
Abstract
Actinobacillus pleuropneumoniae, the etiological agent of porcine pleuropneumonia, is increasingly resistant to antibiotics. However, little is known about the mechanisms of antibiotic resistance in this pathogen. In this study, we experimentally evolved the reference strain of both A. pleuropneumoniae serovar 1 and serovar 7, the most prevalent serovars worldwide, to quinolone resistance by sequential exposure to subinhibitory concentrations of ciprofloxacin. The adaptive ciprofloxacin-resistant mutants of A. pleuropneumoniae serovar 1 and serovar 7 had a minimum inhibitory concentration (MIC) increment from 0.004 to 1 or 2 μg/mL, respectively. Adaptation to ciprofloxacin was shown to confer quinolone resistance with a 32- to 512-fold increase (serovars 1 and 7, respectively) as well as cross-resistance to ampicillin with an increased MIC by 16,384- and 64-fold (serovars 1 and 7, respectively). The genetic analysis of quinolone resistance-determining region mutations showed that substitutions occurred in gyrA (S83A) and parC (D84N) of serovar 1, and gyrA (D87N) of serovar 7. The ciprofloxacin-resistant mutants showed significantly reduced bacterial fitness. The mutants also showed changes in efflux ability and biofilm formation. Notably, the transcription and secretion levels of Apx toxins were dramatically reduced in ciprofloxacin-resistant mutants compared with their wild-type strains. Altogether, these results demonstrated marked phenotypic changes in ciprofloxacin-resistant mutants of A. pleuropneumoniae. The results stress the need for further studies on the impact of both the genotypic and phenotypic characteristics of A. pleuropneumoniae following exposure to subinhibitory concentrations of antibiotics.
Introduction
A
At present, the use of antibiotics and vaccines are the most effective approaches to reduce the severity of clinical symptoms and mortality of A. pleuropneumoniae infection. 1 Although A. pleuropneumoniae is susceptible to the majority of antibiotics, a dramatic increase in resistance to a wide range of antibiotics such as quinolones, tetracyclines, β-lactams, and sulfonamides has been reported worldwide.6–9 Therefore, it is imperative to understand the mechanisms of antibiotic resistance to ensure the appropriate and targeted use of antibiotics in the control of A. pleuropneumoniae infections. Currently, resistance genes such as tet(B) or tet(H) for tetracycline resistance, sul2 for sulfisoxazole resistance, blaROB-1 for ampicillin resistance, and the multiple resistance genes ICEAplChn1 have been identified by whole-genome sequence or PCR.7,9–11 In addition to the plasmid-mediated resistance, chromosomal-mediated resistance due to mutations in the quinolone resistance-determining region (QRDR) and active effluxes have also been observed to contribute to enrofloxacin resistance. 8 Despite this increasing trend of resistance, the mechanisms of A. pleuropneumoniae resistance to various antibiotics remain poorly understood.
Recent evidence has suggested that exposure of bacteria to subinhibitory antibiotic concentrations is likely to have an important role in the emergence of antibiotic resistance. 12 Mutants with a raised minimum inhibitory concentration (MIC) that have been generated by exposure to subinhibitory antibiotic concentrations have been used to study emergence of antibiotic resistance and the impact of specific mutation on bacterial physiology and pathogenesis. For example, the emergence of fluoroquinolone resistance in Pseudomonas aeruginosa was evaluated by exposure to subinhibitory levels of ciprofloxacin, which provided new insights into the changes of QRDR mutations, cross-resistance, bacterial fitness, and biofilm populations in ciprofloxacin-resistant mutants.13–15 These studies greatly improved understanding of the emergence of fluoroquinolone resistance in gram-negative bacteria.
Previous studies have been reported that fluoroquinolones (especially ciprofloxacin and enrofloxacin) were very effective against A. pleuropneumoniae.6,16,17 Nevertheless, increasing levels of acquired resistance to fluoroquinolones have been reported in recent years.8,9,18 The occurrence of resistance and trends in fluoroquinolone resistance should be monitored carefully for A. pleuropneumoniae. To better understand the mechanisms of chromosomal-mediated fluoroquinolone resistance in A. pleuropneumoniae, mutants of both the serovar 1 and serovar 7 reference strains with an increased MIC to ciprofloxacin were generated by exposure to subinhibitory concentrations of ciprofloxacin. The MICs of quinolones and other antibiotics for the resistant mutants were compared with that of the wild-type (WT) and control strains. The characteristics of the resistant mutants were evaluated in terms of growth fitness, QRDR mutations, efflux ability, biofilm formation, and Apx toxin secretion. The transcriptional levels of the genes involved in the physiological characters were also tested by real-time quantitative reverse transcription PCR (qRT-PCR). These results will be helpful for understanding the impact of fluoroquinolone resistance at the chromosomal level in A. pleuropneumoniae.
Materials and Methods
This study was approved by the Institutional Review Board (IRB) of Institute of Animal Husbandry and Veterinary Medicine, Beijing Academy of Agriculture and Forestry Sciences.
Bacterial strain and culture conditions
A. pleuropneumoniae reference strains 4074 (serovar 1) and WF83 (serovar 7) were routinely grown in tryptic soy broth (TSB; Difco, BD) or on tryptic soy agar (TSA; Difco, BD), with both media supplemented with 10 μg/mL of nicotinamide adenine dinucleotide (NAD; Sigma), at 37°C unless otherwise mentioned.
