
Abstract
Resistance to third-generation cephalosporins (3GC) in Escherichia coli has been reported worldwide from humans and animals, but the situation in Cuba is still poorly understood. This study aimed to gain new insights into the phenotypic and genotypic characteristics of third-generation cephalosporin-resistant (3GC-R) E. coli isolated from pigs in Cuba. Rectal swabs from 215 healthy pigs were taken from different municipalities in the western region of Cuba and spread on MacConkey agar supplemented with cefotaxime and ceftazidime. Ninety-six isolates were identified as 3GC-R E. coli and 87.5% of them were resistant to at least three antibiotic classes as determined by the measurement of the minimum inhibitory concentration (MIC) of 14 antibiotics. Twenty-seven different isolates were selected for Illumina next-generation sequencing, and subsequent in silico analysis was performed for the detection of antibiotic resistance and virulence genes, plasmid incompatibility (Inc) groups, multilocus sequence typing (MLST), and core genome MLST (cgMLST). The sequenced isolates contained extended-spectrum β-lactamase genes blaCTX-M-32 (n = 17), blaCTX-M-15 (n = 5), and blaCTX-M-55 (n = 4) as well as with pAmpC gene blaCMY-2 (n = 2). They also harbored genes for resistance to other clinically important classes of antibiotics, as well as several diverse virulence factors. The 3GC-R E. coli were genetically highly diverse, belonging to 16 different sequence types. IncX1 was the most frequent Inc group. The presence of 3GC-R E. coli in pigs from Cuba containing several different antibiotic resistance mechanisms emphasizes the need for surveillance programs and the establishment of strategies for the prudent use of antibiotics in food-producing animals.
Introduction
Antimicrobial-resistant bacteria and their spread represent one of the biggest challenges that modern medicine and animal production are facing today. 1 Overuse and misuse of antimicrobials both in humans and animals have led to both the selection of antimicrobial-resistant bacteria and the acquisition of several mechanisms of resistance.2,3
Resistance to clinically important antibiotics such as cephalosporins is commonly observed in Escherichia coli associated with hospital- and community-acquired infections around the world. 4 In addition, third-generation cephalosporin-resistant (3GC-R) E. coli has been reported worldwide in food-producing animals, which may contaminate meat at slaughterhouses, posing the risk that they reach the human gut through the food chain where they can transfer their resistance genes to human pathogens.5–9 Therefore, the survey of 3GC resistance in E. coli has been proposed by the World Health Organization (WHO) to be part of the antibiotic resistance surveillance program in all countries. 10 Cephalosporin resistance in E. coli is usually mediated by extended-spectrum beta-lactamases (ESBLs, e.g., blaCTX-M) generally carried on mobile genetic elements such as plasmids and transposons,11,12 and by AmpC beta-lactamases located on plasmids (pAmpC) such as blaCMY genes or by derepression of the chromosomal AmpC. 13 3GC-R E. coli are also frequently resistant to other clinically important classes of antibiotics such as fluoroquinolones and aminoglycosides.14–16
Cuba is facing challenges with multidrug-resistant organisms,17–20 including ESBL-producing E. coli in both humans and animals.20–22 A pilot study conducted in 2016–2017 in pig husbandry in the Matanzas province revealed the presence of ESBL-producing Enterobacteriaceae. 21 However, the phylogenetic, virulence, and antibiotic resistance characteristics have never been investigated in any 3GC-R E. coli isolates from animals in Cuba. We, therefore, screened pigs for the presence of 3GC-R E. coli and selected genetically diverse 3GC-R E. coli based on their rep-PCR profiles, resistance phenotype, origin and date of collection for next-generation sequencing (NGS), and subsequent genetic characterization using bioinformatic tools.
