
Abstract
The occurrence and characterization of carbapenemase-producing Enterobacteriaceae from companion animals in Guangzhou, China, are investigated. Six isolates (2.3%, 6/257) were positive for blaNDM-5, that is, one Enterobacter cloacae, one Citrobacter freundii, and four Escherichia coli. Three E. coli isolates obtained from the same animal hospital were ST410 and showed identical pulse field gel electrophoresis pattern, resistance profiles, and resistance genes. blaNDM-5 was located on IncX3 (n = 5) and IncK2 (n = 1) plasmid, respectively. The presence of carbapenemase-producing Enterobacteriaceae among companion animals needs continued surveillance.
Introduction
Carbapenemase-producing isolates particularly New Delhi Metallo-β-lactamase (NDM)-encoding Enterobacteriaceae have been increasingly reported worldwide and have become a global challenge. 1 Although NDM-producing Enterobacteriaceae are mainly described in clinical samples, they have also been identified in food-producing animals, wildlife, and animal-derived food products.2–4 To date, some cases of NDM-producing Enterobacteriaceae have been described in companion animals in several countries, such as Korea, Egypt, and the United Kingdom, with NDM-5–encoding IncX3 being the major carriers.5–8 However, the blaNDM gene has been rarely report in companion animals in China, although blaNDM-1–carrying Escherichia coli ST167 was previously reported in a diseased dog, and blaNDM-5 was detected in three E. coli isolates (2.36%) from diseased dogs in Beijing.9,10 In this study, we report six Enterobacteriaceae isolates producing NDM-5 recovered from healthy and diseased companion animals in Guangzhou, China.
A total of 257 Enterobacteriaceae isolates, including E. coli (n = 203), Klebsiella pneumoniae (n = 36), Enterobacter cloacae (n = 16), and two Citrobacter freundii (n = 2), were obtained from healthy and diseased companion animals at four animal hospitals located in four districts in Guangzhou, China, during 2016–2017. Enterobacteriaceae isolates were obtained and species identification was performed as previously described. 11 Among them, 153 Enterobacteriaceae isolates collected in 2016 were previously described. 11 Details of the source and origin of the 104 isolates collected in 2017 are given in Supplementary Table S1. All 257 Enterobacteriaceae isolates were detected for the presence of carbapenemase genes by PCR and sequencing. 12 We found that one E. cloacae, one C. freundii, and four E. coli isolates carried blaNDM-5 (Table 1). The minimal inhibitory concentrations (MICs) of 12 antimicrobial agents were determined by the agar dilution method or broth microdilution method (limited to colistin). 13 All NDM-producing strains exhibited MIC of 2–8 mg/L to imipenem, and showed resistance to ampicillin, ceftazidime, cefotaxime, gentamicin, and sulfamethoxazole/trimethoprim, but susceptibility to colistin; five exhibited resistance to tetracycline and ciprofloxacin. In addition, two, one, and one isolates displayed resistance to amikacin, florfenicol, and fosfomycin, respectively (Table 1 and Supplementary Table S3). The NDM-5–encoding isolates were confirmed to carry other resistance genes by PCR and sequencing with primers listed in Supplementary Table S2, including blaCTX-M (four blaCTX-M-55 and one blaCTX-M-3), blaCMY-2 (n = 4), rmtB (n = 1), armA (n = 1), floR (n = 1), and fosA3 (n = 1) (Table 1).
Characteristics of New Delhi Metallo-β-Lactamase-5–Producing Enterobacteriaceae and Their Transformants
The transformants were designated as the name of their correspondent donor strains plus dash, number, and T (means transformant).
H, healthy; S, sick, specific disease was not recorded or unknown.
Antimicrobial susceptibility tests were performed and interpreted according to CLSI M100, 28th edition. Colistin (>2 mg/L) and florfenicol (>16 mg/L) were interpreted according to EUCAST. IMP, imipenem; AMP, ampicillin; CTX, cefotaxime; CAZ, ceftazidime; GEN, gentamycin; AMI, amikacin; TET, tetracycline; FFC, florfenicol; SXT, sulfamethoxazole/trimethoprim; FOS, fosfomycin; CIP, ciprofloxacin. All isolates were susceptible to colistin.
CLSI, Clinical and Laboratory Standards Institute; MIC, minimal inhibitory concentration; ORF, open reading frame.
