomes, bacterial secretion systems, protein export, and cationic antimicrobial peptide (CAMP) resistance was downregulated, while expression of proteins related to quorum sensing, biofilm formation, and amino acid biosynthesis was upregulated (Fig. 5B).
Discussion
PA is a common pathogen of pneumonia. The increasing incidence of resistance, together with characteristics of PA drug resistance, has complicated the clinical treatment of pneumonia.16,17 Therefore, novel approaches to overcome antibiotic resistance of PA are urgently needed. 18
Although IPM is the first choice for clinical treatment of infectious pneumonia due to its superior antibacterial activity and broad antibacterial spectrum, some features of IPM induce resistance in PA and the frequency of IPM-resistant strains are increasing.19–23 In this study, we found that serum from rats treated with the herbal mixture Qi Gui Yin effectively delayed onset of PA resistance to IPM, indicating that use of Qi Gui Yin could prolong the clinical utility of IPM for the treatment of pneumonia, which is consistent with findings by previous studies.7,8 Also, we confirmed that astragaloside IV and chlorogenic acid are abundant in Qi Gui Yin. Chlorogenic acid has a direct antibacterial effect, whereas astragaloside IV has an indirect antibacterial effect mediated by its metabolic derivatives.14,15 The mechanisms underlying PA resistance to antibiotics include biofilm formation, induction of quorum sensing, promotion of efflux pump activity, and reduction in bacterial outer membrane permeability.20,24–29
To identify potential target proteins of Qi Gui Yin involved in PA resistance, in this study, we employed iTRAQ technology to the compare protein expression profile of PA cultured with serum from rats that were or were not treated with Qi Gui Yin. Our results demonstrated that upon culture with serum from Qi Gui Yin-treated rats, 76 proteins having either known or unknown functions were differentially expressed compared to PA cultured with serum from untreated control rats. Of these proteins, 29 and 47 showed upregulation and downregulation, respectively. Findings from GO analyses suggested that Qi Gui Yin could regulate antibiotic resistance of PA by altering expression levels of proteins such as ArcA and IscU, which are involved in bacteria biofilm formation.
Furthermore, KEGG pathway analysis indicated that Qi Gui Yin impacted antibiotic resistance of PA by altering expression of the proteins PhzA, PhzB, PhzM, MetQ1, ArcA, IscU, lpsJ, and PilA, which are individually or synergistically involved in regulation of PA quorum sensing, the bacterial secretion system, bacterial biofilm formation, and CAMP resistance. In particular, PhzA, PhzB, PhzM, MetQ1, ArcA, and IscU could promote quorum sensing and biofilm formation of PA to promote antibiotic resistance.30,31 Downregulation of the expression of these proteins in PA cultured in serum from Qi Gui Yin-treated rats could render PA sensitive to IPM. On the other hand, PilA, PilJ, and LpsJ are related to virulence and pathogenicity of PA.32–34 Upregulation of these proteins in PA cultured in serum from Qi Gui Yin-treated rats is expected to increase bacterial virulence. This apparent paradox could be explained by the complex relationship between bacterial virulence and antibiotic resistance. 35 The downregulation of ArcA and IscU expression may contribute to Qi Gui Yin-mediated inhibition of PA antibiotic resistance.
In future studies, we will characterize the role of these proteins in mediating the effects of Qi Gui Yin to overcome PA resistance to antibiotics. Moreover, investigation of whether the differential expression of proteins with unknown functions affects antibiotic resistance of PA is needed. In conclusion, based on the in vitro results of this study, Qi Gui Yin could effectively inhibit resistance of PA to IPM, and chlorogenic acid and astragaloside IV might be promising components for further characterization for potential therapeutic applications. Among the 76 differently expressed proteins, Qi Gui Yin could downregulate the expression of proteins involved in aerobic growth and bacterial biofilm formation. Further studies are needed to screen and characterize these differentially expressed proteins to clarify the mechanisms by which Qi Gui Yin inhibits antibiotic resistance of PA.
Footnotes
Authors' Contributions
J.D. contributed to study design, related experiments, data analysis, and writing of the article. X.G. and H.G. provided the tests. X.D., Y.L., and S.A. performed the animal experiments. Q.L. contributed to study design and revision of the article.
Acknowledgment
We thank Qun Ma for supplying experimental materials.
Disclosure Statement
All authors declare no potential conflicts.
Funding Information
This work was supported by the Beijing Natural Science Foundation (No. 7182071), the National Natural Science Foundation of China (No. 81503399), and the National Major Scientific and Technological Special Project for “Significant New Drugs Development” (No. 2013ZX09102026).