
Abstract
The development of carbapenem resistance in extraintestinal pathogenic Escherichia coli (ExPEC) has significant clinical implications, particularly in countries where second-line antimicrobials are not readily available, rendering treatments ineffective, and ExPEC infections untreatable. Thus, early detection of high-risk ExPEC lineages and raising awareness of the specific mechanisms underlying carbapenem resistance are mandatory for the selection of appropriate treatment options and the prevention of E. coli spread. This study aims to investigate the phenotypic and genotypic features of the first NDM-5 carbapenemase-producing ExPEC strain isolated from the blood of a patient admitted to the Maputo Central Hospital (MCH), in Mozambique. E. coli SSM100 isolate was identified by MALDI-TOF, it displayed high-level resistance to third generation cephalosporins, carbapenems, fluoroquinolones, and aminoglycosides, performing antimicrobial susceptibilities testing by VITEK 2 system. E. coli SSM100 isolate was classified through whole-genome sequencing as ST405-D-O102: H6, a globally distributed lineage associated with antimicrobial resistance, carrying the blaNDM-5 gene located on an F1:A1:B49 plasmid, coharboring blaCTX-M-15, blaTEM-1, aadA2, sul1, and dfrA12 genes. In addition, mutations in gyrA (S83L and D87N), parC (S80I and E84V), and parE (I529L) conferring fluoroquinolone resistance were also found. Moreover, SSM100 isolate carried 88 virulence genes, of which 28 are reported to be associated with UPEC. The emergence of NDM-5 carbapenemase in a pandemic ST405-D-O102:H6 clone in Mozambique is of great concern. Locations of extended-spectrum β-lactamase determinants and NDM-5 carbapenemase gene on IncF-plasmid can increase their spread reinforcing the need for antimicrobial surveillance and the urgent introduction of carbapenemase detection tests in diagnostic laboratories of the country.
Introduction
Infections caused by extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae are becoming more frequent worldwide. Treatment options are narrowing, with carbapenem agents, tigecycline, and colistin considered as the last-resort drugs1,2
During the last decades, the rising of carbapenem resistance in Enterobacteriaceae has dramatically increased, posing a major public health threat globally. 2
In recent years, the importance of extraintestinal pathogenic Escherichia coli (ExPEC) as pathogens capable of causing a wide spectrum of different infections has gained prominence, particularly following the emergence of multidrug resistance. 3 Notwithstanding that carbapenem resistance in Escherichia coli (E. coli) strains is still unreported in most countries, the emergence of ExPEC-producing carbapenemases, particularly New Delhi Metallo-β-lactamase (NDM), has posed a severe threat to public health worldwide, due to a significant reduction in patient's treatment options.4–6
This emergence has serious implications, particularly for low-income countries, such as Mozambique, where the national health surveillance and controls of antibiotic consumption is still poorly addressed, thus facilitating the dissemination of antimicrobial resistance.7,8
NDM-producing E. coli have been identified worldwide in several sequence types (STs), most represented by ST101, ST167, ST410, ST131, ST405, and ST648, where some are pandemic or/and emerging ExPEC clones, where they also are implicated as vehicles in driving the international spread of blaCTX-M-15.9,10 The NDM-1 and NDM-5 are the most detected variants in clinical isolates worldwide, with NDM-5 being more dangerous because of its increased resistance to carbapenems and high dissemination capacity.9–11 Besides, NDM-5 has been associated with different ExPEC lineages from many parts of the world, including several African countries, notably ST410, ST167 (South Africa), ST636 (Malawi), ST5018 (Egypt), and ST2659 (Algeria).9,12–16
NDM determinants are described to be carried by 19 different replicon types of promiscuous epidemic plasmids mainly IncX3, IncFII, or IncC. 6 These plasmids often coharbor other resistance determinants and are capable of disseminating among bacteria through horizontal gene transfer resulting in a spread of resistance to multiple classes of antibiotics. 6
To the best of our knowledge, there are still no reports of NDM carbapenemase resistance in E. coli in Mozambique. However, despite the paucity of the data, the high prevalence of ESBLs in the country was confirmed through recent studies conducted in the Manhiça District regarding on the prevalence of E. coli harboring blaCTX-M-15 gene from blood and urine cultures of hospitalized children and in Maputo area from colonized university students.17,18
In this study, we report a ST405 NDM-5-producing ExPEC isolated from blood in the Medicine Department of Maputo Central Hospital (MCH), which was investigated both phenotypically and genetically.
