
Abstract
Background:
Klebsiella pneumoniae (Kp), a common multidrug-resistant pathogen, causes a wide spectrum of nosocomial infections with high rates of morbidity and mortality. The emergence of pan drug-resistant international high-risk clones such as ST258, ST14, ST15, ST147, and ST101 is a global concern. This study was performed to investigate the carbapenemases, the plasmid profile, and the clonal relationship among Indian K. pneumoniae.
Materials and Methods:
A total of 290 K. pneumoniae isolates from seven centers in India were characterized to determine sequence types (STs) and carbapenemases. A subset of isolates was subjected to whole genome sequencing and hybrid genome assembly to obtain the complete genome. Plasmids carrying carbapenemases were characterized to determine the dissemination of carbapenem-resistant (CR) K. pneumoniae.
Results:
From this study, 75 different STs were observed with ST231 being predominant. About 79% of the analyzed isolates were CR with 59% (n = 136) producing OXA48-like carbapenemases. While ST231 was the predominant clone among the OXA48-like producers; NDM producers and NDM+OXA48-like producers were mostly associated with ST14. Interestingly, 61% (n = 138) of the total CR K. pneumoniae were colistin resistant, belonging to 22 different STs. Plasmid profiling shows that blaOXA48-like was exclusively carried by ColKP3, whereas blaNDM was associated with IncFII-like plasmids.
Conclusion:
The highly mosaic genome of K. pneumoniae coupled with the diverse ecological niches in India makes it a hotspot for antimicrobial resistance, leading to increased morbidity and mortality. Extensive molecular surveillance of the clonal spread of K. pneumoniae could help in understanding AMR dynamics and thus rework therapeutic management.
Introduction
Klebsiella pneumoniae, a common multidrug-resistant (MDR) pathogen, causes a wide spectrum of nosocomial infections, including pneumonia, bacteremia, urinary tract infections, and surgical site infections. 1 The use of carbapenems to treat K. pneumoniae infections resulted in the global emergence of carbapenem-resistant K. pneumoniae (CRKp). 2 Klebsiella pneumoniae carbapenemase (KPC) producing K. pneumoniae have high-level resistance to carbapenems and are strongly associated with high mortality and prolonged hospitalization worldwide. Although KPC is predominant in Europe and the United States, NDM and OXA48-like carbapenemases are endemic in India.3,4 Resistance to colistin, one of the few remaining treatment options for carbapenemase producing (CP) K. pneumoniae infected patients, is on the rise among CRKp in India, predominantly due to chromosomal mutations.5–7 In addition, the increased occurrence of the hypervirulent variant of K. pneumoniae (hvKp) and more recently the emergence of MDR-hypervirulent isolates from China and South East Asia has made hvKp a recognized global threat.8,9
There are international high-risk clones (HRC) described among K. pneumoniae, which are MDR and are not region specific. These include CC258 (ST11, ST258, and ST512), CC15 (ST14, ST15), ST147, ST101, ST17, and ST37.3,9 ST258 is the predominant KPC-2 producing K. pneumoniae clone reported globally. 9 CC23 is the predominant hypervirulent K. pneumoniae clonal complex, initially widespread in South East Asia but now emerging in other regions across the globe.8,10 Earlier, this clonal complex was susceptible to antimicrobials, but recently MDR and extensively drug-resistant (XDR) ST23 have been reported.10–12
Little is known about the prevalence of CRKp clones in India and their plasmid profile. This study aimed to determine the HRC and the mobile genetic elements (MGEs) that play an important role in the dissemination of antimicrobial resistance among K. pneumoniae in India.
