
Abstract
Trimethoprim-/sulfamethoxazole-resistant small colony variants (SCVs) of Staphylococcus aureus, which are selected by use of trimethoprim/sulfamethoxazole, are involved in intractable biofilm-forming infection. This study aimed to determine the biofilm formation ability in trimethoprim-/sulfamethoxazole-resistant SCVs of S. aureus and investigate the bactericidal activity of differential antimicrobial agents to its biofilm-forming S. aureus. Between 32 S. aureus wild type (WT) and 32 SCVs selected from its WT, the amount of formed biofilm was compared. Vancomycin, daptomycin, rifampicin, and minocycline were exposed to biofilm-forming S. aureus to determine viable bacterial counts and its susceptibility. The biofilm-producing quantify of SCVs was approximately twice that formed by its WT. Vancomycin and daptomycin reduce 4 logs the bacterial counts of biofilm-forming WT at 24 hours, but did not affect SCVs. Rifampicin and minocycline considerably decreased both WT and SCVs; however, both bacterial counts recovered to an initial number 48 hours later. These survival strains showed resistance to each drug, and rpoB mutation or tet38 mRNA overexpression was confirmed.
Introduction
S
On the other hand, we reported that 80% of clinical isolates of S. aureus in Japan were hetero-resistant strains, including trimethoprim-/sulfamethoxazole-resistant small colony variants (SCVs). 11 Because these SCVs emerge only on a unique medium, it is not detected in conventional laboratory testing. 11 Generally, the infection caused by a hetero-strain, including SCVs, becomes chronic and shows a tendency to repeat a recurrence. 12
Furthermore, antibiotic administration is usually prolonged in cases of bone and joint infections, especially when there is a prosthetic joint infection caused by S. aureus, 13 because in these infections, there is a strong possibility of biofilm formation. Because various antimicrobial agents cannot easily penetrate the biofilm, treatment becomes very difficult. It was recently pointed out that SCVs are involved in biofilm formation. 14
In this study, we investigated the biofilm-forming ability in trimethoprim-/sulfamethoxazole-resistant SCVs of S. aureus, and antimicrobial tolerability against biofilm-forming SCVs.
Materials and Methods
Bacterial strains
A total of 32 S. aureus clinical isolates, including 17 of MRSA, were used. Furthermore, 32 SCVs selected from each clinical isolate (wild type [WT]) within the mutant selection window of trimethoprim/sulfamethoxazole were also used. These unstable SCVs were prepared by selecting from WT through the challenge of trimethoprim/sulfamethoxazole as previously described. 11 Briefly, WT was incubated for 120 hours since inoculated 1010 CFU onto the Mueller-Hinton agar (MHA), including half of the mutant prevention concentration of trimethoprim/sulfamethoxazole. SCV phenotype was maintained by incubation with trimethoprim/sulfamethoxazole and thymidine during the experiments.
Antimicrobial susceptibility testing
Vancomycin (Sigma-Aldrich, St Louis, MO), daptomycin (Toronto Research Chemicals, Toronto, Canada), rifampicin (WAKO, Osaka, Japan), and minocycline (WAKO) were tested. Antimicrobial susceptibility testing was performed by the standard agar dilution method using MHA plates according to Clinical and Laboratory Standards Institute (CLSI) guidelines. 15 For daptomycin testing, MHA was adjusted at a final concentration of 50 μg/mL of Ca2+ by adding CaCl2. CLSI breakpoints (M100-S28) 16 were used for interpretation of resistance. S. aureus ATCC 29213 was used for quality control.