Methods
Bacterial identification and antimicrobial susceptibility testing
In June 2018, an E. coli isolate was detected from the blood of a patient admitted at the Medicine Department of the Maputo Central Hospital (MCH), which is the quaternary and referral hospital in Mozambique. Ethical approval was provided by the National Health Bioethics Committee (CNBS) of Mozambique (Ref 78/CNBS/2017). The blood sample was collected aseptically by venipuncture in aerobic flasks (Becton-Dickinson, Franklin Lakes, NJ) and transported to the Microbiology Laboratory of the Medicine Faculty of Eduardo Mondlane University (MLMF-UEM) for a 5 day culture in BACTEC 9050 instrument (Becton-Dickinson). Preliminary E. coli identification was performed by subculture on MacConkey, Chocolate and Blood agar plates at 37°C overnight, Gram staining followed by conventional biochemical tests (lactose fermentation and beta-glucuronidase, indole, and oxidase tests). The isolate was then sent to Sassari University (Italy) where the species was confirmed by mass spectrometry using a matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) (MALDI Biotyper; Bruker Daltonics, Inc.). Antimicrobial susceptibilities were performed using VITEK 2 compact system with Vitek2 AST- GN377 cards (bioMérieux, Marcy-l'Etoile, France) and interpreted following the European Committee on Antimicrobial Susceptibility Testing (EUCAST) recommendation, 2018.
Whole-genome sequencing and analysis
Genomic DNA was extracted using the Wizard® Genomic DNA Purification Kit (Promega Corporation), quantified using Nanodrop (Thermo Fisher Scientific) and sent for whole-genome sequencing (WGS) using Illumina NextSeq platform, at a 30x coverage (NGX Bio, Inc.). The short reads generated were assembled into contigs using de novo assembly, SPAdes 3.13.1 web-based tool. The resulting contigs in each assembly were classified as chromosomal or plasmid by using mlplasmids version 1.0 specifying “Escherichia coli” model. 19 Sequence data have been deposited in the EMBL-EBI database under the project accession number ERS4552076. The assembled genome was subjected to the online search tools ResFinder 3.2; MLST 2.0, PlasmidFinder 2.1 and pMLST 2.0, SerotypeFinder 2.0, FimTyper 1.0, and CHTyper 1.0 available at the Center for Genomic Epidemiology (CGE) (www.genomicepidemiology.org/). For E. coli Phylogroup typing the http://clermontyping.iame-research.center/ and https://nickp60.pythonanywhere.com/ web sites were used. The alignment between plasmid contigs and the reference plasmid pAMA1167-NDM-5 (CP024805.1) was performed using BLAST Ring Image Generator (BRIG, PMID: 21824423) using a threshold e-value of 0.01 for BLAST and displaying alignments with an identity of 90–100% and 70–90% in different colors. VirulenceFinder 2.0 was used to identify virulence genes.
In parallel, the recently published ASA 3 P analysis pipeline was used (https://github.com/oschwengers/asap).
In addition, other 28 virulence factors, typically of ExPEC (F1C fimbriae, hylA (AF037578), hylD (Y13891), sfaA (AL513382), sfaS (KY084254), afa (AF325672), fimH (JX847135), usp (AB027193), papA (NC_011750), papC (JX485631), papG (AF237474), papGII (AY212279), papGIII (AY212281), iucC (MK941173), iutA (JX466848), fyuA (KP657549), ireA (KU295572), traT (KY020407), malX (MH753045), cvaC (JN704082), ompT (HM210637), crl (X67207), sitA (KP657545), cdtB (NC_012944.1), KpsM (M57382), K1 (AY779018), neucK1 (NZ_JQFH01000102), and ibeA (AY248744), were searched manually using the Geneious R11 software.