The genetic diversity of blaNDM-5–positive isolates was analyzed by pulse field gel electrophoresis (PFGE), and was characterized by multilocus sequence typing (limited to E. coli).14,15 GZ7DS11 belonged to ST9437, and the remaining three E. coli isolates obtained from the same hospital belonged to ST410 (Table 1). Three ST410 NDM-5–producing strains exhibited identical PFGE patterns, with identical antimicrobial susceptibility profiles and resistance genes, indicating that clonal spread of blaNDM-5–positive E. coli had occurred among companion animals within the same hospital in Guangzhou (Table 1 and Supplementary Fig. S1). E. coli ST410 has been increasingly reported as NDM-5 carriers in companion animals,5–8 this lineage has been identified worldwide and has emerged as new international successful high-risk clones associated with resistance to carbapenems. 16
Conjugation experiments were performed using streptomycin-resistant E. coli C600 as the recipient strain as previously described, 17 selected using 1 mg/L imipenem and 3,000 mg/L streptomycin. Transformation was carried out by heat shock 18 using E. coli strain DH5α as the recipient strain, selected by 0.5 mg/L imipenem. The presence of blaNDM in the transconjugants/transformants was determined by PCR and sequencing. 12 The blaNDM-5 gene in six isolates was successfully transferred to E. coli strain C600 with frequencies of 10–5 to 10–2 transconjugants/recipient and DH5α with 103–105 CFU/mg (Supplementary Table S4). Transformants with a single blaNDM-positive plasmid were selected for further study, confirmed by S1-PFGE 19 and Southern blot hybridization according to the manufacturer's instruction. The results showed that blaNDM-5 was located on ∼46 kb plasmid (n = 5), or ∼104.5 kb plasmid (n = 1). Five ∼46 kb plasmids were classified as IncX3 plasmids, 20 whereas the replicon type of ∼104.5 kb could not be determined by PCR-based replicon typing. 21 The transfer of blaNDM-5–carrying plasmid caused resistance to ampicillin, ceftazidime, and cefotaxime, and increased MICs for imipenem (1–2 mg/L; four–eightfold). Resistance to sulfamethoxazole/trimethoprim was only observed in the transformant GZ7DS11-1T. Other resistance genes were not detected in any transformants.
Six NDM-5–encoding plasmids were extracted from their transformants using QIAGEN® Plasmid Midi Kit (Qiagen, Hilden, Germany) following the manufacturer's instructions, and were sequenced by Illumina Hiseq 2500 (Illumina, San Diego, CA, USA). Sequence reads were assembled into contigs with SOAPdenovo version 2.04. Plasmid contigs were assembled into the complete plasmid sequence with PCR and Sanger sequencing (limited to pHN7DS11; Supplementary Table S5). Analysis and annotation of plasmids were performed using the RAST server, 22 ISfinder, ResFinder, and BLAST. Integrons were analyzed using INTEGRALL. Three IncX3 plasmids (pHN7CS2, pHN7DH6, and pHN7DH7) obtained from three ST410 NDM-5–producing isolates were 46,161 bp long with identical nucleotide sequences, and pHN6DS3 differed from them by only one nucleotide at position 5934 resulting in amino acid change in ISKox3 transposase. The remaining IncX3 plasmid pHN7DS4 had a size of 45,907 bp, and was similar to other IncX3 plasmids in our study, differed by two nucleotide changes within the plasmid backbone (pHN6DS3, position 361, and 42579) or three nucleotide changes (pHN7CS2, pHN7DH6, and pHN7DH7, position 361, 5934, and 42579), and by deletions of 254 bp of the ΔTn2 that was truncated by IS3000 (Fig. 1A).

Sequence comparison of
The five IncX3 plasmids in this study were identical or highly similar to other NDM-5–producing IncX3 plasmids obtained from several members of Enterobacteriaceae of various origins worldwide (99.9–100% identity and 100% coverage) (Fig. 1A), as well as plasmids encoding other NDM alleles, for example, pM216_X3 (NDM-4, AP018146), pJN05NDM7 (NDM-7, MH523639), and pAD-19R (NDM-17, KX833071). The observation of identical or similar IncX3 plasmids found in our study further highlights the important role of the epidemic IncX3 plasmids in carbapenem resistance transmission, but also the potential for IncX3 plasmids to horizontally transfer between diverse species and hosts.