MIC testing
The MICs of various antibiotics for A. pleuropneumoniae were determined using a broth microdilution method, as described previously. 19 Briefly, the antibiotic solution was freshly prepared and diluted before use. A 20 μL aliquot of the diluted antibiotic solution was added into the 180 μL TSB+NAD medium in the first column of the 96-well plate and a serial 2-fold dilution performed across the plate. A 0.5 mL aliquot of overnight bacterial culture of each A. pleuropneumoniae strain was reinoculated into 4.5 mL TSB+NAD medium and the broth incubated for 3 hr with shaking at 37°C. The optical density (OD) of the incubated broths was adjusted to be OD600 = 1.0 and then diluted 200-fold. A 5 μL volume of the diluted culture was then added per well (meaning a final concentration of ∼2.5 × 104 colony-forming unit [CFU] per well). The inoculated plates were sealed and incubated at 37°C for 22 hr. The MICs were recorded as the lowest concentration of antibiotic that completely inhibited the bacterial growth, as judged by visual examination, in the wells. The microdilution assay was carried out three times in duplicate.
Induction of ciprofloxacin resistance by exposure to serially increased concentration of ciprofloxacin
Following the determination of ciprofloxacin MIC (0.004 μg/mL) for both A. pleuropneumoniae strains, mutants were generated by exposure to subinhibitory concentrations of ciprofloxacin in sequential passages as described previously 20 with some modifications. Briefly, A. pleuropneumoniae WT strains 4074 (Ap1-WT) and WF83 (Ap7-WT) were grown overnight and reinoculated into the fresh TSB+NAD medium at 1:50 dilution, respectively. The diluted cultures were grown by shaking for 3 hr before inoculation of the microplate. Selection of ciprofloxacin-resistant mutants was performed in a sterile 96-well microplate (Costar, Corning, NY) in triplicate for each ciprofloxacin concentration. In the first passage, 0.9 mL of serial twofold dilutions of ciprofloxacin solutions at concentrations of 8 × MIC, 4 × MIC, 2 × MIC, 1 × MIC, 0.5 × MIC, 0.25 × MIC was inoculated with 0.1 mL of broth culture containing ∼2.5 × 106 CFU/mL. For each passage, a ciprofloxacin-free culture control was included. The microplates were incubated at 37°C for 24 hr. The cultures grown at the highest concentration of ciprofloxacin were chosen for the next passage, after subculture in ciprofloxacin-free TSB+NAD broth medium. The ciprofloxacin concentration was gradually increased, and this process was repeated for 50 generations until no growth was observed in the medium. After the final passage, the cultures were plated on the TSA+NAD medium and a single colony was cultured for MIC determination. The high-level ciprofloxacin-resistant mutants, named Ap1-CIP and Ap7-CIP, were stored in 20% glycerol at −80°C for further use. Simultaneously, the control strains Ap1-Con and Ap7-Con were also generated by serially passaging in ciprofloxacin-free TSB+NAD medium. To determine the stability of the MICs, all the evolved strains were serially passaged for five generations in medium without addition of ciprofloxacin.
PCR and sequencing analysis
To determine the mutations of quinolone target enzymes in the ciprofloxacin-resistant mutants of A. pleuropneumoniae, the genes encoding DNA gyrase (gyrA and gyrB) and topoisomerase IV (parC and parE) were amplified from the genomic DNA of Ap1-WT, Ap1-Con, Ap1-CIP, Ap7-WT, Ap7-Con, and Ap7-CIP by PCR, respectively. Primers used for amplifying the four genes are listed in Supplementary Table S1. The PCR assays were performed in a 50 μL volume using PfuUltra® High-Fidelity DNA Polymerase (Stratagene) as described previously. 21 PCR products were purified by QIAquick PCR Purification Kit (Qiagen) and cloned using the pEASY-Blunt Zero Cloning Kit (TransGen Biotech). Four clones of each gene fragment were chosen for Sanger sequencing using ABI 3730xl DNA analyzer (Sangon Biotech). The sequence results were analyzed using Nucleotide BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and compared with the reference genomes of A. pleuropneumoniae serovar 1 strain 4074 (NZ_CP029003.1) and serovar 7 strain AP76 (NC_010939.1).
Growth curve
To determine the bacterial fitness of ciprofloxacin-resistant mutants, the growth rates of A. pleuropneumoniae were measured in TSB+NAD broth using a Bioscreen C Analyzer (Oy Growth Curves Ab Ltd, Helsinki, Finland). The concentration of an overnight culture of the strains in TSB+NAD broth was adjusted to be OD600 = 1.0. Then, the adjusted cultures were 100-fold diluted and 100 μL inoculated per well in a 100-well plate. The cultures were grown at 37°C for 26 hr, and the OD600 measurements were taken every 1 hr with shaking. The growth curve of each strain was drawn using the OD600 measurements. The growth rates were carried out three times in duplicate.
Accumulation assay
Accumulation of ciprofloxacin in A. pleuropneumoniae was determined as described previously 22 with minor modifications. Briefly, bacteria were grown in TSB+NAD medium at 37°C overnight with shaking and harvested by centrifugation. The bacterial pellet was washed, resuspended, and adjusted to an OD600 of 1.0 (∼109 CFU/mL). The cell suspension was incubated for 10 min at 37°C, and ciprofloxacin was added to a final concentration of 10 μg/mL. After addition of ciprofloxacin, 0.5 mL samples were removed every 1.5 min. At 6 min after ciprofloxacin addition, the efflux pump inhibitor carbonyl cyanide 3-chlorophenylhydrazone (CCCP) was added to one-half of the suspension at a final concentration of 6 μg/mL and the other half was used as control. Collections from both fractions continued every 1.5 min until 21 min. All the collected samples were immediately subjected to measurement for ciprofloxacin accumulation. The fluorescence of the supernatants was measured with a Synergy H1 microplate reader (BioTek) at excitation and emission wavelengths of 279 and 447 nm, respectively. The concentrations of ciprofloxacin in the supernatants were calculated by comparison with a standard curve of ciprofloxacin in 0.1 M glycine hydrochloride (pH 3.0). The accumulation assay was carried out three times in duplicate.