Results
Phenotypic and genotypic analysis of ExPEC SSM100 isolate
In June 2018, strain SSM100, isolated from the blood of an 84-year-old patient admitted to the Medicine department of MCH, was identified as E. coli.
SSM100 E. coli isolate displayed multidrug resistance (MDR), characterized by high-level resistance to third generation cephalosporins, carbapenems, fluoroquinolones, and aminoglycosides, but remaining susceptible to tigecycline, colistin, and fosfomycin (Table 1).
Antimicrobial Susceptibility Pattern of Escherichia coli SSM100 Isolate
MIC, minimum inhibitor concentration; AMC, amoxicillin-clavulanic acid; TZP, piperacillin-tazobactam; CTX, cefotaxime, CAZ, ceftazidime; ETP, ertapenem; MEM, meropenem; AMK, amikacin; GEN, gentamicin; CIP, ciprofloxacin; TGC, tigecycline, FOF, fosfomycin; CST, colistin; SXT, trimethoprim-sulfamethoxazole; R, resistant; S, susceptible. MICs were calculated according to EUCAST recommendation.
WGS revealed that the genome size of isolate SSM100 was 5,560,355 kb with an average GC content of 50% and contained 5596 genes.
The in silico analysis typed the isolate as phylogroup D, serotype O102:H6, and fumC37_fimH29 clonotype, which belongs to the globally distributed clonal lineage ST405 based on multilocus sequence typing (MLST).
Three β-lactamases resistance genes, including blaTEM-1B, the ESBL blaCTX-M-15, and the carbapenemase blaNDM-5, were identified. In addition, SSM100 harbored other genes linked to aminoglycosides (aac(3)-IIa, aph(3′)-Ia aph(3′)-Ib, aph(6)-Id, and rmtB a 16S rRNA methylase conferring a high-level resistance to all aminoglycosides, and aadA2), chloramphenicol (catA1), sulfonamides (sul1 and sul2), tetracycline (tetB), and trimethoprim (dfrA12) resistance (Table 2). Chromosomal mutations associated with fluoroquinolone resistance were found in topoisomerase genes: two in gyrA (S83L and D87N), two in parC (S80I and E84V), and one in parE (I529L). Notably, plasmid-mediated quinolone resistance (PMQR) aac(6’)Ib-cr gene cassette and other qnr genes were not identified.
Distribution of Resistance Genes in Escherichia coli Strain SSM100
PlasmidFinder identified different replicon plasmids, including IncFIA, IncFIB, IncFII of pMLST type [F1:A1:B49], ColBS512, IncI1, IncI2, ΔIncQ1, and IncX4, all except for ColBS512 and ΔIncQ1, resided on plasmid contigs by the mlplasmids analysis.
When we looked at the localization of the resistance genes, we found that aadA2, dfrA12, sul1, rmtB, blaCTX-M-15, blaNDM-5, blaTEM-1B, and catA1 genes were located on plasmid contigs by mlplasmids analysis (Table 2). In particular, blaNDM-5 was located on the same plasmid contig_135, bracketed by two insertion sequences IS26 (partially assembled in the contig with 14 bases) containing a conserved region (bleMBL-trpF-tat), an ISCR1 transposase element, and a class I integron with aadA2 and the dfrA12 resistance genes (IS26-ΔIS30 -blaNDM-5-bleMBL-trpF-tat-ISCR1-sul1-qacEΔ1-aad2-orf-dfrA12-IntI1-IS26) (Fig. 1). BLAST search of this sequence showed 100% of identity with nucleotide sequences surrounding NDM-5 gene previously described in pAMA1167-NDM-5 (GenBank accession No CP024805) from an E. coli strain isolated in Denmark and other 17 plasmids, mainly from IncF type, found in 16 E. coli and one Klebsiella pneumoniae (Fig. 1).