pHN7DS11 was 115,619 bp in size with an average GC content of 54.41%, and was identified as IncK2 plasmid. The backbone of pHN7DS11 was highly similar in organization to other IncK2 plasmids (96.9–99.9% identity and 66–94% coverage), possessing a complete array of genes involved in replication, conjugation, maintenance, and stability, such as pDV45 (blaCMY-2, 85,963 bp, KR905384), p1106-NDM (blaNDM-5, 113,687 bp, and MG825375), p92944-NDM (blaNDM-9, 118,644 bp, and MG838260), pEC1107-NDM-116K (blaNDM-5, 116,042 bp, and MG601057), pHNTH02–1 (blaNDM-9, 118,337 bp, and MG196294), pHNSD138–1 (blaNDM-9, 103,506 bp, and MG271839), and pMb1488 (mcr-1, 101,065 bp, and KY565558) (Fig. 1B). These plasmids mainly differed by the presence of a putative group II intron inserted upstream of the ssb gene (pHN7DS11) and variable shufflon region (Fig. 1B and Supplementary Fig. S2). The shufflon is a site-specific recombination system identified in the IncI1/I2 plasmids as well as IncK plasmids.23,24 It contains different DNA segments flanked by conserved 19-bp repeats (recombination sites), resulting in the diversity of the C-terminal end of the PilV protein. 24 The shufflon region of these IncK2 plasmids is highly diverse, with different combination of repeat sequences (previously designated as repeats e, f, g, and h), 23 such as pHN7DS11 (g-g-e-e-g-f), p1106-NDM (e-e-g-f-g-g), pEC1107-NDM-116K (h-g-g-g-e-e), and pDV45 (e-e-e-e-g-g-h-f) (Supplementary Fig. S2).
In addition to companion animal, similar IncK2 plasmids carrying blaNDM genes (blaNDM-5/-9) have been detected in E. coli isolates from food-producing animal 25 and food 26 and human clinic in China (Fig. 1B). However, the multiresistance regions (MRRs) of these IncK2 plasmids were varied, which may have resulted from the acquisition, deletion, and rearrangement events mediated by mobile elements (e.g., IS1294, IS26). In general, MRRs in NDM-producing IncK2 plasmids were all bounded by inverted repeat of ΔTn1721 and ΔTn5393 and inserted at the same location, which differed from other IncK2 plasmids (Fig. 2). The MRR in pHN7DS11 was related to those of other NDM-encoding IncK2 plasmids. In these plasmids, blaNDM-5/-9 gene was embedded within the blaNDM module associated with an ISCR1 complex class 1 integron and flanked by IS26 (IS26-intI1-dfrA12-gcuF-aadA2-qacEΔ1-sul1-ISCR1-cutA-tat-trpF-bleMBL-blaNDM-ΔISAba125-IS26) (Fig. 2). However, the class 1 integron was truncated by IS26, resulting in the deletion of a 4,109-bp segment in pHN7DS11 (Fig. 2). In pHNSD138-1, the whole blaNDM-9 segment was located in inverse orientation, one copy of IS1294 was inserted into the tat gene, and 765-bp of IS26 upstream of ΔISAba125 and 418-bp of 5′-CS of the class 1 integron along with 20-bp spacer were absent (Fig. 2). The MRRs of these IncK2 plasmids were further differed by the presence of the 3,317-bp fosA3 segment (Δorf3-orf2-orf1-fosA3-IS26) (pHNTH02-1 and p92944-NDM), by the loss of the 5,010-bp macrolides resistance module mph(A)-mrx-mphR(A)-IS6100-IS26 (p92944-NDM), by the insertion of IS1294 within ΔTn1721 and ΔTn21 (pHN7DS11), and by the acquisition of a 3,338-bp resistance segment (aac(3)-IV-aph(4)-Ia-ISEc59) and the deletion involving ΔIS5075, ΔTn21, the macrolides region and ΔTn2, and a 165-bp segment of ΔTn5393 (pHNSD138–1) (Fig. 2). In addition, pHNSD138-1 had a 301-bp shorter Tn1721 (Fig. 2). In addition to blaNDM genes, IncK2 plasmids were also described as vectors for blaCMY and mcr-1 in E. coli isolates from calves, poultry, poultry meat, and human in Europe.23,27–29 It highlighted the potential of IncK2 plasmids to become efficient vectors for blaNDM dissemination and to capture more resistance genes during horizontal spread.
