
Abstract
Objectives:
The limitations of treatment options in bloodstream infections caused by multidrug-resistant Acinetobacter baumannii (MDRAB) have been related to high morbidity and mortality. The aim of our present study was to determine antimicrobial susceptibility profiles, molecular resistance patterns, and biofilm properties of A. baumannii isolated from bloodstream infections.
Materials and Methods:
In the present study, a total of 44 A. baumannii bloodstream isolates were included. Antimicrobial susceptibility profiles and biofilm formation ability were assessed. The distribution of class D carbapenemases, ISAba1, ISAba1/blaOXA-23, blaNDM-1, mcr-1, and ompA was investigated by polymerase chain reaction (PCR). Arbitrarily primed-PCR (AP-PCR) was performed to evaluate clonal relationships.
Results:
A total of 32 isolates were MDRAB, whereas 6 isolates were also resistant to colistin without mcr-1 positivity. All isolates were harboring blaOXA-51 gene, whereas blaOXA-23 positivity was 63.6%. Fifty percent of the isolates had ISAba1. ISAba1 upstream of blaOXA-23 was determined in 18 isolates. None of the isolates were positive for blaNDM-1 gene. Majority of the strains were strong biofilm producers (86.8%). A total of 56.8% of the isolates were positive for ompA gene with no direct association with strong biofilm formation. However, blaOXA-51 + 23 genotype and trimethoprim–sulfamethoxazole resistance showed a significant relationship with biofilm formation. AP-PCR analysis revealed six distinct clusters of A. baumannii.
Conclusions:
Herein, majority of the A. baumannii blood isolates were characterized as blaOXA-51+OXA-23 carbapenemase genotype and were strong biofilm formers. None of the isolates were positive for blaNDM-1, which was promising. Resistant isolates were tended to form strong biofilms. Our results highlight the emergence of oxacillinase-producing MDRAB isolated from bloodstream with high biofilm formation ability.
Introduction
Acinetobacter baumannii is an important nosocomial pathogen, which is responsible for a wide range of infections with high morbidity and mortality rates. 1 In particular, bloodstream infections caused by A. baumannii have become a major clinical concern in clinical settings due to high rates of multidrug-resistant (MDR) phenotypes. 2 Overall mortality rates due to A. baumannii bloodstream infections have been reported between 34.0% and 43.4% in intensive care units. 3
The increasing rates of multidrug-resistant strains of A. baumannii (MDRAB) have been reported worldwide. Resistance in Acinetobacter spp. may occur by chromosomal mutations or mobile genetic elements. The resistance mechanism mainly due to permeability changes in outer membrane proteins, activation of efflux pumps, and hydrolysis by β-lactamases. 4
Class D carbapenemases are the main reason for increasing resistance rates in A. baumannii isolates, which mainly include acquired carbapenemases blaOXA-23, blaOXA-24, and blaOXA-58 as well as blaOXA-51, which is intrinsic to A. baumannii. 5 The oxacillinase genes are usually associated with insertion sequences (IS). 6 Class D carbapenemases, particularly blaOXA-23, are weak carbapenemases. However, mainly ISAba1 overexpresses the downstream located blaOXA-23 and enhances the hydrolyzing capacity of carbapenems. 6 Moreover, the availability of IS leads to transfer of resistance genes.7,8 Thus, the availability of IS in A. baumannii strains is also an important determinant in carbapenem-resistant A. baumannii (CRAB). Metallo-β-lactamases such as blaNDM-1 are also another major concern in MDRAB, however, in less frequency. 5
Due to the limited treatment options, CRAB has become a major concern. Colistin, tigecycline, and fosfomycin are the last resort treatment options in CRAB bacteremia. Due to the increasing use of colistin to treat CRAB, colistin resistance has been emerged. 5
Ability of biofilm formation on biotic and abiotic surfaces is one of the most important virulence factors of A. baumannii. Previous studies have reported an association between biofilm formation and antibiotic resistance in A. baumannii isolates. 9 Outer membrane protein A (OmpA) is the major surface protein of A. baumannii, which involves in biofilm formation, 10 adherence to epithelial cells, and inducing cytotoxicity, 11 and an important determinant of the mortality risk in bacteremia. 12
Genotypic and phenotypic resistance patterns and biofilm formation characteristics are the main factors effecting progression of A. baumannii infections.2,12 Accordingly, the aim of our present study was to determine antimicrobial susceptibility, distribution of molecular resistance genes and ompA, clonal relationship, and biofilm formation in A. baumannii bloodstream isolates.