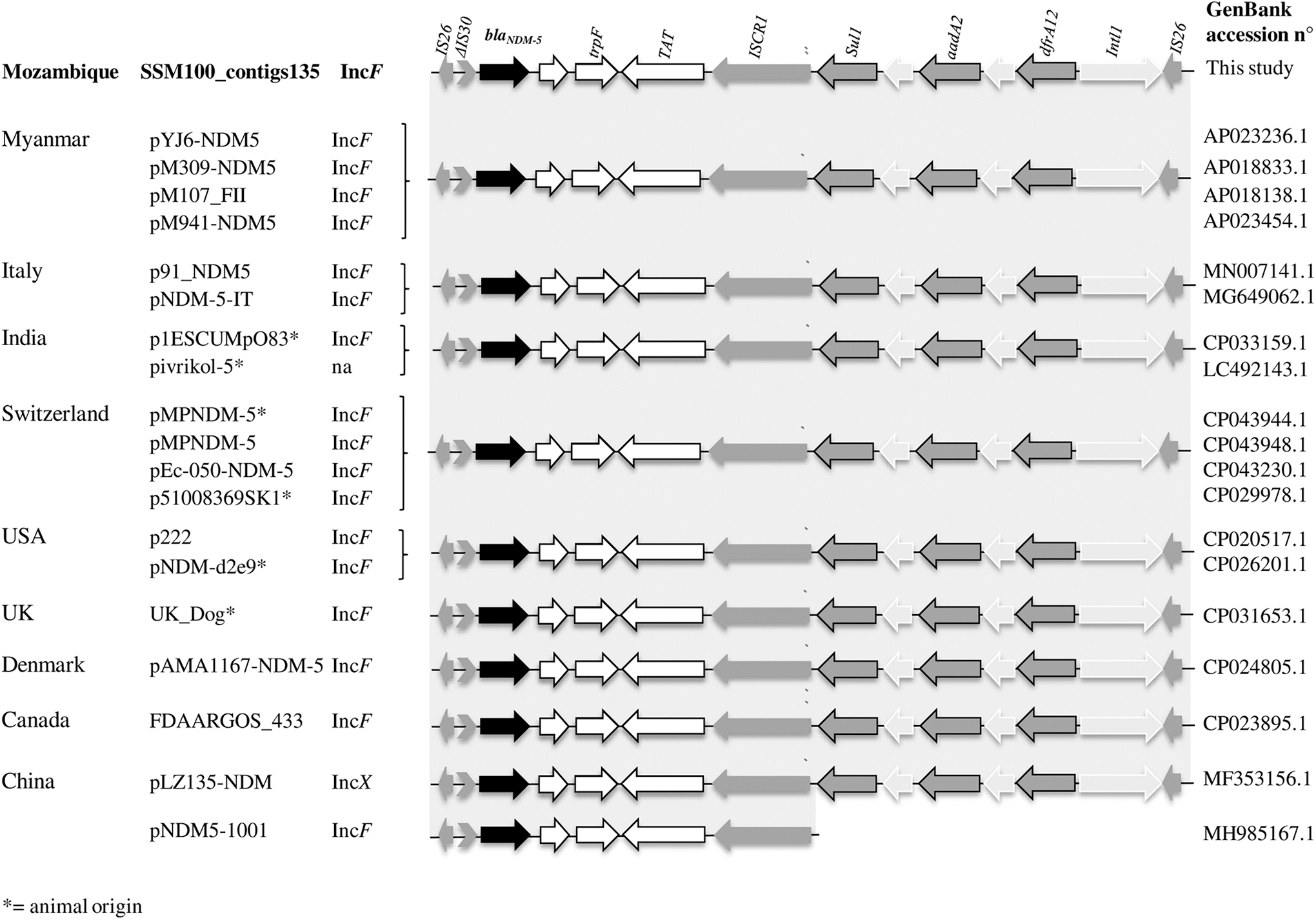
Schematic representation of the genetic structures surrounding blaNDM-5 gene here found and its alignment with plasmids of different countries showing 100% of coverage and identity by BLASTn (https://blast.ncbi.nlm.nih.gov/). The alignment with plasmid p1001-NDM5 from China carrying blaNDM-5, the conserved region and the ISCR1 transposase element (99.89% identity), and lacking the class I integron, is also shown. White arrows with black outline represent conserved genes, black is blaNMD-5 gene, gray in black outline arrows are resistant genes, and gray arrows are insertion sequences.