Materials and Methods
Samples collection
The samples were collected from 215 healthy pigs during the rainy and hot season in July 2016, September 2016 and 2017, and May 2018 (average temperature ∼30°C) in three farms and two slaughterhouses of different municipalities of the Western region of Cuba: pigs from one farm in San José de las Lajas (SJL/F) (n = 52), one farm in Calimete (CLM/F) (n = 42), one farm in Matanzas (MAT/F) (n = 42), and from one slaughterhouse in Matanzas (MAT/S) (n = 10) and one slaughterhouse in La Lisa (LLA/S) (n = 69). Pigs slaughtered in Matanzas originated from the farm of Matanzas (MAT/S/F) (n = 10) and pigs slaughtered in La Lisa were from the farms of La Lisa (LLA/S/F) (n = 57) and Caimito (LLA/S-CAI/F) (n = 12). Only the samples from Matanzas were taken once at the farm and once at the slaughterhouse, but at different time. The samples in the abattoirs were taken before slaughter and from animals with different origins and kept in different waiting areas. All the samples of a single facility were taken the same day and no pigs were sampled twice. This study was part of project 9190 entitled “Capacity building for the control of antimicrobial resistance” conducted within the framework of the National Program of Animal and Plant Health at the National Centre for Animal and Plant Health (CENSA). The Committee on Animal Research and Ethics at CENSA approved the experimental design, and the project including sampling was authorized by the Directorate of Animal Health of the Ministry of Agriculture in Cuba. All farm owners gave informed consent to sample collection. Export of the samples from Cuba to Switzerland was authorized under the Zoosanitary Export Certification number R.S.9192018. Animals sampled at the abattoirs consisted of fattening pigs, and those sampled at the farms consisted of newborn and postweaning piglets; sow pigs were also sampled in the farm of Calimete. The breed of the pigs consisted of hybrids originating from Yorkshire, Landrace, and/or Yorkland (maternal breeds) and Duroc, CC21 and/or L35 (paternal breeds). The sample size was based on the number of pigs present in each facility (either at the farms or at the slaughterhouses) at the time of sampling, considering a prevalence of 20% with a confidence level of 95% and 10% estimated error based on previous studies. 21 The rectal swabs were collected in 1 mL of Cary–Blair medium. 23 All swabs were transported from each facility to the Laboratory of Animal Bacteriology at CENSA in a cooler containing ice packs and were processed the same day.
Isolation and identification
Screening for 3GC-R E. coli isolates was performed using two selective plates of MacConkey agar (BioCen, BioCubaFarma, Cuba), of which one plate contained 4 μg/mL of cefotaxime (Sigma-Aldrich; Merck®, Missouri, USA) and the other 8 μg/mL of ceftazidime (Sigma-Aldrich; Merck®). Plates were incubated in aerobic conditions at 37°C for 18–24 hr. Lactose-positive colonies were selected and streaked three times on selective MacConkey plates to obtain pure cultures. E. coli isolates were initially identified by biochemical tests using API-20E multisubstrate gallery (bioMérieux, Lyon, France). Identification was confirmed by matrix-assisted laser desorption/ionization time of flight (Microflex LT; Bruker Daltonics GmbH, Bremen, Germany).
Minimal inhibitory concentration testing
The minimal inhibitory concentrations (MICs) of 14 antibiotics were determined for all isolates by the microdilution method in cation-adjusted Mueller–Hinton broth using EUVSEC Sensititre plates (Thermo Fisher Scientific, Inc., Waltham, USA). MICs were interpreted using the clinical recommendations of the European Committee on Antimicrobial Susceptibility Testing 24 as indicator for resistance for ampicillin (>8 μg/mL), ceftazidime (>4 μg/mL), cefotaxime (>2 μg/mL), chloramphenicol (>8 μg/mL), ciprofloxacin (>0.5 μg/mL), colistin (>2 μg/mL), gentamicin (>4 μg/mL), meropenem (>8 μg/mL), tigecycline (>0.5 μg/mL), and trimethoprim (>4 μg/mL), and using the Clinical and Laboratory Standards Institute guidelines 25 for the interpretation of MIC values for nalidixic acid (≥16 μg/mL), sulfamethoxazole (≥512 μg/mL), and tetracycline (≥16 μg/mL).