Biofilm preparation and quantification
A biofilm model was prepared as previously described. 17 Each WT strain and SCVs were suspended in phosphate-buffered saline at a density of McFarland no. 0.5, and then the bacterial suspension was diluted at a ratio of 1:300 in tryptic soy broth containing 1% glucose (TSBG). To form the biofilm, 100 μL of bacterial solution was inoculated to each well of a 96-well polystyrene microtiter plate, which was then incubated for 48 hours at 37°C, aerated by shaking on an orbital shaker, and under anaerobic conditions. The amount of biofilm was demonstrated by adsorption of crystal violet (CV). After the formation of biofilm was confirmed, the medium was removed and each well was rinsed thrice with saline; then, it was air dried. Biofilm was stained with 0.1% CV for 10 minutes and unbound dye was removed by washing the wells thrice with distilled water. The dye that adhered to the biofilm was dissolved with 30% acetic acid (WAKO), and absorption was measured at 595 nm using a Microplate Reader Model 680 (Bio-Rad, Hercules, CA). Furthermore, the amount of biofilm was quantified, formed in presence of TSBG, including the dilution series of trimethoprim/sulfamethoxazole that mixed sulfamethoxazole (WAKO) with trimethoprim (Nacalai tesque, Kyoto, Japan) at 19:1.
RNA extraction and quantitative reverse transcription PCR
To extract mRNA from the biofilm formed by WT or SCVs, each strain was cultured in cell culture dishes (VIOLAMO, Osaka, Japan) as mentioned above. After the bacteria formed biofilm, the medium was removed and the dishes were rinsed thrice with saline. Biofilm-forming S. aureus was scratched using a sterile cotton swab and suspended in milli-Q water. Total RNA was extracted using TRI Reagent™ LS (Molecular Research Center, Inc., Cincinnati, OH). The level of mRNA expression was analyzed as previously described using iTaq Universal SYBR Green One-Step Kit (Bio-Rad) and CFX Connect Real-Time System (Bio-Rad) with specific primers.18–20 The primers sequence was as follows, sigB-F: 5′-TTTCACCTGAGCAAATTAACC A-3′; sigB-R: 5′-TCTTCGTGATGTGATTGTCCTT-3′; sarA-F: 5′-TCTTGTTAATGCACAACAACG TAA-3′; sarA-R: 5′-TGTTTGCTTCAGTGATTCGTTT-3′; agrA-F: 5′-GCACATACACGCTTACAATTGTTG-3′; agrA-R: 5′-ACACTGAATTACTGCCACGTTTTAAT-3′; gyrB-F: 5′-CAAATGATCACAG CATTTGGTACAG-3′; and gyrB-R: 5′-CGGCATCAGTCATAATGACGA T-3′. The relative level of mRNA expression of the target gene was standardized against the expression gyrB and displayed as a multiple of that of ATCC 29213 using the standard curve method.
Measurement of antimicrobial activity against biofilm-forming S. aureus
To determine antimicrobial activity against biofilm-forming S. aureus, vancomycin, daptomycin, rifampicin, and minocycline were used. The concentration of vancomycin, daptomycin, rifampicin, and minocycline was 16, 16, 8, and 2 μg/mL, respectively. These were considered to a trough level at the usual dose administration of each drug.21–24 Biofilm formation was induced on 96-well microtiter plates as mentioned above. One-hundred microliters of TSBG containing an antimicrobial agent was added to the biofilm that had formed in each well, and the plate was incubated for 72 hours. Every 24 hours, the medium and planktonic cells were removed and biofilm-forming strains were rinsed thrice with saline; then, added fresh medium containing the corresponding antimicrobial agent was added. Every 24 hours, the count of surviving bacteria was determined by measuring absorption in a microplate reader model 680 (Bio-Rad),25,26 and calculated from a calibration curve. Furthermore, MICs were determined by the agar dilution method after the SCV phenotype reversed to the normal colony.
Analysis of DNA sequence of rpoB gene
The rpoB gene sequence of the strain whose resistance to rifampicin was confirmed was analyzed. The PCR and DNA sequencing primer used for rpoB were rpoB1: 5′-ACCGTCGTTT ACGTTCTGTA-3′ and rpoB2: 5′-TCAGTGTAGCATGTGTATC-3′ as previously described. 27 The amplification procedure comprised denaturation at 95°C for 3 minutes followed by 30 cycles of denaturation for 30 seconds at 94°C, annealing for 30 seconds at 63°C, and polymerization for 1 minute at 72°C. The amplification product was purified using Fast Gene™ Gel/PCR Extraction Kit (NIPPON Genetics Co., Ltd. Tokyo, Japan) and then sequenced by the Dye Terminator Cycle Sequencing method using Genomelab GeXP and Quick Start Kit (Beckman Coulter, Inc., CA).