The blaCTX-M-15 and blaTEM-1B genes, assembled into different contigs, also showed 100% of identity with plasmid pAMA1167-NDM-5 (Fig. 2).
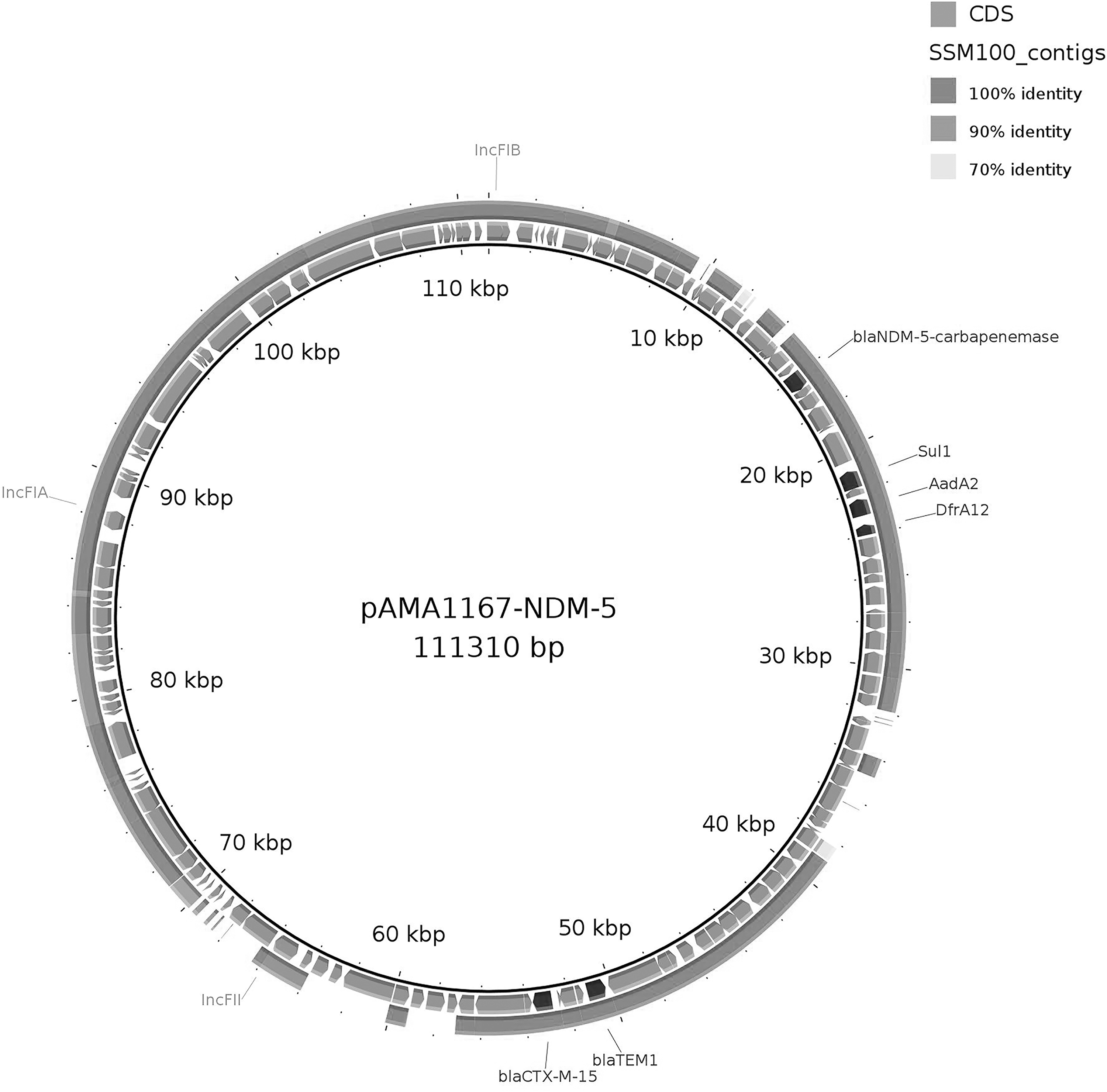
Comparative genetic map of Escherichia coli SSM100 genome contigs and plasmid pAMA1167-NDM-5 (CP024805.1) using BLAST Ring Image Generator (BRIG). Bars indicate the positions of antibiotic resistance genes and IncF replicons. The outer ring indicates the regions of nucleotide sequence identity of 70–100% in different shades of gray between E. coli SSM100 and plasmid pAMA1167-NDM-5.