Genetic relatedness of E. coli
Repetitive element palindromic PCR (rep-PCR) was performed with all E. coli isolates to select those for whole-genome sequencing (WGS) based on their fingerprinting profile. DNA for rep-PCR was extracted from a loop-full of bacterial cells suspended in Chelex 100 buffer [100 mM Tris pH8, 0.5 nM EDTA, 10% (m/v) Chelex 100] and incubated at 58°C for 60 min followed by a 15-min denaturation step at 95°C. PCR was performed using 5 × HOT FIREPol® Master Mix Ready to Load (Solis BioDyne, Tartu, Estonia) following previously described conditions. 26 The rep-PCR products were visualized in 2% agarose gels stained with RedSafe (Solis BioDyne) and 100 bp DNA ladder (Solis BioDyne) was used as a reference. Gels were visualized by Gel Doc™ XR+ (Bio-Rad Laboratories, California, United States) using Image Lab™ software v5.1 and rep-PCR fingerprints patterns were analyzed using Bionumerics v7.6 software (Applied Maths NV, Sint-Martens-Latem, Belgium). An unweighted pair-group dendrogram was obtained applying the Pearson correlation similarity coefficient (1% of tolerance and optimization of 0.5%). Isolates showing rep-profile identity ≥90% were considered as genetically related. 27
WGS and in silico analysis
Isolates for WGS were selected according to the following criteria: rep-PCR profiles, resistance phenotypes, and origin of the isolates. One to six isolates were selected from each sampling site namely SJL/F (n = 6), CML/F (n = 6), MAT/F (n = 5), LLA/S/F (n = 5), LLA/S-CAI/F (n = 4), and MAT/S/F (n = 1). DNA was extracted from liquid cultures of E. coli obtained in Luria-Bertani broth after 18–24-hr incubation at 37°C with agitation using DNeasy Blood and Tissue (Qiagen, Hilden, Germany). WGS was performed using the Illumina NovaSeq6000 system (2 × 150 bp paired ends) (Eurofins, Constance, Germany). Short reads were trimmed using Trimmomatic v0.36 28 to remove adaptor residues and low-quality (Q ≤ 20) ends. Assembled genomes were generated using the SPAdes (v3.12) assembler 29 and were analyzed in silico for known antibiotic resistance genes and mutations with known phenotype (ResFinder v3.1), 30 virulence genes (VirulenceFinder v2.0), 31 sequence type (ST) based on Achtman E. coli multilocus sequence typing (MLST) scheme (MLST v2.0) 32 and incompatibility groups (Inc types) of plasmids (PlasmidFinder v2.0) 33 using default parameters of the web-based bioinformatics tools from the Center for Genomic Epidemiology. Contigs >10 kb and containing Inc groups were compared with sequences from the GenBank using BLASTN at the National Center for Biotechnology Information (www.ncbi.nlm.nih.gov). Similarity with known plasmid sequences was defined when query coverage and identity were ≥90%.
Core genome MLST analysis
The genome-wide gene-by-gene comparison of the sequenced isolates was carried out by core genome MLST (cgMLST) using the Stable cgMLST scheme for E. coli (2,513 genes) and SeqSphere+ software v5.1 (Ridom GmbH, Münster, Germany). The neighbor-joining tree based on phylogenetic distances was generated using SeqSphere+ software and clusters were determined considering a distance of 10 different genes.
Results
Ninety-six of the 215 pig sampled were found to be positive for 3GC-R E. coli. The 3GC-R E. coli isolates were detected in pigs from each investigated farms and slaughterhouses with 65 of 136 pigs being positive in the farms (25 of 52 in San José de las Lajas, 28 of 42 in Calimete, 12 of 42 in Matanzas), and 31 of 79 in the slaughterhouses (3 of 10 in Matanzas and 28 of 69 in La Lisa of which 8 of 12 were from the farm of Caimito and 20 of 57 were from the farm of La Lisa).
The 96 3GC-R E. coli isolates were resistant to ampicillin and cefotaxime. The isolates also exhibited resistance to others antibiotic such as tetracycline (94.8%), sulfamethoxazole (75%), trimethoprim (69.8%), ceftazidime (64.6%), chloramphenicol (44.8%), nalidixic acid (51%), ciprofloxacin (31.3%), gentamicin (27.1%), and tigecycline (4.2%). All isolates were susceptible to colistin and meropenem. Resistance to at least three antibiotic classes was detected in 87.5% of the isolates (Supplementary Fig. S1).