Materials and Methods
Bacterial isolation
A total of 44 nonduplicated A. baumannii isolated from bloodstream infections in Hacettepe University Central Laboratory between September 2017 and November 2019 were included in the present study. Blood cultures were incubated in BD BACTEC Fx (Becton Dickinson, Maryland). Positive blood cultures were subjected to Gram staining and subcultured into 5% sheep blood agar, eosin methylene blue agar, and chocolate agar plates and incubated at 35°C for 24–28 hours in 5% CO2. Recovered isolates were identified by conventional methods and Matrix-assisted Laser Desorption/Ionization Time-Of-Flight Mass Spectrometry (MALDI-TOF MS) (Bruker Daltonics, Germany) as A. baumannii.
Antimicrobial susceptibility testing
Antibiotic susceptibility profiles were determined by the Kirby–Bauer disk diffusion susceptibility test for ampicillin–sulbactam, ceftazidime, imipenem, meropenem, gentamicin, ciprofloxacin, piperacillin–tazobactam, trimethoprim–sulfamethoxazole, amikacin, cefepime, and netilmicin. 13 MDRAB was defined as resistance to at least one antibiotic in three or more antimicrobial groups. 4 Imipenem and meropenem resistance was also confirmed by antimicrobial gradient test and classified as carbapenem resistant if they were resistant to one or more carbapenems tested. Colistin resistance was determined by broth microdilution method, and European Committee on Antimicrobial Susceptibility Testing (EUCAST) colistin resistance breakpoint of >2 mg/L was used. 14 Briefly, twofold serial dilutions of colistin sulfate (Sigma) were prepared ranging between 0.0625 and 64 μg/mL. Escherichia coli ATCC 25922 and mcr-1-positive E. coli NCTC 13846 were used as quality control strains.
Polymerase chain reaction amplification of antibiotic resistance genes
For bacterial DNA extraction, five to seven colonies were picked from fresh overnight cultures in Tryptic Soy Agar and suspended in 200 μL Tris-EDTA buffer. The tubes were then kept at 95°C for 15 minutes. After centrifugation at 10,000 rpm for 10 minutes, the supernatants containing sufficient concentration and good quality DNA were transferred into sterile micro tubes. Class D oxacillinases were determined by multiplex polymerase chain reaction (PCR). The PCR conditions and primers for amplification of carbapenemase-encoding genes including blaOXA-23-like, blaOXA-24-like, blaOXA-51-like, blaOXA-58-like, and blaNDM-1 were performed as previously described.15,16 Colistin-resistant strains were also tested for the presence of mcr-1 gene as reported by Rebelo et al., 17 by using mcr-1-carrying E. coli NCTC 13846 isolate as positive control. A frequent insertion sequence ISAba1 was also analyzed. 18 To detect the presence of ISAba1 located upstream of blaOXA-23, PCR amplification was carried out using the reverse primer of blaOXA-23 gene and forward primer of ISAba1 by using previously described primers and conditions.19,20 Since prior studies showed the importance of OmpA in biofilm formation 21 and its role in determining mortality in bacteremia, 12 ompA gene was also investigated. The specific primer pairs used in the study are presented in Table 1. Each amplicon was subjected to electrophoresis in 2% agarose gel. Amplified products were detected after staining with ethidium bromide (50 mg/L).
Primer Sequences Used in This Study
Biofilm formation assay
Biofilm formation degree of A. baumannii isolates was determined using a microplate assay as previously described with slight modifications. 22 Briefly, overnight A. baumannii cultures in Tryptic Soy Broth (TSB) were diluted to obtain 107 cfu/mL cell suspension. One hundred microliters of aliquots of cell suspension were added to each well. The plates were subsequently incubated at 37°C for 24 hours. The same amount of sterile TSB was added to top and bottom rows to avoid edge effects. After washing nonadherent cells, biofilms were stained with 0.1% crystal violet for 20 minutes and stained biofilms were solubilized with 33% acetic acid. Optical densities (OD) were measured at 570 nm. The OD570 values of uninoculated wells were used as a negative control. Biofilm assay was carried out in duplicates. Strains were classified as follows: non-biofilm producer (OD < ODc), weak biofilm producer (ODc < OD ≤2 × ODc), moderate biofilm producer (2 × ODc < OD <4 × ODc), and strong biofilm producer (4 × ODc < OD). 9 Biofilm-producing strain of Pseudomonas aeruginosa PA01 was used as positive control.