Biofilm assay
The effects of ciprofloxacin-induced resistance on biofilm formation were determined by the microtiter plate biofilm assay as described previously 23 with some modifications. Briefly, all the A. pleuropneumoniae strains were grown in brain heart infusion broth (Difco, BD) with addition of NAD. The values of all the overnight cultures were adjusted to be OD600 = 1.0. Then, the adjusted cultures were 100-fold diluted and 100 μL inoculated per well in sterile 96-well microtiter plates (Corning). Following incubation for 24 hr or 48 hr at 37°C, each well was washed three times with PBS and 100 μL of methanol was added to fix the remaining attached bacteria. After air-drying, the wells were stained with 100 μL per well of crystal violet (0.1%) for 10 min at room temperature. After removing the crystal violet solution, the wells were washed with water and dried. Finally, 100 μL per well of glacial acetic acid (33%, v/v) was added, and the OD590 value per well was measured with a Synergy H1 microplate reader (BioTek). The biofilm assay was performed three times in triplicate.
Production of recombinant ApxIA and ApxIIA and generation of polyclonal antisera
The histidine-tagged recombinant ApxIA (rApxIA) and recombinant ApxIIA (rApxIIA) were produced in Escherichia coli BL21 (DE3) using pET-30a vector (Novagen), respectively. Briefly, a 1,044-bp fragment covering the 348-aa N-terminal segment of ApxIA or ApxIIA was PCR amplified from A. pleuropneumoniae reference strain 4074 (serovar 1) using the primers listed in Supplementary Table S2. The amplified product and the vector pET-30a were digested with NdeI and XhoI and ligated onto each other. After cloning and successful expression, the rApxIA and rApxIIA were purified by the AKTA Purifier 100 System (GE Healthcare, Bucks, United Kingdom) using a HisTrap FF affinity chromatography column. The highly purified rApxIA and rApxIIA were used for production of rabbit polyclonal antisera. The prepared rabbit polyclonal antisera against rApxIA and rApxIIA were used for the following Western blot analysis.
Apx toxin secretion assay
To determine the effects of ciprofloxacin-induced resistance on secretion of Apx toxin, the secretion level of ApxI and ApxII toxins in supernatants of A. pleuropneumoniae cultures was tested in Ap1-WT, Ap1-Con, Ap1-CIP, Ap7-WT, Ap7-Con, and Ap7-CIP as described previously. 24 Briefly, all strains were cultured in pleuropneumonia-like organisms (PPLO) broth (Difco, BD) supplemented with 10 μg/mL NAD. The overnight cultures were inoculated into fresh PPLO+NAD medium with the addition of CaCl2 at a final concentration of 25 mM. After shaking incubation at 37°C for 5 hr, the values of all the cultures were adjusted to be OD600 = 1.0. Then, 10 mL of the adjusted culture of each strain was harvested by centrifuging at 10,000 × g for 10 min at 4°C. The supernatants were concentrated 20-fold using Amicon Ultra-15 10 kDa filtration centrifugal tube (Millipore). The Apx toxins in the concentrated supernatants of each strain were examined by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot using the anti-rApxI and anti-rApxII rabbit polyclonal antisera with 1:200 dilution following the standard procedures.
Real-time quantitative reverse transcription PCR
Genes involved in efflux pump, biofilm formation, and Apx toxin expression and secretion were selected for analysis at the transcriptional level by qRT-PCR. The selected genes and primers used for qRT-PCR are listed in Supplementary Table S3. Total RNA was extracted from strains of Ap1-WT, Ap1-Con, Ap1-CIP, Ap7-WT, Ap7-Con, and Ap7-CIP. The qRT-PCR was performed as described previously. 25 The relative transcription level of each gene was determined by normalization to that of the 16S rRNA gene using the 2−ΔΔCt method.
Statistical analysis
Statistical analysis was performed using a one-way analysis of variance. Duncan's multiple range test was used to compare the differences among the treatment groups. A p-value of <0.05 was taken to indicate statistical significance.
Results
Induction of ciprofloxacin-resistant A. pleuropneumoniae by exposure to sublethal ciprofloxacin
Following imposition of selective pressure by serial transfers of A. pleuropneumoniae in increasing concentrations of ciprofloxacin, the strains Ap1-CIP and Ap7-CIP were able to increase their resistance to ciprofloxacin by 256- and 512-fold (MICs from 0.004 to 1 or 2 μg/mL) compared with the WT strains Ap1-WT and Ap7-WT, respectively. In comparison, the control strains Ap1-Con and Ap7-Con passaged in ciprofloxacin-free TSB+NAD medium simultaneously only increased twofold (MICs from 0.004 to 0.008 μg/mL) under the same experimental conditions (Table 1). The strains Ap1-CIP and Ap7-CIP were stable in terms of the MIC to ciprofloxacin after five serial transfers in ciprofloxacin-free TSB+NAD medium and after reculturing from −80°C storage.
Susceptibilities of Wild-Type Strains, Control Strains, and Ciprofloxacin-Resistant Mutants of Actinobacillus pleuropneumoniae Strains 4074 (Serovar 1) and WF83 (Serovar 7) to Different Antimicrobials a
Ap1-WT = A. pleuropneumoniae serovar 1 strain 4074; Ap1-Con = A. pleuropneumoniae serovar 1 strain 4074 passaged in the absence of ciprofloxacin; Ap1-CIP = A. pleuropneumoniae serovar 1 strain 4074 passaged in the presence of increasing subinhibitory concentrations of ciprofloxacin; Ap7-WT = A. pleuropneumoniae serovar 7 strain WF83; Ap7-Con = A. pleuropneumoniae serovar 7 strain WF83 passaged in the absence of ciprofloxacin; Ap7-CIP = A. pleuropneumoniae serovar 7 strain WF83 passaged in the presence of increasing subinhibitory concentrations of ciprofloxacin.