Based on profiles obtained by rep-PCR, resistance phenotype and origin (Supplementary Fig. S1), 27 diverse isolates were selected for WGS. The 27 sequenced isolates belonged to 16 sequences types (STs) (Fig. 1), namely ST162 (n = 4), ST2599 (n = 3), ST9362 (n = 3), ST1695 (n = 3), ST48 (n = 2) and ST648 (n = 2), ST58 (n = 1), ST165 (n = 1), ST205 (n = 1), ST457 (n = 1), ST641 (n = 1), ST710 (n = 1), ST772 (n = 1), ST1397 (n = 1), ST9360 (n = 1), and ST9361 (n = 1). ST9360, ST9361, and ST9362 represented new STs. The phylogenetic analysis performed by cgMLST confirmed that the E. coli strains from different ST were genetically diverse, whereas those of the same ST were closely related, gathering into the same cluster (Fig. 1). Strains belonging to the same ST differed from each other by either different resistance profiles (ST162, ST648, ST1695, and ST2599) (Fig. 1) or different sampling locations (ST162 in farms from Calimete and San José de Las Lajas; ST1695 in farm and slaughterhouse from Matanzas). Strains of a same ST (ST2599 and ST9362) were also found at the slaughterhouse of La Lisa in pigs originating from different farms (La Lisa and Caimito) (Fig. 1).
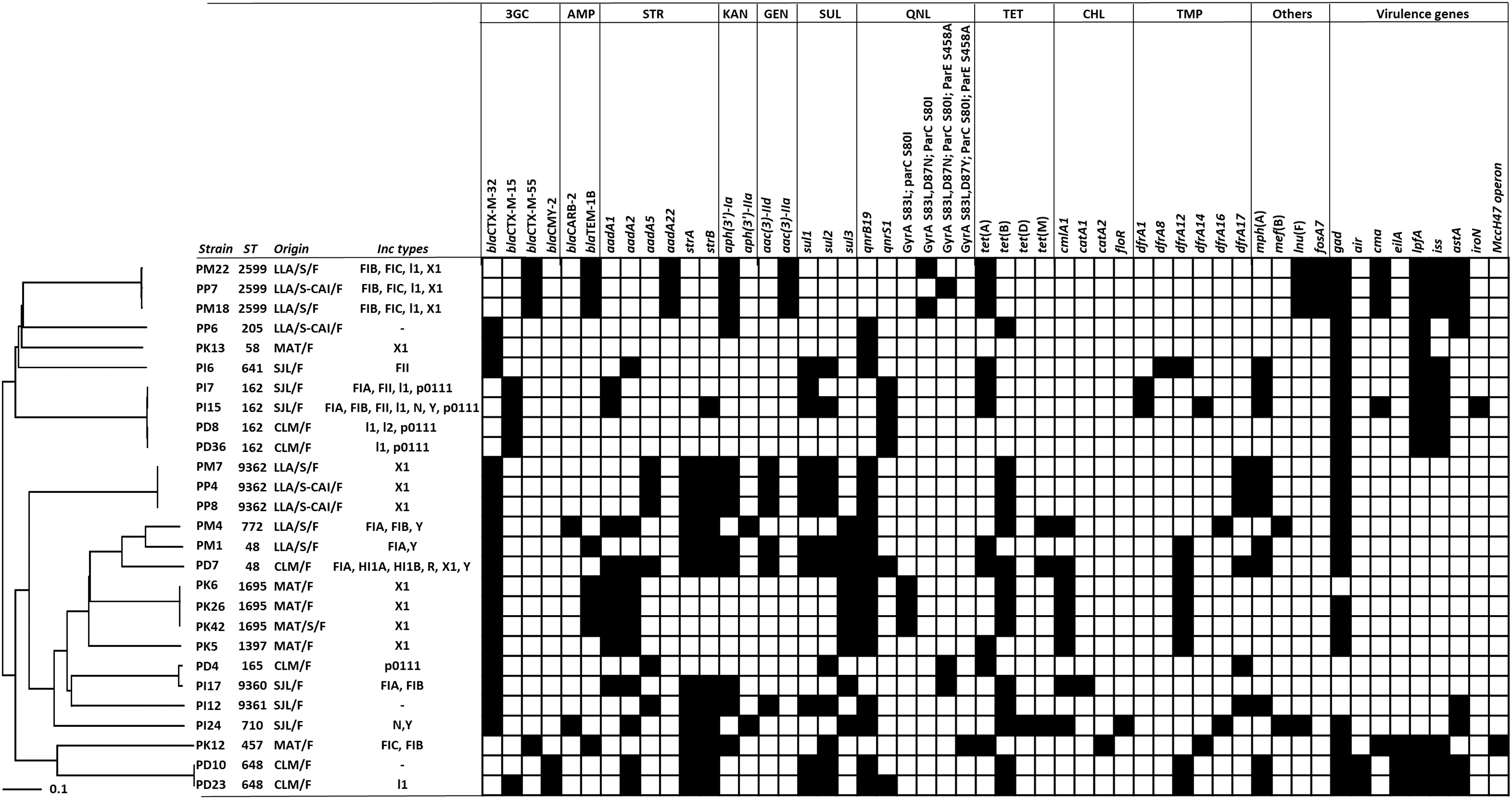
Phylogenetic neighbor-joining tree of 27 sequenced 3GC-R-Escherichia coli isolates from healthy pigs in Cuba based on stable cgMLST, including 2513 genes, and distribution of the STs (n = 12), Inc types (n = 13), antibiotic resistance (n = 39), and virulence genes (n = 9). The tree was generated with SeqSphere+ software v5.1 (Ridom GmbH, Münster, Germany). The genes were detected using web-based bioinformatics tools available at the Center of Genomic Epidemiology. Antibiotics: 3GC, third-generation cephalosporins; AMP, ampicillin; CHL, chloramphenicol; GEN, gentamicin; KAN, kanamycin; QNL, quinolones; TET, tetracyclines; TMP, trimethoprim; STR, streptomycin; SUL, sulfonamides; others, macrolides, lincosamides, and fosfomycin. Sampling places: CLM/F, farm of Calimete; LLA/S/F, slaughterhouse of La Lisa from pigs raised in the farm of La Lisa; MAT/F, farm of Matanzas; MAT/S, slaughterhouse of Matanzas from pigs raised in the farm of Matanzas; SJL/F, farm of San José de Las Lajas; LLA/S-CAI/F, slaughterhouse of La Lisa from pigs raised in the farm of Caimito. 3GCR, third-generation cephalosporin-resistant; cgMLST, core genome multilocus sequence typing; Inc, incompatibility.
Resistance to cephalosporins in the 27 sequenced 3GC-R E. coli isolates was mainly conferred by ESBLs of the CTX-M family (Supplementary Table S1). The most frequent ESBL genes were blaCTX-M-32 (n = 17) followed by blaCTX-M-15 (n = 5) and blaCTX-M-55 (n = 4). The blaCTX-M-32 gene was detected in E. coli of 12 different STs, and the blaCTX-M-15 and blaCTX-M-55 genes were found in 2 STs each (ST162, ST648 and ST457, ST2599). In contrast, the pAmpC gene blaCMY-2 was only found in two isolates belonging to ST648, one of them also containing blaCTX-M-15 (Fig. 1). Distribution of the other resistance genes among the 3GC-R E. coli isolates is presented in Fig. 1.
Beyond the 3GC resistance genes, the isolates also contained genes conferring resistance to gentamicin (n = 9), chloramphenicol (n = 9), ampicillin (n = 10), kanamycin (n = 14), trimethoprim (n = 19), sulfamethoxazole (n = 20), streptomycin (n = 23), and tetracycline (n = 24) (Supplementary Table S1 and Fig. 1). Amino acid substitutions in the quinolone resistance-determining regions (QRDR) of GyrA (S83L, D87N, D87Y), ParC (S80I), and ParE (S458A) were found in the nine isolates exhibiting high-level resistance to ciprofloxacin and nalidixic acid (Supplementary Table S1). Decreased susceptibility to both ciprofloxacin and nalidixic acid was observed in 13 isolates containing qnrB19 and qnrS1 genes without mutations in the QRDR of the topoisomerases. Four isolates containing either qnrB19 and qnrS1 exhibited low MIC to nalidixic acid (Supplementary Table S1).