DNA fingerprinting by arbitrarily primed PCR
Clonal relatedness of the isolates was determined using arbitrarily primed PCR (AP-PCR). Since AP-PCR does not require any particular sequence, 23 we used M13 universal primers for random amplification (5′-GTAAAACGACGGCCAGTG-3′ and 5′-CAGGAAACAGCTATGACCATG-3′). Reaction mixture was prepared in 25 μL final volume containing 10 × PCR buffer, 2.5 mM MgCl2, 2.5 mM dNTP mix, 2.5 mmol of each primer, and 2.5 U Taq polymerase (SibEnzyme). Amplification reactions include an initial denaturation (4 minutes at 95°C), followed by 8 cycles (94°C for 30 seconds, 37°C for 1 minute, 72°C for 2 minutes) and 35 cycles of denaturation (30 seconds at 94°C), annealing (1 minute at 60°C), and extension (2 minutes at 72°C), with a single final extension of 5 minutes at 72°C. Each amplicon was subjected to electrophoresis in 1.8% agarose gel. Amplified products were detected after staining with ethidium bromide (50 mg/L). The DNA fragments were visualized, and gel images were analyzed with the GelJ software v.2.0. 24 The dendrogram was constructed by unweighted pair-group method with arithmetic mean. Dice coefficient with a 2% band tolerance was used. Isolates were considered within the same clone if they had higher than 90% similarity.
Statistical analysis
Data are expressed as percentage and mean ± standard deviation of the mean. Comparison of the mean OD570 values for biofilm formation between the groups was performed with the nonparametric Mann–Whitney U test or Kruskal–Wallis test with Dunn's multiple comparison testing, where appropriate. Correlation analysis was performed using the Spearman rank correlation since the variables are not normally distributed. p-Values of <0.05 was considered as significant.
Results
Antimicrobial susceptibility of the isolates
All bloodstream isolates included in the study were identified as A. baumannii by MALDI-TOF MS. A total of 32 isolates were defined as MDRAB according to antimicrobial susceptibility profiles (Table 2). Of the 44 bloodstream isolates, resistance rates for tested antibiotics were as follows: 72.7% for ampicillin–sulbactam, imipenem, meropenem, ciprofloxacin, piperacillin–tazobactam, cefepime, netilmicin; 75.0% for ceftazidime; 70.5% for gentamicin; 81.8% for trimethoprim–sulfamethoxazole, and 68.2% for amikacin.
Antimicrobial Susceptibility Patterns of Acinetobacter baumannii Bloodstream Isolates (Number of Isolates)
Antimicrobial susceptibility testing was determined by Phoenix™ 100 (Becton Dickinson). Imipenem and meropenem resistance was further confirmed by antimicrobial gradient test. Colistin resistance was determined by broth microdilution method.
NA, not applicable; TMP/SMX, trimethoprim–sulfamethoxazole.
According to broth microdilution results, six isolates were determined as colistin resistant (13.6%). All colistin-resistant isolates were resistant to all tested antibiotics. None of the colistin-resistant isolates were harboring mcr-1 gene.
Molecular detection of antibiotic resistance genes and ompA
PCR analysis for class D β-lactamases revealed that all isolates were harboring intrinsic blaOXA-51 gene as expected. 25 None of the isolates were positive for blaOXA-58 and blaOXA-24 genes. blaOXA-23 gene was found in 28 of the 44 A. baumannii bloodstream isolates (63.6%). However, when we only consider carbapenem-resistant isolates, 87.5% of them were positive for blaOXA-23 gene (Supplementary Fig. S1).
Insertion sequence ISAba1 gene was present in 22 isolates (50.0%). Thus, we further ask the question if blaOXA-23 was located in the downstream of ISAba1 or not. By using the forward primer for ISAba1 and reverse primer for blaOXA-23, ISAba1 upstream of blaOXA-23 was determined in 18 isolates of 28 blaOXA-23-positive isolates (64.3%). All blaOXA-23-positive strains and ISAba1/blaOXA-23-positive strains were resistant to carbapenems.