MIC fold change for Ap1-Con compared with Ap1-WT.
MIC fold change for Ap1-CIP compared with Ap1-WT.
MIC fold change for Ap7-Con compared with Ap7-WT.
MIC fold change for Ap7-CIP compared with Ap7-WT.
CIP, ciprofloxacin; MIC, minimum inhibitory concentration; WT, wild type.
MIC determination to different antibiotics
The MIC values of the strains Ap1-WT, Ap1-Con, Ap1-CIP, Ap7-WT, Ap7-Con, and Ap7-CIP to 17 antimicrobial agents belonging to different antibiotic classes were determined using a broth microdilution assay (Table 1). Compared with the WT strains, the MIC values for Ap1-CIP and Ap7-CIP to quinolones including ciprofloxacin, norfloxacin, enrofloxacin, and nalidixic acid increased 32- to 512-fold, whereas the control strains Ap1-Con and Ap7-Con only changed by 0.5- to 2-fold. Remarkably, the MIC values of Ap1-CIP and Ap7-CIP to ampicillin increased 16,384- and 64-fold compared with the WT strains, respectively. In contrast, strains Ap1-CIP and Ap7-CIP showed a 16- and 4-fold reduction in the MIC value for colistin compared with Ap1-WT and Ap7-WT, respectively. Furthermore, inconsistent results for Ap1-CIP and Ap7-CIP were also observed for MIC values to trimethoprim, a 16-fold decrease and an 8-fold increase compared with the WT strains, respectively.
Mutations in quinolone target genes
Compared with the WT strains Ap1-WT and Ap7-WT, all the amino acid substitutions in gyrA, gyrB, parC, and parE of strains Ap1-Con, Ap1-CIP, Ap7-Con, and Ap7-CIP are listed in Table 2. The most mutations were observed in Ap1-CIP with 8, 1, 20, and 8 site substitutions in gyrA, gyrB, parC, and parE, respectively. Sequence analysis of the QRDRs revealed double mutations in Ap1-CIP (gyrA S83A, parC D84N) and single mutation in Ap7-CIP (gyrA D87N), which have been previously reported to be responsible for quinolone resistance.26,27 Notably, the 8 continuous amino acids (741–748) at the terminal of parC were substituted by the 7 amino acids (741–747) in all four strains. No mutations were observed in strains of Ap1-Con and Ap7-Con except 7 amino acid substitutions at the terminal of parC.
Site Mutations of Quinolone Target Genes in Actinobacillus pleuropneumoniae Control Strains and Ciprofloxacin-Resistant Mutants
The one-letter abbreviation nomenclature was used to represent amino acids. Substitutions were noted as follows: X#Y, where X represents the wild-type amino acid, # the codon number, and Y the substituted amino acid.
The insertional mutation between the two adjacent positions.
The eight amino acids (741–748) at the terminal of parC were substituted by the seven amino acids (741–747).