Materials and Methods
Bacterial isolates
A total number of 290 K. pneumoniae isolates were collected between 2013 and 2018 for a 6-year period from seven Indian centers. The centers include Christian Medical College, Vellore (n = 163); All India Institute of Medical Sciences (AIIMS), New Delhi (n = 38); AIIMS, Raipur (n = 11); P.D. Hinduja Hospital and Research Centre, Mumbai (n = 38), Woodlands Multispeciality hospital, Kolkatta (n = 11); Fortis Hospital, Mahim (n = 15) and Choitram Hospital and Research Centre, Indore (n = 14). Representative isolates were collected from Vellore over the 6 years and the isolates from other centers were collected during 2016 and 2017. The isolates were recovered from various specimen sources, including blood, sputum, endotracheal aspirate, bronchoalveolar lavage, and pus specimens. Bacterial species were identified using standard microbiological biochemical methods 13 and confirmed with Vitek-MS (bioMerieux, France). The isolates were stored in glycerol stock at −80°C until use. The study was approved by the institutional review board (IRB) of Christian Medical College and Hospital, Vellore, India with minute number 9616 (2015).
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was performed by Kirby Bauer disk diffusion method according to CLSI guidelines. 14 Cefotaxime (30 μg), ceftazidime (30 μg), piperacillin/tazobactam (100/10 μg), cefoperazone/sulbactam (75/30 μg), imipenem (10 μg), meropenem (10 μg), ciprofloxacin (5 μg), levofloxacin (5 μg), gentamicin (10 μg), amikacin (30 μg), and minocycline (30 μg) were the antimicrobials tested. Commercial antibiotic disks were procured from Oxoid, England. Escherichia coli ATCC 25922, Enterococcus faecium ATCC 29212, and Pseudomonas aeruginosa ATCC 27853 were used as the quality control strains for susceptibility testing. Results were interpreted according to CLSI breakpoints.14,15
For all the isolates colistin susceptibility was determined by broth microdilution as recommended by EUCAST guidelines using colistin sulfate (Sigma-Aldrich). mcr-1 positive E. coli NCTC 13846 (Courtesy: Dr. Olga Perovic, National Institute for Communicable Diseases, Johannesburg) was used as positive control and E. coli ATCC 25922 and P. aeruginosa ATCC 27853 were used as quality controls. MIC results were interpreted as per EUCAST guidelines 2016. 16
Genomic DNA extraction
Total genomic DNA from all the isolates was extracted from an overnight culture (14–16 hours) grown at 37°C on blood agar using QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. The extracted DNA was quantified using Qubit (Life Technologies, Carlsbad, CA) and stored at −20°C until further use.
Detection of resistance and virulence genes
Molecular characterization of carbapenemase genes was performed for all the carbapenem-resistant (CR) isolates using multiplex PCR to detect blaIMP, blaVIM, blaNDM, blaOXA48-like, and blaKPC using primer pairs previously published. 4 The key virulence genes rmpA and rmpA2 were amplified using specific primer pairs as described previously.17,18 The amplified PCR products were separated using 2% agarose gel, which was stained with ethidium bromide.
Multilocus sequence typing
The genotypic diversity of K. pneumoniae isolates were evaluated using multilocus sequence typing (MLST) based on the seven housekeeping genes as described by Diancourt et al. 19 Seven housekeeping genes used to determine the clonal type among K. pneumoniae include gapA, infB, mdh, pgi, phoE, rpoB, and tonB. The sequence type (ST) was assigned by determining the allele number for each of the housekeeping gene using the database maintained by Pasteur Institute. Novel STs identified in this study were submitted to the database.
Whole genome sequencing and hybrid assembly
A subset of eight isolates carrying blaNDM, blaOXA48-like and/or both these, belonging to diverse clones were chosen for whole genome sequencing (WGS) to determine the plasmids responsible for carbapenemase dissemination. For short-read sequencing, the genomic DNA samples were subjected to WGS using the Ion Torrent PGM platform with Ion 318™ chip v2 for 400 bp chemistry sequencing (Life Technologies). DNA library was prepared using 1 μg of the genomic DNA using Ion Xpress Plus Fragment Library Kit (Life Technologies) following the protocol recommended for 400 bp fragment library preparation. For long-read sequencing, Oxford Nanopore MinION sequencing platform was used with FLO-MIN106 R9 MinION flow cells. Long read DNA library was prepared using the SQK-LSK108 ligation sequencing kit (v. R9) along with ONT EXP-NBD103 Native Barcode Expansion kit following the manufacturer's protocol (Oxford Nanopore Technologies, Oxford, UK). The library was loaded onto the flow cells, run for nearly 48 hours using the standard MinKNOW software. The Fast5 files generated from MinION sequencing was subjected to base calling with Albacore software (v.2.0.1) and the adapter sequences were trimmed off using Porechop.