None of the isolates were harboring mobile plasmid-mediated colistin resistance gene mcr-1 and blaNDM-1. ompA gene was determined in 25 isolates (56.8%). Among ompA-positive strains, 21 of them were MDRAB.
Biofilm formation
According to the microtiter plate biofilm assay, among the 44 tested isolates, only 3 isolates (6.8%) were not biofilm producers (OD570: 0.10 ± 0.02), 5 isolates (11.4%) were moderate biofilm formers (OD570: 0.34 ± 0.07), and 36 isolates (86.8%) were strong biofilm formers (OD570: 1.44 ± 0.62) (Supplementary Fig. S2). Among 32 MDRAB, 24 were strong and 5 were moderate biofilm formers, whereas 3 of them were not biofilm formers. All 12 non-MDRAB isolates were strong biofilm formers.
Next, we aimed to determine the relationship between the biofilm formation ability and antimicrobial susceptibility to representative antibiotic agents covering penicillin combination, carbapenem, cephalosporin, sulfonamide, aminoglycoside, fluoroquinolone, and polymyxin groups (Fig. 1A–L). Due to the substantial differences in sample size, only one isolate was intermediate resistant to piperacillin–tazobactam (Fig. 1B) and one isolate was susceptible to ceftazidime (Fig. 1E), we exclude these groups form group comparisons. For all tested antibiotics, resistant isolates tended to form strong biofilms according to their OD570 values, except for colistin. The isolates resistant to trimethoprim–sulfamethoxazole showed significantly higher OD570 values for the biofilm formation ability when compared with susceptible isolates (p = 0.01; Fig. 1G). We also conducted a correlation analysis between biofilm formation ability and antimicrobial susceptibility profiles. There was a significant positive correlation between the biofilm formation ability and trimethoprim–sulfamethoxazole resistance (r = 0.38, p = 0.01). Resistance to all tested antimicrobials was positively correlated with biofilm formation, except colistin (r = −0.07, p = 0.67), however, not significant.
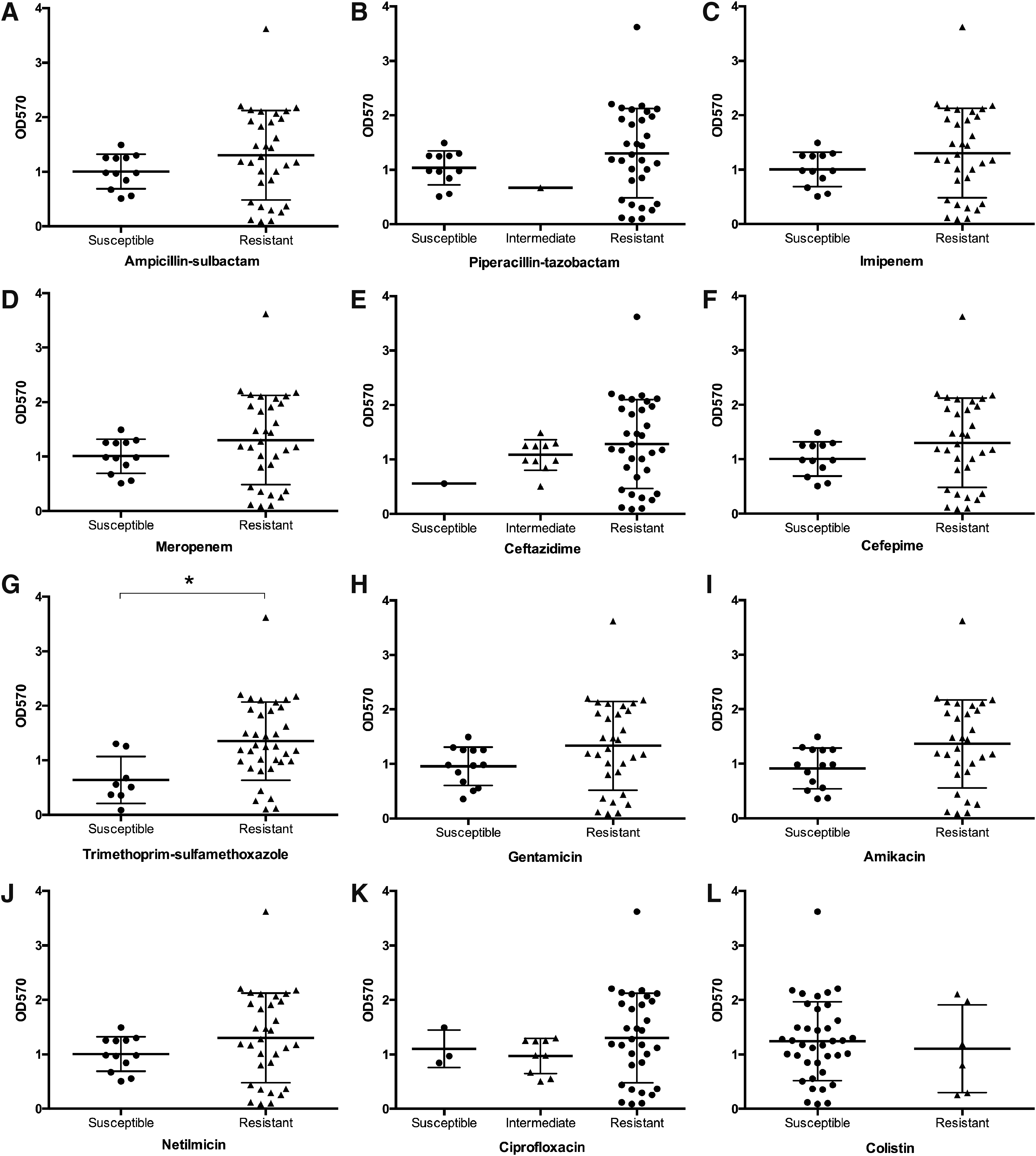
Comparison of biofilm formation capability and different antibiotic resistance phenotypes