Ciprofloxacin-resistant mutants and bacterial fitness
To compare the bacterial fitness of the ciprofloxacin-resistant mutants with the WT parent strains, the growth curves of the strains Ap1-WT, Ap1-Con, Ap1-CIP, Ap7-WT, Ap7-Con, and Ap7-CIP were measured under the same drug-free growth conditions (Fig. 1). Compared with the WT strains Ap1-WT and Ap7-WT, the ciprofloxacin-resistant mutants Ap1-CIP and Ap7-CIP had significantly lower OD at the early stationary phase. However, the control strains, Ap1-Con and Ap7-Con, showed higher OD at this same stage than the WT strains. The results indicated that the ciprofloxacin-resistant mutants had reduced the fitness while the control strains that had been subcultured in the absence of ciprofloxacin had increased bacterial fitness.
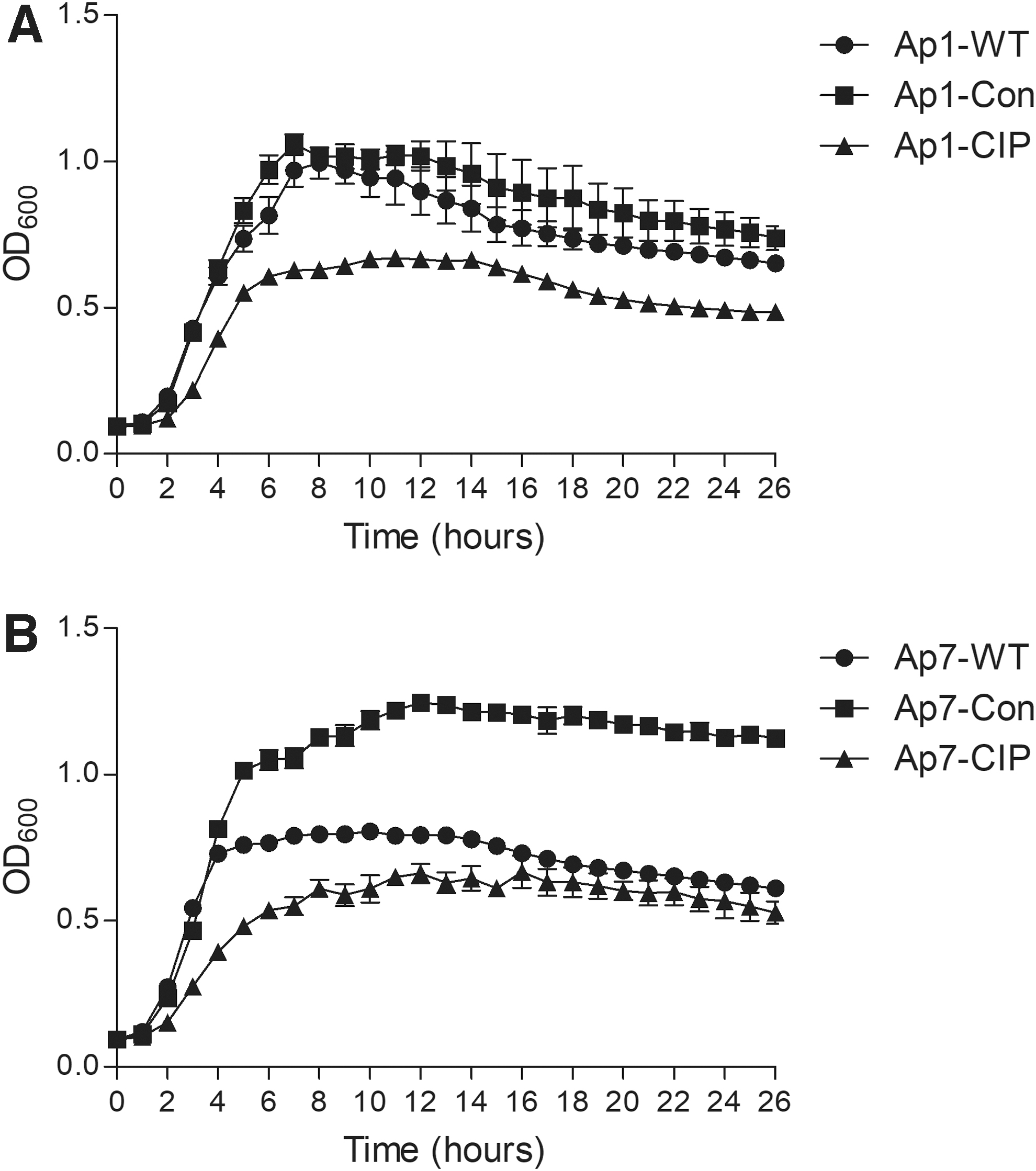
Growth curves of Actinobacillus pleuropneumoniae WT, control strains, and ciprofloxacin-resistant mutants.
Efflux pump contributes to ciprofloxacin resistance
To determine efflux-mediated resistance in ciprofloxacin-resistant mutants, the efflux pump inhibitor CCCP was used to evaluate the ciprofloxacin accumulation in bacterial cells. Compared with the slight ciprofloxacin concentration increase in Ap1-Con (Fig. 2A) and Ap7-Con (Fig. 2C), there was a very rapid and dramatic increase by three- to fourfold in the accumulation levels of ciprofloxacin in Ap1-CIP (Fig. 2B) and Ap7-CIP (Fig. 2D) at the steady state following the addition of CCCP. To further evaluate the roles of efflux pump in ciprofloxacin resistance, the transcriptional levels of two deduced efflux pump system components in A. pleuropneumoniae were tested by qRT-PCR. The fold changes of the transcriptional levels between the strains are listed in Table 3. Compared with the WT and control strains, the ciprofloxacin-resistant mutants showed that the second AcrR-AcrAB system components (acrR2, acrA2, acrB2) were upregulated while the first AcrR-AcrAB system components (acrR1, acrA1, acrB1) were downregulated. The transcriptional levels of tolC1 and tolC2 in the ciprofloxacin-resistant mutants were downregulated. These results suggested that an efflux pump was involved in the evolved ciprofloxacin resistance of A. pleuropneumoniae.
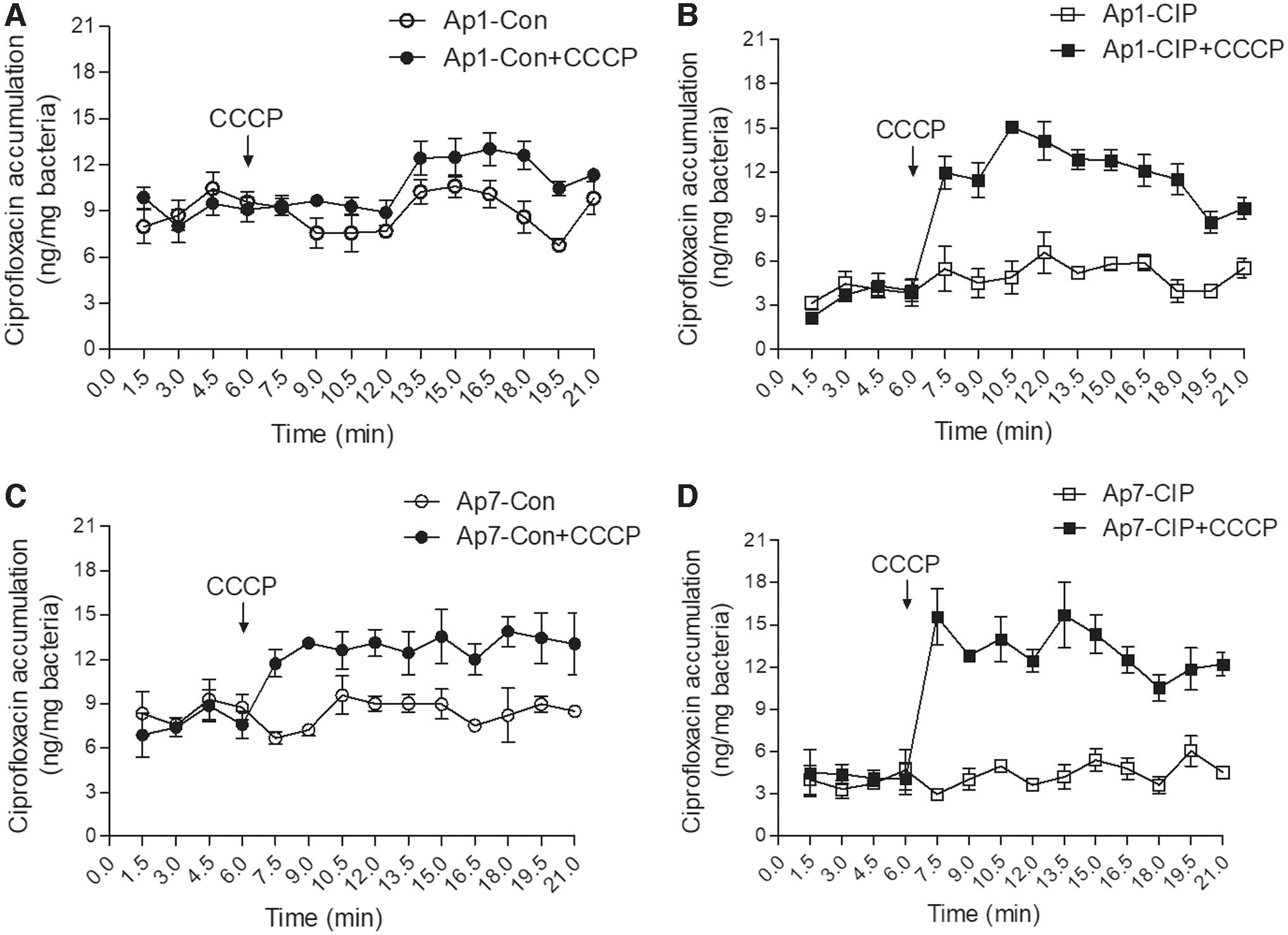
Accumulation of ciprofloxacin in Actinobacillus pleuropneumoniae control strains and ciprofloxacin-resistant mutants.
Fold Change of the Deduced Efflux Pump System Components of Wild-Type Strains, Control Strains, and Ciprofloxacin-Resistant Mutants in Actinobacillus pleuropneumoniae