Expression level of tet38 mRNA
The expression level of tet38 mRNA in minocycline-resistant strains was analyzed. RNA extraction and quantitative reverse transcription PCR were performed as already mentioned. Primers of tet38-F: 5′-TGACAGGTGTGGCTATTGGT-3′ and tet38-R: 5′-TTGCCTGGGAAATTTAATGC-3′ were used. 28 The level of mRNA expression of the target gene was normalized using the standard curve method against the expression level of gyrB and was shown as a multiple of that of the parental strain.
Statistical analysis
All experiments were performed in duplicate. Student's t-test was used to compare the amount of biofilm, the expression level of mRNA, and bactericidal activity of antibiotics between WT and SCVs. A p-value of <0.05 was considered statistically significant.
Results
Antimicrobial susceptibilities
The susceptibility profile of each antimicrobial against S. aureus clinical isolates is shown in Table 1. All WT strains were susceptible to vancomycin, daptomycin, and rifampicin. Four strains (23.5%) of MRSA strains were resistant to minocycline.
Antimicrobial Activity of Each Antimicrobial Agent Against Clinical Isolates of Staphylococcus aureus
MIC, minimum inhibitory concentration; MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-susceptible Staphylococcus aureus.
Amount of biofilm formed by S. aureus
SCVs having a colony size of 1 mm or less were observed on the agar, including submutant prevention concentration of trimethoprim/sulfamethoxazole. In the absence of trimethoprim/sulfamethoxazole, the amount of biofilm formed by WT strains of MSSA showed an absorbance of 0.4 ± 0.3, and that of MRSA an absorbance of 0.6 ± 0.2 (Fig. 1). On the other hand, the amount of biofilm in MSSA SCVs was 1.0 ± 0.5, which was significantly higher compared with that formed by its WT strains (p < 0.05) (Fig. 1A). Similarly, the amount of biofilm formed by SCVs of MRSA was approximately two times that of WT strains (p < 0.05) (Fig. 1B). Biofilm formation by WT strains, with or without methicillin resistance, was inhibited concentration dependently by trimethoprim/sulfamethoxazole. On the other hand, although similar tendency was shown in SCVs, biofilm amount was 5–7 times high compared with that of WTs.

Amount of biofilm formed by Staphylococcus aureus. The amount of biofilm formed by MSSA
Expression level of biofilm-regulating gene mRNA in biofilm-forming S. aureus
The expression level of sigB mRNA in biofilm-forming SCVs of MSSA and MRSA was 0.6 + 0.6 and 1.2 + 0.9, respectively. These values were 2.9-fold (p < 0.05) and 6.7-fold (p < 0.05) that in WT strains. Furthermore, sarA mRNA in SCVs of MSSA and MRSA was 5.1-fold (p < 0.05) and 2.1-fold (p < 0.05) that in WT strains, respectively. The expression of agrA mRNA in MRSA SCVs was only 0.6-fold (p < 0.05) that in WT strains (Fig. 2).