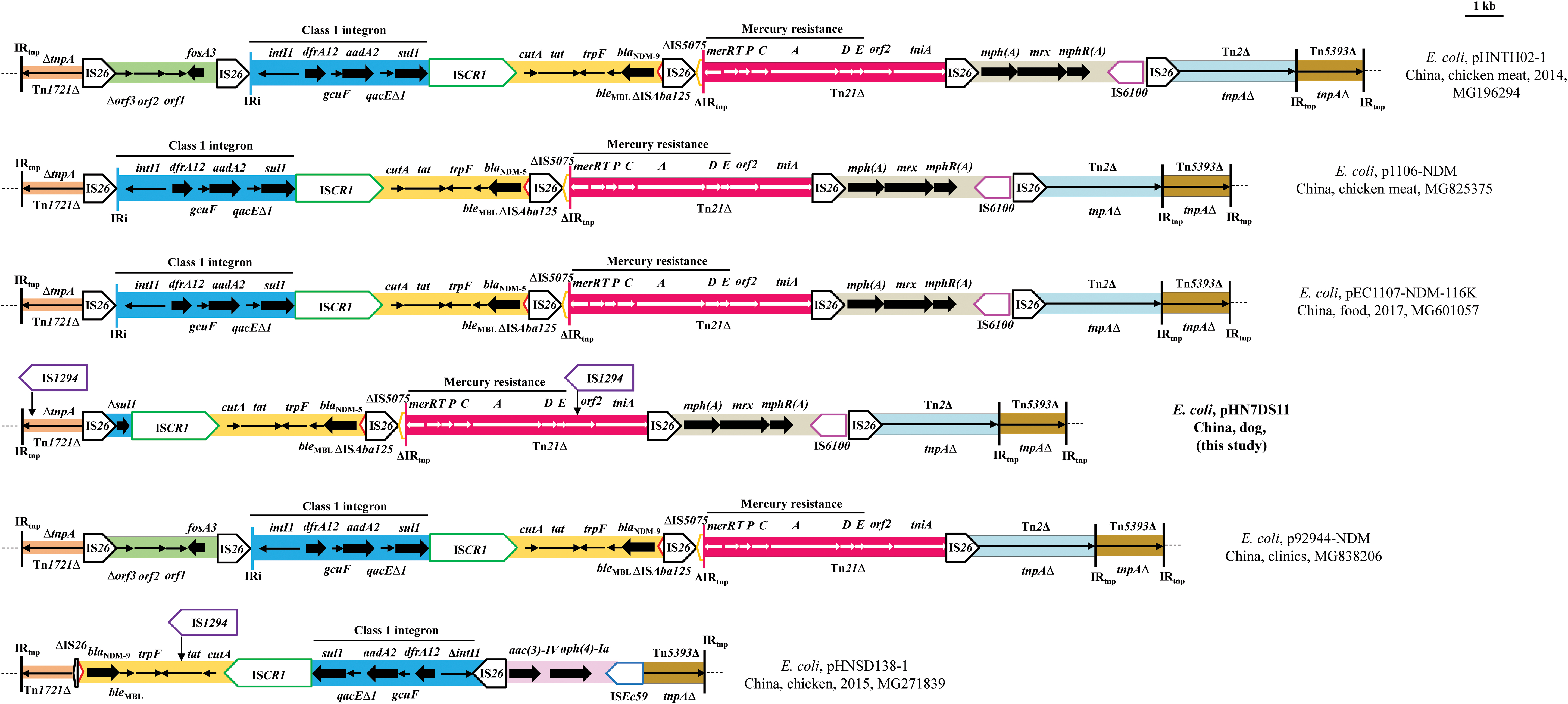
Genetic organization of the multiresistance region of plasmid pHN7DS11, and structural comparison with other IncK2 plasmids. The extents and directions of antibiotic resistance (thick arrows) and other genes are indicated. Δ indicates a truncated gene or mobile element. ISs are shown as boxes labeled with their name. Labeled vertical arrows with IS box indicate the insertion site of IS element. Tall bars represent the inverted repeats of transposon. The collection years of plasmids pEC1107-NDM-116K, pHNTH02-1, pHNSD138-1 are provided in Figure 2. IS, insertion sequence. Color images are available online.
In this study, the dissemination of blaNDM-5 in companion animals may be mainly associated with clonal spread of E. coli ST410 isolates within the same hospital, involving with the global successful pandemic blaNDM-carrying IncX3 plasmids. The horizontal transfer of IncK2 plasmid also facilitates the blaNDM-5 dissemination, and may further spread to other Enterobacteriaceae.
Nucleotide sequence accession numbers
Plasmids pHN6DS3, pHN7CS2, pHN7DH6, pHN7DH7, pHN7DS4, and pHN7DS11 have been deposited in GenBank under the accession numbers MN276078 to MN276083.
Footnotes
Acknowledgment
We are very grateful to Sally Partridge for her assistance in IncK2 shufflon analysis.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the National Natural Science Foundation of China (Grant Nos. 31625026 and 31830099).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.