The fold change was calculated from the 2−ΔΔCt value of real-time quantitative reverse transcription PCR.
The reference genomes of A. pleuropneumoniae serovar 1 strain 4074 (NZ_CP029003.1) and serovar 7 strain AP76 (NC_010939.1).
The two deduced efflux pump systems were designated based on the operon structure and similarity with other efflux pumps in gram-negative bacteria.
The tolC1 and tolC2 genes were from Li et al. (2016). 19
Ciprofloxacin-resistant mutants and biofilm formation
As biofilm production has been reported to be associated with antibiotic resistance,
28
the ability of the strains Ap1-WT, Ap1-Con, Ap1-CIP, Ap7-WT, Ap7-Con, and Ap7-CIP to form biofilm was investigated using crystal violet staining. In Ap1-CIP, the amount of biofilm was significantly increased compared with Ap1-WT and Ap1-Con at 24 and 48 hr of incubation (Fig. 3A). However, the biofilms of Ap7-CIP and Ap7-Con were significantly reduced compared with that of Ap7-WT at 24 and 48 hr of incubation (Fig. 3B). The transcriptional analysis on the poly-β-1,6-N-acetyl-

Biofilm formation and PGA biosynthetic genes expression level in Actinobacillus pleuropneumoniae.
Ciprofloxacin-resistant mutants and Apx toxin secretion
To determine the ability of the ciprofloxacin-resistant mutants to secret Apx toxins, the secretion levels of ApxI and ApxII toxins in the supernatants of bacterial cultures were tested in all six strains. In contrast to the similar secretion level of ApxI toxin in Ap1-WT and Ap1-Con, no visible band of ApxI toxin was detected in Ap1-CIP (Fig. 4A). The secretion level of ApxII toxin was also dramatically decreased in the strains Ap1-CIP and Ap7-CIP compared with that of the WT and control strains (Fig. 4A). Accordingly, compared with the WT strain Ap1-WT, the transcriptional levels on the activation and structural genes of apxIC, apxIA, apxIIC, apxIIA, and apxIV in Ap1-CIP were dramatically decreased by 245-, 819-, 7-, 25-, and 4-fold, respectively (Fig. 4B). Compared with the WT strain Ap7-WT, the transcriptional levels of apxIIC, apxIIA, and apxIV in Ap7-CIP were also dramatically decreased by 7-, 11-, and 9-fold, respectively (Fig. 4C). The transcriptional levels on the secretion genes of apxIB and apxID were inconsistent between Ap1-CIP (four- and onefold increase) and Ap7-CIP (four- and fivefold decrease) compared with their WT strains (Fig. 4B, C). The results indicated that ciprofloxacin-resistant mutants of A. pleuropneumoniae had reduced transcription and secretion of Apx toxins.