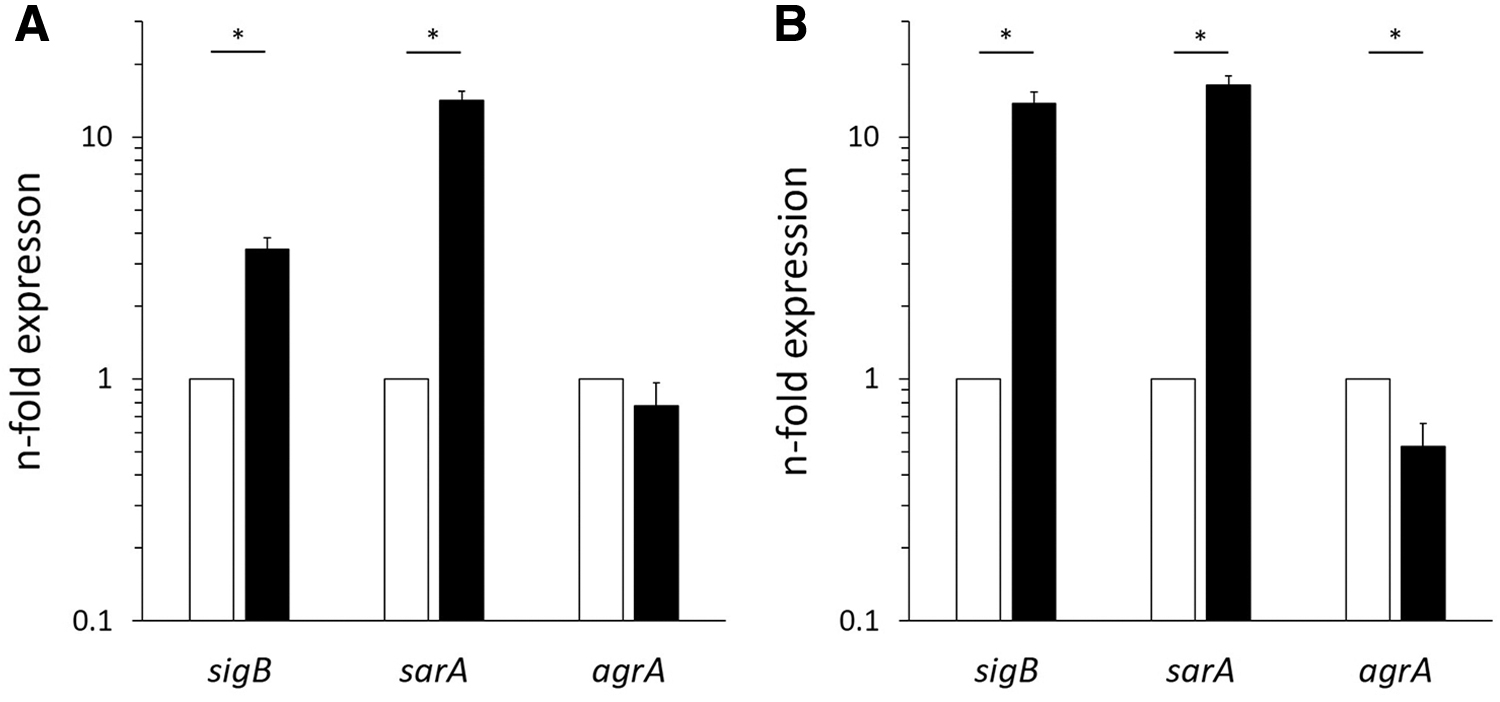
Relative expression of mRNA in biofilm-embedded Staphylococcus aureus. The expression in MSSA
Bactericidal activity of each antimicrobial agent against biofilm-forming S. aureus
After exposure to vancomycin for 24 hours, the bacterial count of WT strains of MSSA and MRSA decreased to 2.5 ± 1.8 and 2.7 ± 2.4 log CFU/mL, respectively. Besides, these bacterial counts persisted for 72 hours (Fig. 3A, E). On the other hand, the bacterial count of SCVs decreased slightly. This tendency was similar in MRSA WT strains exposed to daptomycin (p < 0.05) (Fig. 3F). With all antimicrobial agents, except for WT strains exposed to vancomycin, an increase of WT and SCV bacterial counts was observed after exposure for 72 hours (Fig. 3). Especially, the bacterial count of SCVs of MRSA exposed to vancomycin or daptomycin for 72 hours later was ∼100 times higher compared with the WT (p < 0.05). Besides, the bacterial counts recovered to a value close to the baseline count (Fig. 3E, F).

Antimicrobial activity against biofilm-embedded Staphylococcus aureus. The transition of viable count in MSSA
Acquisition of resistance after exposure to each antimicrobial agent
Number of resistant strains after exposure to each antimicrobial agent is shown in Table 2. All 64 strains exposed to vancomycin for 72 hours maintained their susceptibility to vancomycin. However, 13 out of 64 strains showed the minimum inhibitory concentration (MIC) of vancomycin was 2 μg/mL, and 10 of those strains were SCVs. Moreover, 6 out of the 13 strains (including five strains of SCVs) were not susceptible to daptomycin. On the other hand, after exposure to daptomycin for 72 hours, 17 strains were confirmed to be nonsusceptible, and there were more SCVs than WT. For all these strains, the MIC of vancomycin was 2 μg/mL.