Moreover, by using BRIG, we compared SSM100 contigs with the complete sequence of the pAMA1167-NDM-5 plasmid of pMLST type F1:A1:B49 and found that the SSM100 genome covered a significant part of the pAMA1167-NDM-5 plasmid backbone (coverage 71%, identity 99.99%) (Fig. 2).
Virulence-associated genes
A total of 88 virulence-associated genes (VAGs) were detected in the SM100 strain, 17 were identified by VirulenceFinder and a further 69 by ASA 3 P analysis, as shown in the Supplementary Tables S1 and S2. Two additional genes, crl and malX genes, encoding, respectively, for the sigma factor-binding protein Crl and the E. coli pathogenicity island-marker, were manually detected. The VAGs included 28 typically UPEC virulence genes for adhesins: fimABCDEFH, papAEFI, and iha; biofilm formation: crl and csgB; toxins: sat; iron uptake: iucABCD, iutA, and fyuA; protectins/serum resistance: traT, KpsE, and KpsM; outer membrane protein: ompA; outer membrane hemin receptor: chuA and yersiniabactin: ybtPQ.
Other identified VAGs were represented by eilA (involved in aggregation and colonization), air (Enteroaggregative immunoglobulin repeat protein), gad (Glutamate decarboxylase enabling survive to low pH), hra (Heat-resistant agglutinin), irp1 and irp2 (encoding for iron regulatory proteins), and terC (encoding for tellurium ion resistance protein) and others as listed in Supplementary Tables S1 and S2.
Discussion
ExPEC represent an important cause of severe community- and hospital-acquired infections, and it has been reported to be increasingly acquiring resistance to extensive classes of antimicrobials, including third generation cephalosporin and carbapenems. The emergence of the most recently described carbapenemase NDM constitutes a major therapeutic concern due to its high-level resistance to many available β-lactams, its rapid and ongoing evolution, and global dissemination.9,20
Infections caused by NDM-producer strains are associated with epidemic events and increased severity of the disease and mortality worldwide.9,11,21,22 In this study, we identified an ST405-D-O102:H6 ExPEC strain isolated from blood, coharboring blaNDM-5 and blaCTX-M-15 and displaying multidrug resistance to carbapenems, third-generation cephalosporins, aminoglycosides, and fluoroquinolones, which is epidemiologically linked to extraintestinal infections. 9 To our knowledge, this is the first description of an NDM carbapenemase-producing MDR ExPEC isolated in Mozambique. This raises concerns about the sustainability of the use of β-lactams as first-line antibiotics for the treatment of severe infection caused by Enterobacteriaceae species, common in the country especially among children, and it forces the use of last-line antibiotics that are often too expensive or not available, particularly in rural areas.23,24 Here, the finding of an MDR-NDM-5 carbapenemase-producing ExPEC suggested the use in clinical practice of last choice drugs colistin or tigecycline, although there are uncertainties regarding their efficacy. 25
The data retrieved from the genome analysis showed that our isolate harbored many known VAGs typically of ExPEC, of which 28 are reported to be associated with UPEC strains,26,27 making ST405-D-O102:H6 a straight ExPEC comparable with the pandemic high-risk UPEC clone O25b:H4-ST131. 28 Although the patient's clinical history was not available, the set of UPEC genes found in SSM100 strain suggested a urinary tract infection origin.