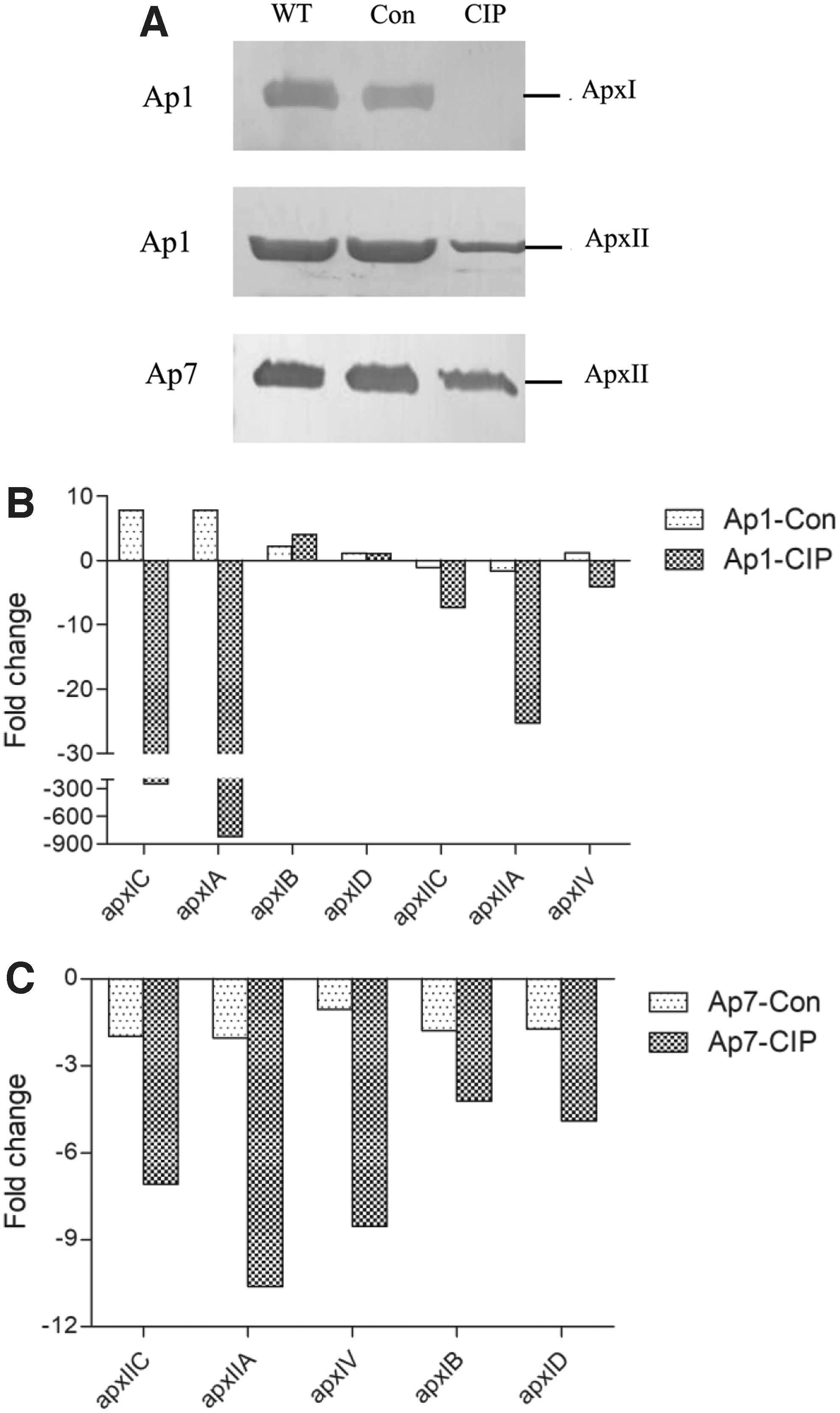
Secretion and expression levels of Apx toxins in Actinobacillus pleuropneumoniae.
Discussion
Bacterial pathogens exposed to subinhibitory concentrations of drugs frequently have an important role in the emergence of antibiotic resistance. 12 In this study, two reference strains that represented two commonly isolated serovars of A. pleuropneumoniae rapidly developed quinolone resistance following the addition of elevated concentrations of ciprofloxacin. The initially ciprofloxacin-susceptible A. pleuropneumoniae strains evolved and developed increased MICs for ciprofloxacin (256- to 512-fold increases) with sequential passages. Rapid development of high-level ciprofloxacin resistance has also been observed in other pathogens.13,15,30 These results indicate that exposure to subinhibitory antibiotic concentrations leads to emergence of bacterial drug resistance by enriching the pre-existing resistant bacteria and/or by selecting for de novo resistance, a process which endangers human and animal health. 12
Our finding that ciprofloxacin-induced resistant mutants showed increased MIC values for other quinolones such as norfloxacin, enrofloxacin, and nalidixic acid is an expected finding. However, the MIC increments of 16,384-fold (4074-serovar 1) and 64-fold (WF83-serovar 7) that we observed for beta-lactam antibiotics in the evolved ciprofloxacin-resistant mutants were a surprising finding. A similar change in MIC values for beta-lactam antibiotics (specifically ceftazidime and meropenem) was also observed in P. aeruginosa during exposure to sublethal concentrations of ciprofloxacin. 15 As the targets for quinolones and beta-lactams are quite different, it is possible that common resistance mechanisms, for example, efflux upregulation or altering the permeability of the cell membrane, may be involved in this kind of cross-resistance.31,32 The current study also showed an MIC reduction in the ciprofloxacin-resistant mutants for colistin compared with WT strains. Mutations to genes encoding lipopolysaccharide-modifying enzymes, particularly the two-component systems PmrAB and PhoPQ, and the MgrB regulator are known to play a role in colistin susceptibility. 33 It is possible that these types of mutations may have occurred during the serial passages and resulted in the reduced colistin MICs seen in this study. Furthermore, inconsistent changes to MIC values to trimethoprim for Ap1-CIP and Ap7-CIP may be also due to mutations in the encoding gene of dihydrofolate reductase. 34 It is important to note that most of the antibiotics we tested showed minimal MIC changes following ciprofloxacin-induced resistance (Table 1). The mechanisms underpinning the cross-resistance need to be further investigated.
A combination of mutations within gyrA, gyrB, parC, and parE has been found to be common in fluoroquinolone-resistant gram-negative bacteria. 35 In this study, multiple mutations in genes of gyrA, parC, and parE were detected in the evolved ciprofloxacin-resistant mutants Ap1-CIP and Ap7-CIP (Table 2). Mutations in the QRDR of these genes, resulting in amino acid substitutions and target protein structure alterations, lead to quinolone resistance. 35 QRDR analysis demonstrated that gyrA codon 83 or 87 and parC codon 80 or 84 (E. coli numbering) are the mostly common mutations.26,27,36,37 Enrofloxacin-resistant isolates in A. pleuropneumoniae were found to carry at least one mutation in the QRDR of gyrA codon 83 or 87. 8 Similar results were also observed in this study with double mutations in Ap1-CIP (gyrA S83A, parC D84N) and single mutation in Ap7-CIP (gyrA D87N).