Hybrid genome assembly and evaluation
Hybrid de novo assembly, quality check, and associated assembly statistics were performed using Ion Torrent and ONT MinION reads as described previously. 20 For accurate assembly, the MinION long reads were error-corrected with the standalone Canu error correction tool (v.1.7) using the “-correct-nanopore-raw” module. 21 This was followed by hybrid de novo assembly using both Ion torrent and MinION reads using the Unicycler hybrid assembly pipeline (v.0.4.6) for prokaryotic genomes with the default settings. 22 The complete circular genome was initially assembled and was polished with multiple rounds of Pilon (v.1.22) to reduce the base level errors as described previously. 23 Genome assembly by combining the Ion Torrent short reads and the MinION long reads improved the contiguity and completeness of the genomes with fewer error rates. The quality measurements of the complete genome sequences after hybrid assembly was compared with the short reads assembled de novo using SPAdes v.3.12 algorithm 24
To validate the quality of assembly after each polishing step, the completeness, correctness, and contiguity parameters were assessed using CheckM v1.0.5 25 and Quast v4.5. 26 CheckM estimated the completeness and contiguity, whereas Quast was used to detect misassemblies, mismatches, and indels by aligning the assemblies with the reference genome (AP006725).
Phylogeny
Minimum spanning tree (MST) was constructed using the allelic profiles of the STs observed in the study with the help of online tool PHYLOViZ. The database of K. pneumoniae MLST with 5189 profiles was used as the auxiliary data to generate the MST. Clonal complexes were defined as STs that differed from each other in ≤3 allelic profiles.
Results
STs identified among K. pneumoniae isolates in this study
From a total of 290 isolates, 75 different STs belonging to 13 different clonal complexes (CC) were identified among which CC231 (26%, n = 75) and CC14 (22%, n = 65) were predominant. The clonal complexes and respective STs observed in this study and its prevalence is mentioned in Table 1. ST147, ST231, and ST395 were the predominant STs among the isolates from all centers. International STs observed among the studied isolates were ST23, ST11, ST14, ST147, and ST231. A MST of MLST of K. pneumoniae constructed using PHYLOViZ online is shown in Supplementary Fig. S1 and the clonal complexes observed are similar to those mentioned in Table 1.
Clonal Complexes, Sequence Types, and Associated Predominant Carbapenemases Seen Among Klebsiella pneumoniae in India
CC, clonal complex; CSKp, carbapenem susceptible Klebsiella pneumoniae; KPC, Klebsiella pneumoniae carbapenemase; ST, sequence type.
CC29 was the most predominant (23%, n = 14) among the 60 carbapenem susceptible K. pneumoniae (CSKp) followed by CC23 (13%, n = 8). The presence of international clones such as ST14, ST147, and ST231 that are frequently MDR were observed among the CSKp. In contrast, among CRKp, CC231 was predominant contributing to 32% (n = 73) followed by CC14 (27%, n = 62). CSKp were more diverse showing 35 different STs among 60 isolates, whereas 230 CRKp were distributed among 48 diverse STs. OXA48-like was the predominant carbapenemase present in 59% of CRKp (n = 136) and these isolates predominantly belonged to ST231 (n = 60, 44%) although it was distributed among 22 different STs such as ST11, ST14, ST15, ST147, and ST395. NDM was present in 12% of CRKp (n = 28) and was distributed among 11 different STs, including ST11, ST14, ST15, ST23, ST147, ST231, and ST395. CRKp that coproduced NDM and OXA48-like contributed to 18% (n = 41) of CRKp and were present among isolates of 15 different STs, including ST11, ST14, ST147, ST101, ST231, and ST14. Some of the CR isolates lacking carbapenemases accounted for 9% (n = 21) and belonged to 15 different STs, including ST11, ST147, ST231, and ST395. Three KPC K. pneumoniae isolated from Vellore belonged to three different STs: ST15, ST101, and ST2957.
