
Abstract
Carbapenemase production in Enterobacterales clinical isolates is a global threat. Multi-drug resistant Klebsiella pneumoniae harboring carbapenemases are a major concern among the hospital settings in Latin America.
Aim:
The aim of this study was to analyze the genetic relatedness between three isolates of K. pneumoniae recovered from one patient in the same bacteriological round on the same day, which exhibited different susceptibility profiles to carbapenems (CP) and to colistin (Col). Isolates' profiles were as follows (susceptible-S/resistant-R): CPS/ColR, CPR/ColR, and CPR/ColS. Pulse-field gel electrophoresis, multilocus sequence typing, and whole genome sequencing were performed. Conjugation assays were carried out and PCR determination in transconjugants (Tcs) was made for: blaCTX-M-groups, blaNDM, blaKPC, blaTEM, qnr alleles, aac(6′)Ib-cr, ermB, and plasmid incompatibility groups (Inc).
Results:
All isolates belonged to the same clone, to ST258 and harbored blaCTX-M-14, blaCTX-M-15, qnrA1, qnrB1, aac(6′)Ib-cr, and wzi154 (capsule-locus KL107). One isolate had additional wzi gene, wzi109 (capsule-locus KL36). In CPR isolates, the pattern was explained for blaNDM-1 or blaNDM-1/blaKPC-2 presence, and in ColR for IS5-like element insertion in mgrB at different positions. Co-mobilization of blaNDM-1/qnrA1 was associated to a different plasmid Inc (A/C-FII) in both blaNDM-1 donors. Mobilization of blaCTX-M-14 was related to IncI1 in one donor.
Conclusion:
These findings highlight the potential plasticity of ST258 K. pneumoniae clone. To the best of our knowledge, this is the first description of blaNDM-1/blaKPC-2-producing K. pneumoniae ST258 in Latin America.
Introduction
The emergence of carbapenem resistance in enterobacteria is worrying, and this could be related to several combined mechanisms involving outer membrane permeability defects, hyperproduction of AmpC cephalosporinases, extended-spectrum β-lactamase, and carbapenemase production. 1 Carbapenemase production in Enterobacterales clinical isolates is a global threat and, in particular, Klebsiella pneumoniae harboring carbapenemases is the main problem in hospital settings in Latin America.1,2 Carbapenemase genes, blaKPC, blaOXA-48, and blaNDM-1, are responsible for carbapenem resistance in K. pneumoniae, whereas colistin resistance is frequently due to mutations that upregulate the PhoQ/PhoP system and pmrHFIJKLM operon, most commonly by inactivation of mgrB via IS insertions. 3 In recent years, transferable resistance to colistin, mediated by different variants of enzymes with phosphoethanolamine transferase activity, has been reported. To date, 10 families of genes coding for these enzymes called mcr-1 to 10 are recognized, several of which have also been reported in K. pneumoniae. 4
The worldwide dissemination of this pathogen has been predominantly linked to the high-risk clone group CG258, with ST258 being the most endemic in North and South America, including Uruguay.1,2 In previous reports, genomic analyses revealed that the most widely recognized antimicrobial resistance clone CG258 has evolved via a series of large-scale genomic recombination events, and it shows not only extensive capsular antigen but also antimicrobial resistance gene diversity, emphasizing the plasticity of this clone. 3
Even though ST258 is almost exclusively associated with K. pneumoniae carbapenemase (KPC), in this study we describe K. pneumoniae ST258 isolates obtained from a single patient from Montevideo, Uruguay, that presented different plasmid patterns associated with a New Delhi-metallo-B-lactamase-1 (NDM-1) or NDM-1 and KPC-2, and alterations in mgrB gene that explained different combinations of resistance profiles to carbapenems and to colistin. These findings highlight the potential plasticity of the ST258 K. pneumoniae clone and, as far as we know, is the first description of blaNDM-1/blaKPC-2 in ST258 in Latin America.
Materials and Methods
In the course of a Nosocomial Infection Surveillance Program on the occurrence of carbapenem-resistant Gram-negative bacilli, three K. pneumoniae isolates (Kpn09, Kpn10, Kpn11) were collected from a patient at the 26th day of hospitalization in an intensive care unit (Montevideo, Uruguay). Kpn09 and Kpn10 were recovered from rectal swabs, and Kpn11 was recovered from nasal swab. The patient received antibiotic treatment with amikacin, colistin, and fosfomycin before collection of isolates.
Identification and antibiotic susceptibility testing were performed by using the VITEK® 2 Compact system (bioMérieux, Marcy-l'Étoile, France) and interpreted according to CLSI guidelines. 5 Susceptibility to colistin was assayed by broth microdilution method recommended by EUCAST guideline (www.eucast.org). Carbapenemases were phenotypically determined by using KPC/MBL Confirm Kit (ROSCO Diagnostica A/S, Denmark).