Antimicrobial Susceptibility After Exposure to Each Antibiotic
Number of strains that showed MIC of vancomycin = 2 μg/mL.
Only minocycline experiments were performed, n = 13, except for four of resistant strains.
SCV, small colony variant; WT, wild type.
The number of resistant strains emerged after exposure to rifampicin or minocycline was almost the same among WT and SCVs. All rifampicin-resistant strains had at least one mutation in RpoB (Table 3). The mutation of Q431G, R503L, Q520D, I524M, G540S, and F541L was a new mutation. In addition, the expression of tet38 mRNA in minocycline-resistant strain was ∼10-fold higher compared with WT (p < 0.05) (Fig. 4).
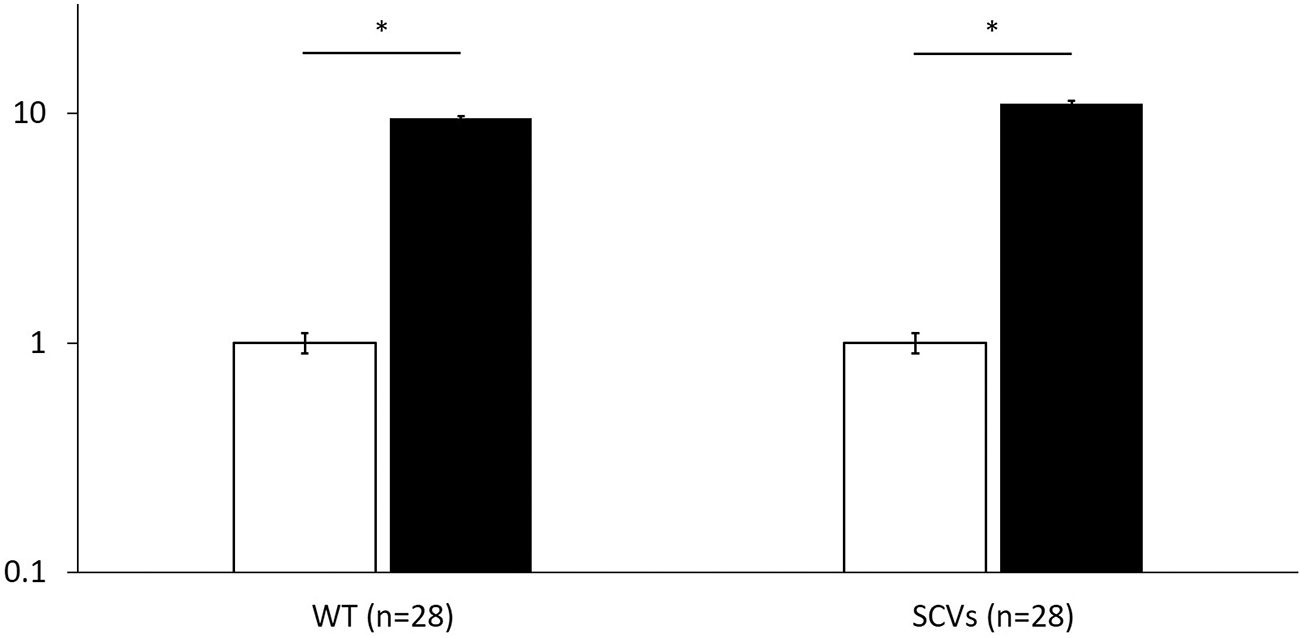
The expression level of tet38 mRNA in minocycline-resistant strain. Data are shown as mean ± standard error of means of the parental strain (open columns) and that of minocycline-resistant strain (filled columns). *p ≤ 0.05 (Student's t-test).
The RpoB Mutation Point in Rifampicin-Resistant Strain
Discussion
Recently, SCVs were reported to relate to chronic infection or recurrent cases of infection caused by S. aureus. 29 Furthermore, most of patients with these infections are administered trimethoprim/sulfamethoxazole for a long term.30–32 We clarified that S. aureus SCVs can be selected by 5 days within the mutant selection window of trimethoprim/sulfamethoxazole, and reported that 80% of S. aureus isolated in Japan correspond to hetero-resistant strains, including trimethoprim-/sulfamethoxazole-resistant SCVs. 11 The use of trimethoprim/sulfamethoxazole for S. aureus infection probably produced these trimethoprim-/sulfamethoxazole-resistant SCVs.