About 61% (n = 138) of the total CRKp isolates analyzed in this study were resistant to colistin. They belonged to 22 different STs with ST231and ST14 being predominant. Three major CCs were observed among the colistin-resistant CRKp, including CC11, CC14, and CC16. ST3249, a novel triple locus variant of ST15, was identified. Notably, STs of Col-R K. pneumoniae were similar to CRKp and no specific STs of Col-R CRKp were identified.
STs of hypervirulent K. pneumoniae
A total of 12 hypervirulent isolates were identified among the analyzed isolates. These isolates were string test positive and carried rmpA and/or rmpA2. Among these 12 isolates, 6 were pan-susceptible, 1 was an extended spectrum beta-lactamase (ESBL) producer, 4 were CR, and 1 was resistant to carbapenem and colistin. Four pan-susceptible hvKp and the colistin-resistant hvKp belonged to ST23 (the international hypervirulent clone). The two other pan-susceptible and ESBL hvKp belonged to ST2318, ST2319, and ST2320. Four CR hypervirulent isolates were identified belonging to MDR clones such as ST11, ST43, and ST231.
Novel STs identified
The six novel STs identified were ST3249, ST3418, ST3419, ST3420, ST3421, and ST3422. ST3418 is a single locus variant of ST43. ST3419 is a double locus variant of ST188, but ST188 was not seen in this study. ST3420 is a triple locus of ST17. ST3421 is a single locus variant of ST1777, whereas ST3422 is a single locus variant of ST231, the predominant clone in this study. The newer STs have evolved from the prevailing clones in India.
Antimicrobial resistance spread through plasmids:
The common carbapenemases seen among Indian CRKp are NDM and OXA48-like, which are carried by diverse clones on various plasmids. From the hybrid genome assemblies performed on CRKp, it was observed that blaOXA-232 was present on ColKP3, whereas blaOXA-181 was present on ColKP3 and IncFIB. blaNDM-5 was carried on IncFII plasmid among isolates belonging to ST147, ST2096 (CC14), and ST437 (CC11). The isolates producing dual carbapenemases such as blaNDM and blaOXA48-like carried these genes on different plasmids types. Interestingly, the uptake of plasmids by the prevalent clones leads to an increase in MDR and XDR K. pneumoniae. Table 2 lists the AMR genes carried by various plasmids among diverse clones as determined by hybrid genome assembly. It is noteworthy that the same plasmid type carried different AMR genes among isolates belonging to different STs. For example, IncFII in ST23 carried a single AMR gene, aac(6′)-lb-cr, whereas the same plasmid type in ST16 carried eight AMR genes as mentioned in Table 2. The structure of the ColKP3 plasmid was similar among the Indian and global isolates (Fig. 1), unlike other plasmid types that vary due to differences in the MGEs such as insertion sequences and transposons mobilizing AMR genes. ColKP3 is found to be conserved and carried truncated ISEcp1 adjacent to blaOXA-232. In addition, ColKP3 was also found among ST14, the NDM clone that is also dual carbapenemase producer.