XbaI-pulsed-field gel electrophoresis (PFGE) analysis was performed, and results were analyzed as previously reported. 6 Multi-locus sequence typing characterization was conducted according to the K. pneumoniae MLST database. (https://bigsdb.pasteur.fr/klebsiella/klebsiella.html).
We conducted next-generation sequencing by using Illumina MiSeq-I with Nextera XT libraries for the whole genome sequencing of Kpn09, Kpn10, and Kpn11 isolates and reads were assembled with SPADES v.3.11 by using k-mers 21, 33, 55, 77, 99, and 127 with “careful” option turned on and cut-offs for final assemblies: minimum contig/scaffold size = 500 bp; minimum contig/scaffold average Nt coverage = 10-fold. Contigs were re-ordered by using the ST258 reference genome (GenBank acc. no. CP006923) and oriented with Contiguator Tool (http://combo.dbe.unifi.it/contiguator). 7
Resistance genes and plasmid replicons were identified by using databases available at the Center for Genomic Epidemiology (http://www.genomicepidemiology.org). A maximum-likelihood tree of the complete genomes of K. pneumoniae Kpn09, Kpn10, and Kpn11, including the genome NJST258_1 as reference, was created by using MEGA7 based on core genome single nucleotide polymorphisms (SNPs) from whole-genome alignment obtained with SNP sites (https://github.com/sanger-pathogens/snp-sites). The substitution model used in MEGA7 for creating the tree was Hasegawa, Kishino, and Yano. A five-parameter model with 100 bootstraps. 7
Conjugation assays were carried out from blaNDM-1 positive donors, isolates Kpn10 and Kpn11 (meropenem resistant, rifampin and sodium azide susceptible) in three independent assays with receptor Escherichia coli strain Top10 (meropenem susceptible, rifampin, and sodium azide resistant). They were cultured for 18 hr at 35°C, in Luria Bertani Agar (LBA) containing meropenem 2 mg/L or rifampin 200 mg/L. Transconjugants (Tcs) were selected in LBA containing meropenem 2 mg/L and rifampin 200 mg/L and confirmed by positive growth in LBA containing sodium azide 100 mg/L and positive PCR for blaNDM-1 gene. 1 Conjugation frequencies were calculated as the number of Tc per receptor cell. PCR was done in Tc for: blaCTX-M-groups, blaNDM, blaKPC, blaTEM, qnr alleles, aac(6′)Ib-cr, and ermB.8–10 Plasmid incompatibility groups were detected by PCR using Tc genomic DNA as template. 11 Positive results were confirmed by Sanger sequencing, and sequences were analyzed by using available databases. Minimal inhibitory concentration to ciprofloxacin in Tc was determined by Etest according to the manufacturer's recommendations.
Results
All isolates were resistant to cefotaxime, ceftazidime, cefepime, ciprofloxacin, and trimethoprim/sulfamethoxazole and susceptible to amikacin, gentamicin, and fosfomycin. Isolates showed different susceptibility (susceptible-S/resistant-R) patterns to carbapenems (CP) and to colistin (Col): Kpn09-CPS/ColR, Kpn10-CPR/ColR, Kpn11-CPR/ColS (Table 1).
Phenotypic and Genotypic Variations in Klebsiella pneumoniae ST258 Isolates
Susceptibility test results, resistance genes, and plasmid incompatibility groups detected in transconjugants. Colistin BMD method (broth microdilution reference method).
Ciprofloxacin MIC determined by Etest.
BMD, broth microdilution reference method; KPC, K. pneumoniae carbapenemase; MIC, minimal inhibitory concentration.
Isolates shared more than 90% of similarity by PFGE analysis, indicating that they belonged to the same clone (data not shown) and the same sequence type was identified, ST258. The CPR pattern was explained by blaNDM-1/blaKPC-2 presence in Kpn10 and by blaNDM-1 presence in Kpn11. The ColR pattern was due to the insertion of IS5-like element in mgrB (position +74 in Kpn09 and +127 in Kpn10). Colistin resistance genes, pmrA, pmrB, pmrC, pmrD, and pmrE in Kpn09 and Kpn10 ColR isolates showed the same sequences present in Kpn11 ColS.
All isolates harbored blaCTX-M-14, blaCTX-M-15, qnrA1, qnrB1, aac(6′)Ib-cr (Table 1 and Fig. 1), and wzi154 (capsule-locus KL107).