The virulence potential of our isolate is a complex issue, requiring further studies on the pathogenicity, including the assessment of pathogenic capability in vivo or in vitro and virulence level (LD50).
ST405-D-O102:H6 is an emerging urosepsis pathogen globally distributed, associated with ESBLs such as CTX-M-15 as found in this study, and increasingly associated with the presence of NDM variants, suggesting that this clonal lineage has the potential to play an important role in the transmission of the blaNDM genes.9,10,28 However, our finding of NDM-5 in ST405 clonal lineage is not common and reported only from few countries, including Italy, China, Spain, Myanmar, and, more recently, India and Japan.29–34
The blaNDM-5 determinant has been extensively reported in association with conjugative plasmids, in particular in ST405 clone, and it has been always associated with an IncF type plasmid rather than the globally disseminated IncX3-type, also described in other Enterobacteriaceae.35–37 Our WGS data analysis also indicated the blaNDM-5 gene in association with an IncF plasmid of pMLST type [F1:A1:B49], showing high identity with pAMA1167-NDM-5 plasmid spreading in Denmark (CP024805). Both plasmids contained the same conserved region (blaNDM-5-bleMBL-trpF-tat), the ISCR1 element, and class I integron with aadA2/dfrA12 resistance genes, also previously described in plasmids from different countries, including Italy, 38 China, 11 and others (Fig. 1), although none of them was from African countries. This conserved region (blaNDM-5-bleMBL-trpF-tat), which was previously labeled not to be lost during the NDM-5 transmission and the ISCR1 element seems to play an important role in assisting blaNDM-5 spread to other plasmids via rolling circle transposition. 11
Notwithstanding that carbapenems are not routinely used in antimicrobial therapies in Mozambique, the NDM-5 conserved sequence or the whole IncF NDM-5 plasmid or the ST405NDM5 ExPEC clone detected here, may have been imported to Mozambique from countries, where E. coli ST405 clone and NDM-5 plasmids are endemic such as China.11,28,30,35
The association of IncF plasmid carrying blaNDM-5 with the globally distributed ST405 clone is worrisome since this plasmid might serve as a reservoir for further dissemination of blaNDM–5 into other E. coli or other Enterobacteriaceae species, as already seen for IncX3 plasmid.11,35,37 An additional concern is the spread of blaNDM-5 in the community since previous reports described the isolation of ExPEC strains carrying NDM-5 on IncX3 and IncFII plasmids from animals, food samples, and environment.4,39–41
Conclusion
The finding in Mozambique of a pandemic MDR ST405-D-O102:H6 ExPEC fimH29 clonotype, carrying NDM-5 carbapenemase constitutes a great concern, posing a challenge for clinicians and a public health threat.
The threat of infections due to carbapenemase-producing ExPEC and the potential spread of blaCTXM-15 and carbapenemase NDM-5 gene via IncF plasmid prompts for a stricter antimicrobial surveillance and for an urgent introduction of carbapenemase detection tests in the diagnostic laboratories of the country.
Footnotes
Acknowledgments
The authors thank James R. Johnson and Brian Johnston from the University of Minnesota (USA) for providing positive controls of ExPEC and all the staff from Medicine department and Microbiology Laboratory of Maputo Central Hospital and Microbiology Laboratory of Medicine Faculty of Eduardo Mondlane University for their commitment and cooperation. BP is supported by Sardinian Region [POR-FSE 2014-2020-Asse Prioritario 3 “Istruzione e Formazione”-Obiettivo Tematico: 10, Priorità d'investimento: 10ii, Obiettivo Specifico: 10.5, Azione dell'Accordo di Partenariato 10.5.12-C.U.P. J86C18000270002].
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Mozambican Government through the Ministry of Science and Technology, Higher Education and Vocational Technical (HEST) and partially supported by the project “Training in Biodiversity and Biotechnology, AID 238 11096”, of the Italian Agency for International Cooperation - AICS.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.