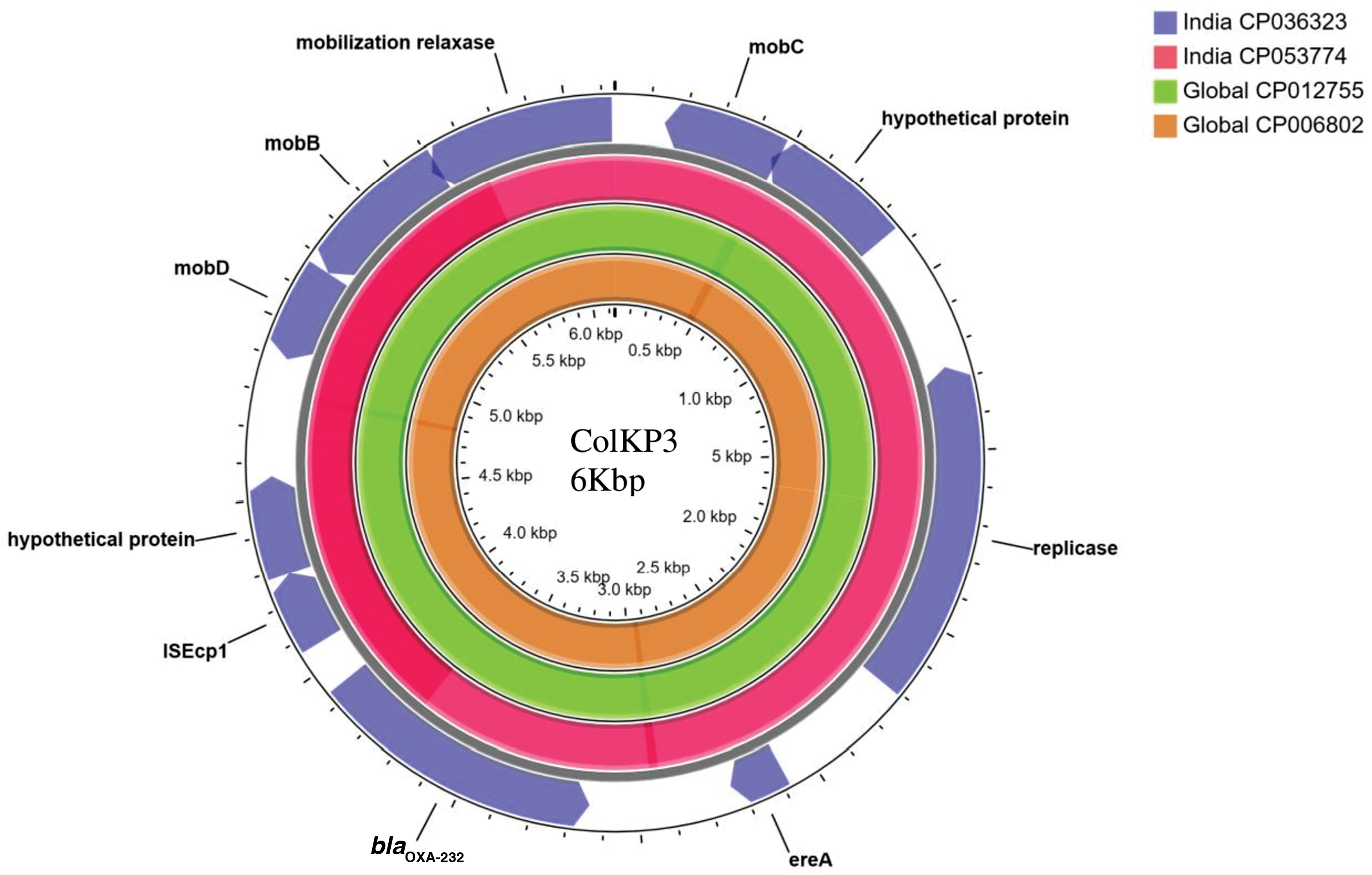
Circular representation of ColKP3 plasmids assembled from Klebsiella pneumoniae isolates displayed using CG view server. Comparison of ColKP3 plasmid carrying blaOXA-232 among (1) K. pneumoniae strain VBA2172 (CP036323) from ST231 (2) K. pneumoniae strain BP3636 (CP053774) from ST2096 (3) K. pneumoniae strain KP617 (CP012755) from ST14 (4) K. pneumoniae strain PittNDM01 (CP006802) from ST14. Truncated ISEcp1 is present adjacent to the carbapenemase encoding gene, blaOXA-232. ST, sequences type. Color images are available online.
Mobile Genetic Elements Associated with Antimicrobial Resistance Genes Observed Among Various Clones of K. pneumoniae in India as Derived by Hybrid Genome Assembly
Carbapenemases are highlighted in bold.