Among 25 ompA-positive strains, 20 of them (80.0%) were strong biofilm formers, whereas 4 of them were moderate and 1 of them was none biofilm former. Moreover, 17 of 19 ompA-negative isolates were strong biofilm formers, whereas 2 of them were none biofilm formers (Table 3). There was no significant relationship between ompA positivity and OD570 values indicating the biofilm formation ability (p = 0.79; Fig. 2A).

Correlation of ompA
Distribution of the Isolates According to Biofilm Formation Ability and Specific Gene Positivity
Since our results highlight the dissemination of blaOXA-23, we also investigated the relationship between blaOXA-23 and biofilm formation ability. blaOXA-23-positive isolates had significantly higher biofilm formation ability according to OD570 values (p = 0.01; Fig. 2B). However, there was no significant relationship between ISAba1/blaOXA-23 strains and biofilm formation (p = 0.51; Fig. 2C). Distribution of the isolates according to the biofilm formation ability and presence or absence of a specific gene is presented in Table 3.
Clonal diversity
Dendrogram was constructed according to AP-PCR patterns (Supplementary Fig. S3). The genotypic and phenotypic characteristics of the isolates are presented in the dendrogram in Fig. 3. A total of 44 bloodstream isolates revealed 11 different clones with 90% similarity cutoff value. At 100% similarity level, there were 22 different clones. We found that 39 isolates were clonally related, which were gathered in 6 clusters with a tolerance of 2% and a cutoff value of 90% similarity. The numbers of the isolates in each cluster were ranging between 2 and 17 (cluster A, n = 2; cluster B, n = 4; cluster C, n = 17; cluster D, n = 6; cluster E, n = 3; cluster F, n = 7). All four isolates in cluster B had 100% similarity. The largest cluster was cluster C. In cluster F, all the isolates were MDR, blaOXA-51 and blaOXA-23 positive, harboring ISAba1, and all of them were located upstream of blaOXA-23. Moreover, cluster F isolates were all ompA positive, and all of them were strong biofilm formers except one isolate. The isolation dates for cluster F was within 1 month. The other clusters did not reveal a specific genotype or phenotype. All six colistin-resistant isolates were within a cluster: three of them were in cluster C, two of them were in cluster F, and one of them was in cluster D.