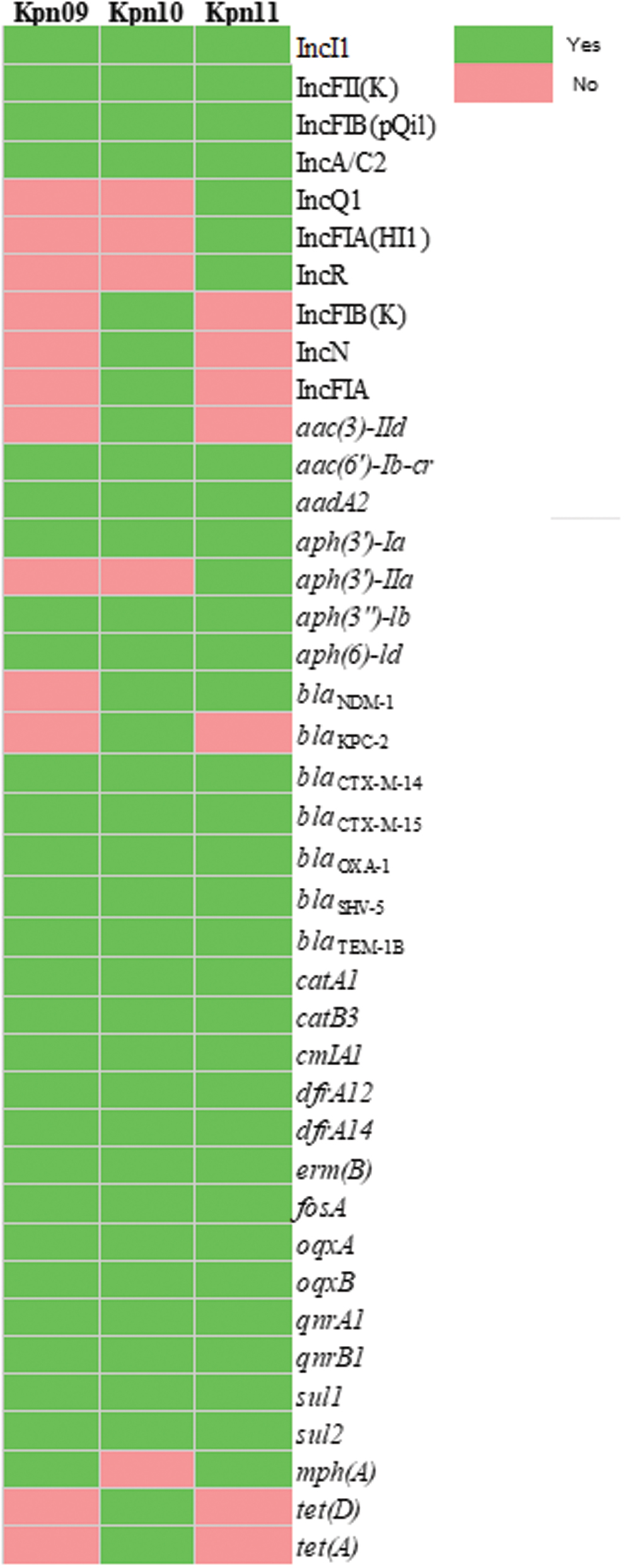
Comparison of plasmid replicons and resistance genes of ST258 Klebsiella pneumoniae isolates, Kpn09, Kpn10, and Kpn11—Uruguay, 2019. Accession nos. of the plasmid Inc group with 100% homology sequence with those present in Kpn09, Kpn10, and Kpn11: JN157804 (IncA/C2). MH674341.1 (Incl1), CP000648 [IncFII(K)], JN233705 [IncFIB(pQil)], AY046276, (IncN), AP001918 (IncFIA), DQ449578 (IncR), M28829 (IIncQ1), and AF250878 [IncFIA(HI1)]. Color images are available online.
Ten different Inc groups and 30 resistance genes were detected (Fig. 1). IncA/C2, IncI1, IncFII(K), and IncFIB(pQil) were present in all isolates according to other studies describing the main resistance-carrying Inc types in Klebsiella sp. 12 IncN, IncFIA, and IncFIB(K) were only present in Kpn10 and IncFIA(HI1); IncR, IncQ1 were only present in Kpn11. Genes conferring resistance to the main antibiotics of clinical use were detected.
Blast analysis from blaNDM-1 containing nodes of Kpn10 (node 98) and Kpn11 (node 66), of 5,338 bp in both cases, showed the following genetic environment: 100% identical (coverage 54%) to Acinetobacter baumanii strain AB17 plasmid pAB17 (GenBank acc. no. MT002974.1): ISAba125/blaNDM-1/bleMBL/trpF/tat.
On the other hand, blast analysis demonstrated that Kpn11 nodes 154, 279, 98, carrying blaCTX-M-14, blaTEM-1B, and ermB, respectively, revealed significant alignment with a previously reported IncI1 plasmid (GenBank acc. no. MH674341.1) from a multi-drug resistant E. coli clinical isolate in Uruguay. 13
Analysis from the wzi gene related to capsule locus exhibited the same allele for all isolates wzi154 related to capsule-locus KL107 (https://bigsdb.pasteur.fr). Interestingly, in Kpn10 we identified a second allele wzi109 related to KL36. In this isolate, the presence of a blaKPC-2 within an IncN plasmid backbone (node 51, 22,849 bp) is another distinct feature in comparison to Kpn09 and Kpn11, which showed a 100% sequence identity with an IncN plasmid containing blaKPC-2 in an E. coli from our country (MH514861.1).