IncR plasmid carried the resistance genes.
There were significant differences observed among the plasmid sequences of IncFIBK and IncFIB(pQil) among the studied isolates. On comparison of the IncFIB(pQil) plasmid among the two isolates, each belonging to ST23 and ST231, it was found that the IncFIB(pQil) in ST23 was larger in size compared with that of ST231 (164 Kbp vs. 127 Kbp). The differences were due to the loss of MGEs carrying AMR genes (Fig. 2). blaTEM-1B and qnrS1 were absent in IncFIB(pQil) of ST23. Genes coding for aminoglycoside resistance such as rmtF and aac (6)-lb3 were carried on the class1 integron on IncFIB(pQil) plasmids (Fig. 2). The integron also carried ARR-2 coding for rifampicin resistance and the cat gene responsible for chloramphenicol resistance. The IncFIBK of BA1559 (ST15, CP036188) carried blaCTX-M-15 associated with the ISEc9 family transposase in a 2389 bp segment. This segment was absent among the IncFIBK of other isolates compared. Besides, blaCTX-M-15 was also carried on other plasmid types such as IncHI1B and IncR.
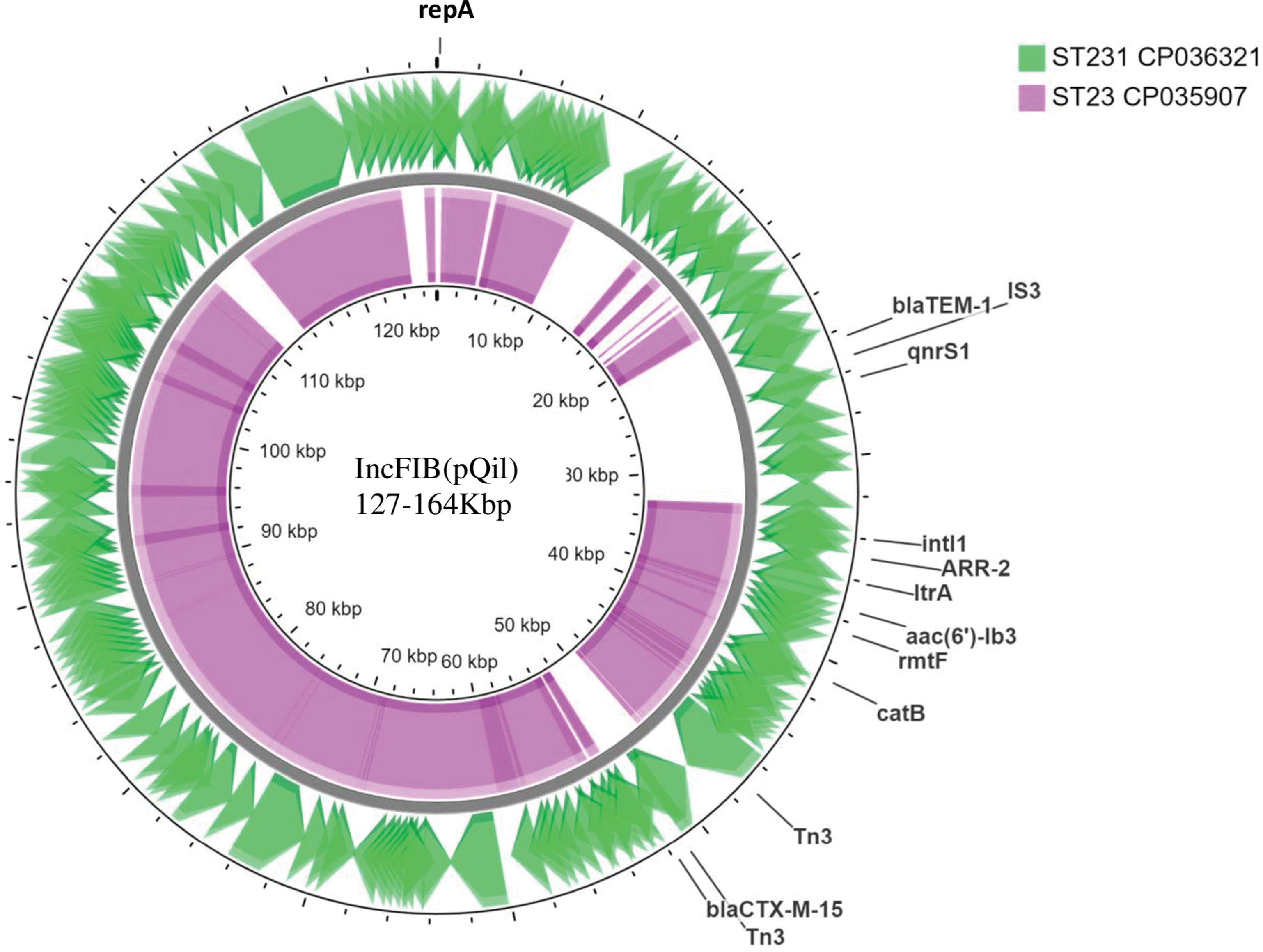
Circular representation of IncFIB(pQil) plasmids assembled from K. pneumoniae isolates displayed using CG view server. Comparison of IncFIB(pQil) plasmid among (1) K. pneumoniae strain BA2172 (CP036321) from ST231 (2) K. pneumoniae strain BA4656 (CP035907) from ST23. Color images are available online.
The plasmid profile of global isolates belonging to clones such as ST11, ST147, and ST231 were also determined. Plasmid, pNDM-MAR with an IncFIB or IncHI1B backbone was found among NDM producing clones such as ST14 and ST15 from the global collection (CP008933.1 and CP006799.1). It carried multiple AMR genes, including aac(6′)-Ib-cr, aadA2, armA, blaNDM-1, blaOXA-1, qnrB1, catB3, dfrA12, sul1, msrE, and mphE, thus conferring resistance to several classes of antibiotics. Surprisingly, among