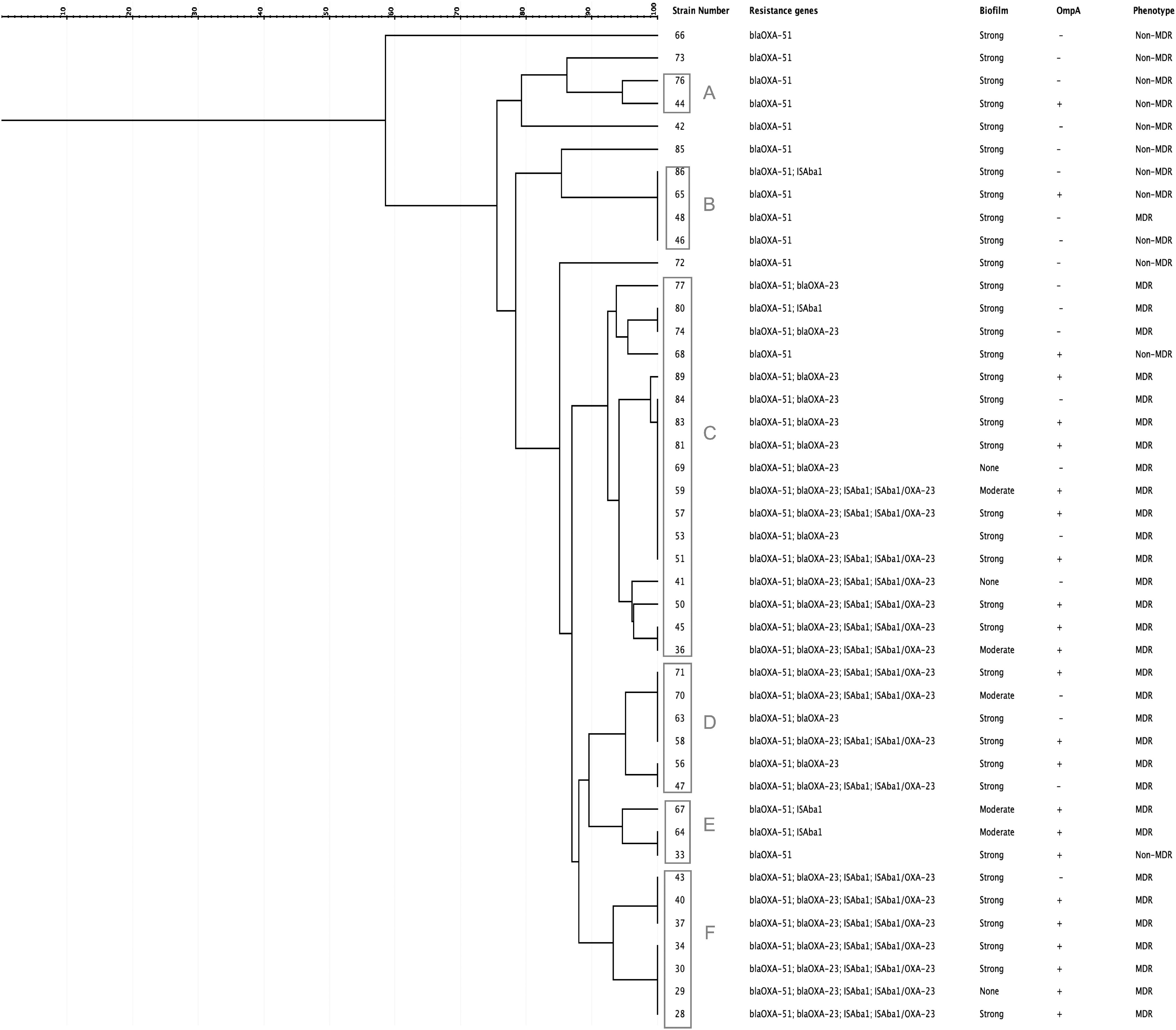
Dendrogram of the Acinetobacter baumannii isolates. The clusters were named with uppercase letters (A–F). Genotypic and phenotypic characteristics of the isolates were presented for individual strains.
Discussion
Higher prevalence rates of MDRAB have become a major problem worldwide particularly in intensive care units. 2 Therefore, it is important to define genotypic and phenotypic characteristics of the MDRAB isolates as a part of infection control measures. In addition to the antimicrobial susceptibility profiles, biofilm formation ability of a clinical isolate is also of importance, because biofilm formation is one of the main determinants of virulence affecting treatment options. 9 Majority of the studies on genotypic and phenotypic characterization of A. baumannii clinical isolates were involving isolates from various clinical samples.8,21,26,27 In this present prospective single-center study, we only included bloodstream isolates of A. baumannii. Herein, we defined 32 of the 44 bloodstream isolates as MDRAB, and 6 isolates were also resistant to colistin. Majority of the isolates were strong biofilm formers. blaOXA-51 and blaOXA-23 were the major responsible oxacillinases in our study population.