Thirteen different Inc types were found in different isolates and different proportions. IncX1 (n = 12) was the most predominant Inc type detected and almost half of the 23 isolates (n = 13) harbored plasmids of two or more Inc types (Fig. 1). The Inc type sequences were found on contigs that were similar to plasmid sequences in the GenBank, except for contig-containing IncFIA in strains PM4, PD7, IncFIB in PI17, PK12, PM4, IncFIC in PK12, IncI1 in PD23 and IncY in PI24. These contigs could not assign the Inc to known plasmid sequences.
Nine different genes associated with virulence were identified in 23 of the 27 isolates, seven of them contained five or more virulence genes. The genes gad (glutamate decarboxylase), lpfA (long polar fimbriae), iss (increased serum survival gene), and astA (EAST-1 heat-stable toxin) were the most frequent virulence genes and were found in 23, 13, 11, and 8 isolates, respectively. Genes related to the enteroaggregative E. coli (EAEC) pathotype such as air (n = 2) and eilA (n = 3) as well as iroN (enterobactin siderophore receptor protein) (n = 1) were also detected but in lower proportion. Genes associated with the production of bactericidal proteins such as colicin M (cma) and microcinH47 (MccH47) were present in five and one isolates, respectively (Fig. 1).
Discussion
Our study showed that pigs from the Western region of Cuba harbored 3GC-R E. coli, similar to what has already observed in other studies worldwide.5,34,35 However, the results herein were obtained during a single prospective study that does not permit to determine the prevalence at farm or slaughterhouse level emphasizing the necessity for establishing a national monitoring program as implemented in several countries. 36 Even though 3GC are not used in pigs in Cuba, 3GC-R E. coli may have been introduced in pig husbandry through environmental and human contamination and/or selected and maintained in pigs by coselection after usage of other antibiotics authorized in Cuba for pigs such as tetracyclines (oxytetracycline, doxycycline, and tetracycline), aminoglycosides (streptomycin, kanamycin, and gentamicin), β-lactams (amoxicillin, cephalexin, and penicillin), sulfonamides (sulfadimidine, sulfamethazine, sulfamethoxypyridazine, and sulfathiazole), florfenicol, and trimethoprim. 37 Indeed, the majority of the isolates described herein also contained resistance genes to these classes of antibiotics. Despite quinolones are not used for pigs in Cuba, some isolates contained mutations in the QRDR of the topoisomerases as well as qnr genes, which have been shown to confer decreased susceptibility to these antimicrobials.38–40 However, four of the qnr-containing isolates exhibited low MIC to ciprofloxacin and nalidixic acid suggesting that the genes may not be expressed.
Contrary to the fact that CTX-M variants commonly detected in pigs are predominantly CTX-M-1 and CTX-M-14,5,16,41–43 none of them were found in the selected E. coli pig isolates from this region of Cuba. Other variants were detected with a predominance of CTX-M-32, followed by CTX-M-15, CTX-M-55, and CMY-2, although less frequent. The CTX-M-15 that is frequently associated with humans worldwide 44 was also recently reported to be the dominant ESBL in clinical E. coli from humans in Cuba. 22 It was mainly associated with the pandemic E. coli ST131 clone, 22 which was not found among the selected E. coli from pigs in our study. In Cuban pigs, CTX-M-15 was found in E. coli ST648 and ST162, which were also found in the human patients in Cuba, but less frequently than in ST131. 22 The CTX-M variants CTX-M-32 and -55 and CMY-2 were also identified in the E. coli of the Cuban patients, but in lower proportion and in E. coli of other STs. 22 Nevertheless, CTX-M-55 and CTX-M-32 have been increasingly reported in humans and animals and have been suggested to originate from animal sources, including cattle and pigs.5,42,45,46