Previous studies showed that fluoroquinolone-resistant mutations reduced the bacterial fitness in E. coli and Streptococcus pneumoniae, and the mutations in fluoroquinolone target genes were responsible for the fitness costs.35,38,39 Similarly, our results also indicated that ciprofloxacin-resistant mutants of A. pleuropneumoniae had decreased fitness. Interestingly, the growth curve analysis showed that Ap1-CIP mutant, which had double mutations (gyrA S83A, parC D84N), showed a great loss of fitness than the Ap7-CIP mutant, which had single mutation (gyrA D87N) (Fig. 1).
The ability to form biofilm is one of the major mechanisms of antibiotic resistance used by bacteria. 28 Previous studies have demonstrated that exposure to subinhibitory concentrations of antibiotics can enhance biofilm formation.12,28 However, a recent study has shown that different bacterial species and antimicrobial agents displayed a diverse correlation between biofilm formation and antibiotic resistance. 40 Our study has found a similar situation in terms of the ability to form biofilm. Following exposure to subinhibitory concentrations of ciprofloxacin, the serovar 1 ciprofloxacin-resistant mutant showed enhanced biofilm formation (compared with the parent strain) while the serovar 7 ciprofloxacin-resistant mutant showed reduced biofilm formation (compared with the parent strain). The pgaABCD operon, the global gene regulator H-NS, and the sigma factor σE are involved in biofilm formation in A. pleuropneumoniae. 41 It is possible that differences in these genes in the parent strains or possibly changes during the serial passage may explain the difference in the ability of the mutants to produce biofilm. We compared the sequence of these genes in the reference genomes of A. pleuropneumoniae serovar 1 strain 4074 (NZ_CP029003.1) and serovar 7 strain AP76 (NC_010939.1) but were unable to see any obvious differences between these two strains. The relationship between biofilm formation and quinolone resistance is not clear in A. pleuropneumoniae and further studies are required.
Efflux upregulation was reported to be one of major mechanisms of quinolone resistance. 35 Based on the results using the efflux pump inhibitor CCCP in this study, we have demonstrated that an efflux pump was involved in the quinolone resistance in A. pleuropneumoniae. This supports the findings of a prior study on A. pleuropneumoniae using Phe-Arg-β-naphthylamide. 8 Previous reports have shown that the common efflux pump AcrAB-TolC was involved in fluoroquinolone resistance in Salmonella enterica and E. coli. 35 To date, no comprehensive study on efflux pumps and A. pleuropneumoniae has been reported. Our transcriptional analysis indicated that one deduced AcrR-AcrAB system component was upregulated in the ciprofloxacin-resistant mutants. In a recent study, two outer membrane channel components TolC1 and TolC2 were identified, and TolC1 was defined to be involved in antibiotic resistance and biofilm formation in A. pleuropneumoniae. 19 However, the genes linked to these two outer membrane proteins were both downregulated in ciprofloxacin-resistant mutants in our study.
In gram-negative bacteria, the HlyDB-TolC type I secretion system (T1SS) is responsible for secretion of the activated RTX toxin. 42 The apxICA and apxIICA genes are responsible for the production of ApxI and ApxII, respectively, in A. pleuropneumoniae serovars 1 and 7. 43 The secretion of ApxI (serovar 1) and ApxII toxins (serovars 1 and 7) relies on the apxIBD genes. 43 The absence of a consistent and marked decrease in the transcription level of apxIB and apxID in this study suggests that the reduced secretion of Apx toxins in ciprofloxacin-resistant mutants may be due to reduced Apx production. This is supported by the reduced transcriptional levels of apxICA and apxIICA found in this study, although the reduction in apxIC and apxIA (245- and 819-fold, respectively) was greater than that observed in apxIIC and apxIIA (7- and 11-fold, respectively). As the outer membrane protein TolC involved in secretion of Apx toxins has not been identified, the relationship between Apx toxins and antibiotics efflux needs to be further investigated.
A key finding of this study is the phenotypic changes in growth fitness, ability to form biofilm, and ability to secret Apx toxins found in the ciprofloxacin-resistant mutants. However, these phenotypic changes are not necessarily linked to the mutations we have identified in the QRDRs. It is possible that other mutations have occurred that underpin some of the phenotypic changes we have detected. Previous study showed that fluoroquinolone resistance-associated mutation in gyrA affects DNA supercoiling and gene expression, consequently affects bacterial the physiology and fitness of bacterial organisms. 44 Further studies are required to gain a full understanding of the changes we have identified following the acquisition of ciprofloxacin resistance in A. pleuropneumoniae.
In summary, this study reports that adaptation to ciprofloxacin in A. pleuropneumoniae resulted in not only an expected occurrence of broad quinolone resistance but also an unexpected cross-resistance to ampicillin. As well, the ciprofloxacin-resistant mutants showed reduced bacterial fitness as well as changes in efflux ability and biofilm formation. The transcription and secretion levels of Apx toxins were dramatically reduced in ciprofloxacin-resistant mutants compared with their WT strains. These new insights underline the importance of further work seeking to understand the impact on A. pleuropneumoniae, a key pig respiratory pathogen, of exposure to subinhibitory concentrations of antibiotics.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the National Natural Science Foundation of China (31572527), the National Key Research and Development Program of China (2016YFD0500706), and the Special Program on Science and Technology Innovation Capacity Building of BAAFS (KJCX201914).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.