NDM producing ST147, blaNDM was also carried on IncFIB(pQil) (CP014757.1). The isolates from the global collection also carried either blaOXA-232 alone or in combination with ereA on the ColKP3 plasmid (CP006802.1 and CP012755.1). ColKP3 is becoming increasingly prevalent especially mobilizing blaOXA-232 and blaOXA-181 among blaOXA48-like carbapenemases.
Discussion
K. pneumoniae is the most common MDR pathogen isolated in Indian hospitals and is associated with several outbreaks.27–29 However, the characterization of circulating K. pneumoniae clones has not been investigated in India. This study is the first multicentric study to describe the predominant clones in India and the MGEs present in these clones.
The clonal distribution of K. pneumoniae in India is unique as ST231, CC14, and CC11 are the most prevalent clones. Interestingly, the dominant clones widely distributed in Europe and North America such as ST258 and ST512 are not present in India. 3 The international HRC such as ST11, ST14, ST15, and ST147 reported from Asia and Europe are seen in India. 9 The Indian endemic clone ST231 (∼26%) is mostly associated with carbapenem resistance but is not often reported in other countries.30,31 Remarkably, 75% of the prevalent clones of K. pneumoniae carry carbapenemases that drive the successful clonal expansion.
The convergence of virulence and AMR in K. pneumoniae is being increasingly observed among the characteristic hypervirulent clone ST23 and also the MDR clones. 32 ST23 carries the virulence plasmid with an IncHI1 backbone possessing the key virulence factors rmpA, rmpA2, and iutA. 33 Classically, ST23 is susceptible to all antimicrobials. However, recently, hvKp belonging to ST23 has been observed carrying blaOXA232 on ColKP3 in India and China.12,34 This is concerning as it poses the dual threat of resistance and virulence, challenging the patient management.