Our results suggest that the spread of 3GC resistance in E. coli is being driven by an interplay between specific STs and mobile genetic elements. In the case of the blaCTX-M-32 gene, its potential for broader dissemination in porcine E. coli in Western municipalities of Cuba seems likely because of its presence in 12 different STs of E. coli from three different farms and at the two slaughterhouses. In addition, the blaCTX-M-32 gene was also reported in human patients in this country. 22 Both the diversity of Inc types found in E. coli carrying blaCTX-M-32 and the diversity of STs associated with blaCTX-M-32 carriage suggest that this ESBL gene is mainly spread by mobile genetic elements, which can be further disseminated if contained in conjugative plasmids. IncX1 was the most frequent Inc group in our study and it was detected in half of the isolates containing the blaCTX-M-32 gene. Complete genome sequence of one of the E. coli IncX1-positive E. coli (strain PK6, ST1695) confirmed the presence of blaCTX-M-32 on a 42,683-kb IncX1 plasmid, 47 but further plasmid-wide analyses need to be performed to better understand the epidemiology of plasmids in E. coli in this country. However, contrary to most reports stating that blaCTX-M-32 has been usually associated with IncN plasmids,48–50 our results agree with Ceccarelli et al. 43 where blaCTX-M-32 was found on IncX1 plasmids in E. coli from pigs and calves. The transmission of ESBLs by horizontal transfer between bacteria along the food chain has been widely suspected,5,51–53 and the discovery of porcine E. coli isolates sharing such genes with human strains in Cuba suggests that a potential risk may also exist for pork meat consumers and personnel working in the swine production industry.
The cgMLST analysis not only confirmed the genetic diversity of the majority of the selected isolates as observed by rep-PCR, but also explored the genetic relationship shared by isolates belonging to the same ST (Fig. 1). In fact, some of the isolates that were highly genetically related, differed by either their different antibiotic resistance or virulence genes. Genetically related isolates were also found in pigs raised on different farms suggesting possible clonal dissemination of 3GC-R E. coli in pig husbandry in this Western part of Cuba, as it has been previously reported in other countries.8,54,55 Other genetically related 3GC-R E. coli were found in pigs from different farms sampled at the same slaughterhouse, suggesting that acquisition of 3GC-R E. coli may occur at the slaughterhouse. Nevertheless, these isolates may also have been introduced earlier on the different farms with pigs having either the same origin or sharing the same contaminating sources. The latter seems to be less probable considering the geographical distance between these farms.
Some specific E. coli strains have been described as important pathogens causing both intestinal and extraintestinal infections either in animals or humans or in both. 56 The air, astA, and eilA genes, also detected in two pig isolates, have been widely associated with the EAEC in humans.57,58 Although these two multidrug-resistant strains lack the aggregative adherence fimbriae operon, the air, astA, and eilA virulence genes may play a role in their pathogenic potential. Other genes such as lpfA, iroN, and iss related to extraintestinal pathogenic E. coli pathotype, which was reported in a human patient in Cuba, were also found in this study. 22 Genes such as gad, cma, and MccH47 operon are related to the survival of E. coli under stress conditions and competition with other related species, favoring the ecological adaptation of these isolates.59–61
The presence of virulence genes in porcine E. coli together with antibiotic resistance genes (including 3GC-R genes) emphasizes the necessity for applying good hygiene and disinfection protocols along the meat production chain to avoid all types of bacterial contamination regardless of human, animal, or environmental origin.
Conclusions
A detailed characterization of 3GC-R E. coli from pigs from different farms and slaughterhouses in different municipalities of the Western region of Cuba using NGS gave some new insights into the molecular characteristics of such isolates in pig farming in the country. The presence of 3GC-R E. coli in each investigated facilities and genetic diversity observed among the isolates emphasized the need for surveillance programs and establishment of strategies for the prudent use of antibiotics in food animals in Cuba.
Footnotes
Acknowledgments
The authors thank Gisleibys Miranda, Yaday Alfonso, Alexandra Collaud, and Alexandra Rossano for the technical assistance. The authors also thank Neily Rivalta and the personnel from farms and slaughterhouses for their cooperation in the sampling.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the Swiss Government Excellence Scholarships for Foreign Scholar (no. 2018.0714) granted to R.E.H.-F., by the National Center for Animal and Plant Health (Cuba) and by the Institute of Veterinary Bacteriology of the University of Bern (Switzerland).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.