The phylogenetic tree obtained clustered Kpn09, Kpn10, and Kpn11 isolates together, showing a high genetic relationship between them with a small difference on average of 45 single nucleotide polymorphism (SNPS) in the core genome (Supplementary Fig. S1).
We analyze the quinolone resistance determining region and mutations detected were the same in all isolates: gyrA Ser83Ile, gyrB Glu466Asp, parC Ser80Ile, parE wild type.
Different results were found for conjugation assays in blaNDM-1 donors. In TcKpn10, blaNDM-1/qnrA1 and IncA/C-FII were detected; whereas in TcKpn11, blaNDM-1/qnrA1/blaCTX-M-14/blaTEM-1/ermB and IncA/C-FII-I1 were observed (Table 1). Conjugation frequency of 1 × 10−6/receptor cell was detected from both donors.
Discussion
In this work, we describe the presence of three isolates of K. pneumoniae ST258 that presented different plasmid patterns and chromosomal modifications that explained different resistance profiles to carbapenems and to colistin. It is remarkable that isolates were obtained from one patient in the same bacteriological round on the same day.
In relation to carbapenem resistance profile, we characterized the genetic background of blaNDM-1 and found an association with ISAba125 and bleomycin resistance gene 14 ; this constitutes the first description of the blaNDM-1 surrounding region in our country.
According to colistin, the resistance pattern could be possibly explained due to the insertion of the IS5-like element in the mgrB gene. The type and positions of insertion described here were previously reported.8,15 The other colistin resistance genes, pmrA, pmrB, pmrC, pmrD, and pmrE in Kpn09 and Kpn10 ColR showed the same sequences present in Kpn11 ColS, indicating that these genes are not involved in the Col resistance phenotype as in the case of mgrB gene.
The presence of another wzi allele in Kpn10 could have been possible due to recombination events in this isolate, which are well known in capsular locus from K. pneumoniae. 16 Capsule locus wzi109/KL36 was associated to ST437, 17 and it is notably their characterization in an ST258 isolate.
Blast analysis for nodes carrying blaCTX-M-14, blaTEM-1B, and ermB in Kpn11 that demonstrates a significant alignment with IncI1 plasmid previously reported from our country was consistent with the results observed in its Tc, where the three genes were co-mobilized in association with IncI1. 13
Meanwhile, taking into account that the plasmid incompatibility groups associated with blaNDM-1 were found in both Tcs, this might suggest that the meropenem-susceptible phenotype of Kpn09 could be explained by the unstable nature of the transposable NDM locus. 18
On the other hand, all isolates had the same mutations in the quinolone resistance determining region. Mutations in gyrA/parC described here were reported in another study. 19 Interestingly, a specific mutation in gyrB was not reported, as far as we know, for K. pneumoniae. These results are consistent with the clonal origin of the isolates. Regarding acquired quinolone resistance mechanisms, qnrA1 was transferred to TcKpn10 and TcKpn11, but these Tcs remain susceptible to quinolones. qnr genes are responsible for low-level quinolone resistance, with an increase in the minimum inhibitory concentration of ciprofloxacin for wild-type E. coli J53 from 0.016 to 0.25 mg/L. This reduced susceptibility is likely most important for the selection of mutants with higher-level resistance. 20
Carbapenemases blaNDM-1 and blaKPC-2 were recently being reported in the Raoultella ornithinolytica strain co-harbouring both genes from urban river sediment. Interestingly, this bacterial species is closely related to Klebsiella spp. 21
It is remarkable that we could determine different patterns of incompatibility groups and resistance genes between the three isolates, revealing the role of lateral genetic transfer as the driven force for multi-resistance development. This reinforces the availability of ST258 clone to acquire different resistance mechanisms.
All findings reported in this study highlight the potential plasticity of the ST258 K. pneumoniae clone, which could colonize or cause an infection in a patient under different antibiotic selective pressure. The detection of different carbapenemases and their genetic location in clinical isolates has a major concern to take correct measures to decrease the spread of these platforms.
To the best of our knowledge, this is the first description of blaNDM-1/blaKPC-2-producing K. pneumoniae ST258 in Latin America.
Data Availability
The genome sequences data of Kpn09, Kpn10, and Kpn11 isolates have been deposited at DDBJ/ENA/GenBank under the accession JAAOKU000000000, JAAOKV000000000, and JAAOKW000000000, respectively. The versions described in this article are version JAAOKU010000000, JAAOKV010000000, and JAAOKW010000000.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.