Recent estimates show 60% of CRKp in India harbors blaOXA48-like followed by coproducers of blaNDM and blaOXA48-like (20%) and blaNDM alone (15%). In contrast, blaKPC-2 and blaKPC-3, which are the most prevalent carbapenemases globally, hardly exist in Indian collection.4,35 From this study, it is also evident that about half of the blaOXA48-like producing CRKp belong to ST231, whereas most of the blaNDM carrying CRKp were distributed among CC11. The coproducers of blaNDM and blaOXA48-like were mostly carried by CC43 and CC11. However, comprehensive global epidemiological data suggest the clonal expansion of ST258 carrying blaKPC-2 and blaKPC-3 in North America, Europe, and China, 3 whereas ST231 harboring blaNDM and blaOXA48-like is established in India. Interestingly ST11 has established as a blaNDM carrying clone in Asia, whereas the same ST has expanded in Europe as blaKPC clone.4,36
The clonal expansion of K. pneumoniae is primarily driven by the acquisition of AMR genes by horizontal gene transfer of plasmids. Globally, the successful spread of ST258 with blaKPC is by employing IncFII group of plasmids3,36–38 and Tn4401 and its variants, 3 whereas in India ST231 has expanded by acquiring blaOXA48-like exclusively aided by the ColKP3 plasmid. 31 Globally blaOXA48-like is being associated with the IncX, IncA/C, ColKP3, and IncL/M plasmids.3,38 To the best of our knowledge, in India, there have been no reports of blaOXA48 carried on IncL/M with Tn1999.3,31 The variants of OXA48-like seen in India are OXA-181 and OXA-232, which are carried mostly on Col plasmids and were associated with truncated ISEcp1. 39
Unlike blaKPC and blaOXA48-like that are carried by a particular group of plasmids, blaNDM is mobilized by diverse plasmid families. This is similar to the international HRC ST11, ST14, and ST147 that carry blaNDM on IncH, IncF, IncX, and IncA/C. 3 In addition, truncated ISAba125 is often associated with blaNDM spread among members of Enterobacterales across many countries. 3
However, we have observed that the proportion of various carbapenemases varies widely between hospitals of different level of care and geographic region, reflecting the infection control practice of the institution. The plasmid-mediated carbapenemases drive CRKp expansion. Plasmid borne clonal dissemination of CRKp can be contained with best infection control practices. The prudent use of colistin using antimicrobial stewardship can help reduce colistin resistance as colistin exposure is the major risk factor.
Remarkably, among Enterobacterales, a greater proportion of carbapenem resistance is seen among K. pneumoniae worldwide. This can be attributed to high genome plasticity and relatively lower fitness cost of K. pneumoniae resulting in the clonal expansion of AMR lineages; genome-wide analysis of global isolates shows an average of three plasmids and ∼3000 accessory gene per genome emphasizing alterable nature of the genome and the high predilection of K. pneumoniae to acquire multiple AMR plasmids.37,40 Furthermore, among K. pneumoniae in India, an average number of five plasmids were present, which is higher than the global average of three plasmids per genome of K. pneumoniae.
Conclusion
In India, we experience a relatively higher proportion of CRKp infections compared with global numbers. The highly mosaic genome of K. pneumoniae coupled with its diverse ecological niches in India makes it a hotspot for antimicrobial resistance. ST11, ST14, and ST147 carry blaNDM on plasmids such as IncH, IncF, and IncA/C, which is seen in India and as well as in other geographical regions across the world. In contrast, in India, ST231 is the predominant ST and is exclusively associated with ColKP3 carrying blaOXA48-like. Continuous molecular surveillance of the prevailing clones and AMR profile of K. pneumoniae will help us to better understand the changing dynamics, to rework therapeutic management and intervention strategy for infection control.
Footnotes
Acknowledgments
We thank the team of the curators of the Institut Pasteur MLST system (Paris, France) for importing novel alleles, profiles, and/or isolates. We thank Ms. Catherine Truman, clinical pharmacist, Christian Medical College, Vellore, India, for her inputs in language correction of the article.
Disclosure Statement
This work is a part of the PhD doctoral thesis of Ms. Chaitra. S under the Tamil Nadu Dr. M.G.R. Medical University, Chennai, India. No competing financial interests exist.
Funding Information
This work did not receive any funding.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.