Colistin (polymyxin E) has been considered as the last resort treatment in infections caused by MDR gram-negative bacteria due to its neurotoxicity and nephrotoxicity. 28 However, it has become an increasingly important treatment option for carbapenem-resistant gram-negative bacteria. 28 Colistin resistance rates in A. baumannii were not as high as observed in Klebsiella pneumoniae isolates in Turkey 29 and Europe. 30 We found a colistin resistance rate of 13.6% by microdilution method in our present study. In a recent study from Hungary, colistin resistance rate of 2.6% for A. baumannii blood culture isolates was reported. 31 In another recent study on A. baumannii bloodstream infections from a Greek university hospital, colistin resistance rate was reported as high as 42%. 32 None of the isolates were harboring plasmid-mediated colistin resistance in our study. Therefore, the resistance to colistin might be mainly due to chromosomal mutations in PmrA/PmrB two-component system 33 or other less frequent mechanisms including efflux pumps 34 and complete loss of lipopolysaccharide. 35
Oxacillinases are the major enzymes involved in antibiotic resistance in A. baumannii strains. 36 In our present study, all A. baumannii isolates had intrinsic blaOXA-51 gene, whereas 63.6% of them were harboring blaOXA-23 gene and none of them were positive for blaOXA-58 and blaOXA-24. These findings are also similar with other studies, which reported the most prevalent oxacillinases in A. baumannii as blaOXA-51 and blaOXA-23 worldwide with various prevalence rates between geographical regions.26,37
Similar to our previous findings, other recent studies from Turkey also reported that all clinical A. baumannii isolates were harboring blaOXA-51 gene. However, blaOXA-23 prevalence was reported between 94% and 97% in those studies.27,38 We also did not determine blaNDM-1 in any of the isolates. This finding is consistent with another recent study from our country. 38 Moreover, we also showed the significant relationship between blaOXA-23 genotype and biofilm formation, which is consistent with a previous study on MDRAB isolates from various clinical samples. 39
The dissemination of blaOXA-23-producing MDRAB isolates has been reported worldwide, including Turkey.26,27 It has been known that the expression of oxacillinases might be increased by ISAba1 sequence upstream of blaOXA-23 gene. 8 We found that ISAba1 was located upstream of the blaOXA-23 gene in 18 of 28 blaOXA-23-positive isolates. Moreover, all blaOXA-23-positive isolates with or without upstream-located ISAba1 showed resistance to carbapenems. It should also be noted that OXA-23-positive isolates without upstream-located ISAba1 could be susceptible to carbapenems indicating the nonfunctional expression of oxacillinase. 40
Higher prevalence rates for ISAba1/blaOXA-23 have been reported from a recent study in which majority of the A. baumannii isolates were from respiratory tract. 8 Since majority of our isolates were carbapenem resistant (32/44), the relatively low prevalence of blaOXA-23 and ISAba1/blaOXA-23 is surprising. However, this can be explained by coexistence of other resistance mechanisms in these strains.
Similar to our present findings, majority of the studies also reported 100% prevalence rates for chromosomally encoded blaOXA-51 in both Turkey 27 and worldwide. 37 These findings support the previous studies that blaOXA-51 gene can be used as a supplementary tool for identification of A. baumannii. 25
To prevent the spread of A. baumannii in clinical settings, it is important to determine its origins and transmission routes. 41 We also performed AP-PCR analysis to determine clonally related isolates. It was noteworthy that all the isolates in one of the dominant cluster (cluster F) were carrying blaOXA-51 and blaOXA-23 with an upstream-located ISAba1. Moreover, all the isolates in this cluster were MDR, and two of them were colistin resistant. Since the isolation dates were closer for that cluster, it might be considered as a small clonal spread within the intensive care unit.
Bacteremia caused by A. baumannii is mostly caused by intravascular and respiratory tract catheters. 1 Thus, it is important to determine biofilm characteristics of A. baumannii isolates since the biofilm formation ability plays an important role in the host–bacteria interaction and infection outcome. 42 As expected, majority of the isolates were biofilm formers in our study. In addition, we determined a high rate of strong biofilm formation (86.8%) when compared with other reported rates in the literature for A. baumannii.43,44 However, it should be noted that we used a static assay for biofilm formation. Continuous flow systems might result in less biofilm formation when compared with static models, as previously reported. 45
The relationship between biofilm formation capability and antibiotic resistance phenotypes remains controversial. 42 While some of the studies demonstrated that the biofilm formation ability was positively correlated with resistance,9,46 some other studies showed that MDR strains tended to form weak biofilms.47,48 In our present study, we investigated the association of biofilm formation capability with individual antibiotic phenotypes. We determined a significant positive correlation between trimethoprim–sulfamethoxazole resistance and biofilm formation ability. For other antibiotics, resistant isolates were tended to form stronger biofilms except for colistin. This might be due to substantial differences in number of resistant and susceptible isolates since there was only six colistin-resistant isolates. Our results are consistent with those of Yang et al., 44 as both studies showed the tendency for stronger biofilm formation in resistant isolates and a significant positive correlation between the biofilm formation ability and resistance to trimethoprim–sulfamethoxazole.
While our antimicrobial susceptibility results were only covering planktonic cells, it should be noted that the minimum biofilm eradication concentration values might be much higher than the minimum inhibitory concentrations determined in our study, as previously reported. 49 Da Cunda et al. 50 investigated a colistin- and gentamicin-susceptible isolate of A. baumannii and reported that none of the antibiotics were sufficient in eradication of the biofilm stage.
OmpA is a prominent surface protein in A. baumannii, which is involved in drug resistance, adhesion to epithelial cells and abiotic surfaces, and biofilm formation. 10 It has been previously shown that OmpA is a risk factor for the mortality rate in bacteremia caused by A. baumannii. 12 Considering its importance in bloodstream isolates, we also investigated ompA gene positivity rates, which was relatively lower (56.8%) when compared with the reported rates in the literature (91–100%).21,44 Moreover, the biofilm formation ability was not significantly different between ompA-positive and ompA-negative isolates. There have been conflicting results and no clear evidence to ascertain the essential role of OmpA in biofilm formation.10,44,51 In a study in which 154 A. baumannii strains were tested for the biofilm formation ability, it was found that 45.4% were strong biofilm formers. Moreover, ompA gene was present in 91.6% of the isolates they tested and they found that ompA-positive strains tended to form strong biofilm than the negative isolates. 44 However, these findings are mostly from A. baumannii isolated from various clinical samples. The role of OmpA might be dependent on the clinical origin of the strain. Besides our majority of the isolates were strong biofilm formers, evidence from the previous literature 51 and our present findings suggests that OmpA may not have an essential role in biofilm formation in blood isolates.
In our present study, we characterized the A. baumannii bloodstream isolates for their antimicrobial susceptibility, major resistance genotypes, and biofilm properties along with OmpA distribution. Moreover, we also showed the significant correlation of trimethoprim–sulfamethoxazole resistance and biofilm formation. Majority of the studies on this field were focusing on various sample sites for A. baumannii characterization. Thus, our present findings may contribute to the current literature on bloodstream infections of A. baumannii.
We acknowledge the following limitations of our study. First, our sample size was relatively small, and only 72.7% of them were MDRAB. However, bloodstream isolates are rare when compared with other clinical sites and we included nonduplicated A. baumannii isolates. Second, we did not screened for all possible resistance and biofilm-related genes, such as efflux pump genes adeABC and adeJK, or bapA for the biofilm formation ability. None of the isolates were harboring mcr-1; however, we did not confirm the PmrA/PmrB mutations, which was likely to be the main colistin resistance mechanism in our A. baumannii isolates.
Conclusions
In conclusion, our present findings highlight the dissemination of blaOXA-23 gene in A. baumannii bloodstream isolates. The emergence of oxacillinase-producing MDRAB bloodstream isolates, with high virulence characteristics such as strong biofilm formation ability, underscores the importance of screening and early detection of these strains. Moreover, most of the oxacillinase genes are located in highly mobile genetic elements. The screening for oxacillinases and metallo-β-lactamases is of importance to control dissemination of these strains as a part of nosocomial infection control measures.
Footnotes
Acknowledgment
We would like to acknowledge Public Health Institution of Turkey, National Antimicrobial Resistance Laboratory, for providing E. coli NCTC 13846.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.