Trimethoprim/sulfamethoxazole is reported to inhibit biofilm formation of Escherichia coli, Pseudomonas aeruginosa, and Acinetobacter baumannii. 33 In this study, biofilm formation by WT and SCVs of S. aureus was inhibited concentration dependently by trimethoprim/sulfamethoxazole (Fig. 1). Generally, because S. aureus grow as WT at the infection site, exposure to trimethoprim/sulfamethoxazole for 48 hours may inhibit biofilm formation depending on its concentration, as shown in this study. However, in case of SCVs selected by exposure to trimethoprim/sulfamethoxazole for 5 days, the amount of biofilm formed by SCVs was twofold that of WT, and five to sevenfolds in case of continued exposure to trimethoprim/sulfamethoxazole. Namely, if trimethoprim/sulfamethoxazole is administered for 5 days or more to treat a device-related infection, SCVs may be selected and biofilm formation be encouraged. Furthermore, the expression of sigB mRNA, which regulates biofilm formation of S. aureus, 34 was two to sixfolds that found in WT. It is known that SCVs regulate various metabolic pathways through the upregulation of sigB expression. Namely, SCVs selected by trimethoprim/sulfamethoxazole show high biofilm formation due to the overexpression of sigB. The expression of sarA mRNA was fivefold that observed in WT. sarA is the main promotor of biofilm formation in MSSA, it leads to an increased production of extracellular polysaccharide due to transcription of the icaADBC operon. 35 This mechanism was considered the reason for the high biofilm formation by MSSA SCVs. sarA-upregulated expression was confirmed in MRSA SCVs, as well. However, transcription of the icaADBC operon by sarA is known to be hardly promoted in MRSA 36 ; therefore, sarA may play no role in biofilm formation by MRSA SCVs. On the other hand, agrA was reported as another gene that regulates biofilm formation in MRSA. 37 agrA, which is related to the expression of the protease group that dissociates binding between extracellular DNA and an adhesion protein, is suppressed by sigB. Actually, the expression of agrA mRNA in MRSA SCVs decreased by more than half compared to that observed in WT. Therefore, the large amount of biofilm produced by MRSA SCVs was probably due to suppression of biofilm disruption as the result of a decreased expression of proteases through decreased expression of agrA.
Then, the antibacterial efficacy of each antimicrobial agent against biofilm-forming WT or SCVs was investigated. Each concentration of vancomycin, daptomycin, rifampicin, and minocycline was equivalent to the trough serum concentration in a stationary state when administered at the usual dosage. In other words, it was corresponding to clinical drug concentration. After exposure of vancomycin (16 μg/mL) for 24 hours to WT strains, bacterial counts of both MSSA and MRSA decreased by 4 logs, but there was hardly a change in the count of SCVs. This was thought to be due to the higher biofilm-forming ability of SCVs than WT, and a weak bactericidal effect against slow-growing SCVs because vancomycin does not penetrate the biofilm too easily. 38 In the case of daptomycin, not only the count of WT but also that of SCVs decreased by 4 logs in MSSA. Daptomycin probably affects slow-growing SCVs, unlike vancomycin. 39 However, changes in the count of MRSA SCVs were similar to those seen with vancomycin. As the reason for the difference between MSSA and MRSA, the involvement of upregulated expression of sigB and sarA mRNA was considered. The upregulated expression of sigB has been reported to decrease daptomycin susceptibility through expression of sarA. 40 The expression of sigB and sarA mRNA was significantly higher in SCVs of MRSA (p < 0.05).
After exposure to vancomycin for 72 hours, the bacterial counts of SCVs increased, while count of WT increased. No vancomycin-resistant strains were detected, but for 13 out of 64 strains, the MIC of vancomycin was 2 μg/mL. Forty-six percent of these 13 strains were also daptomycin nonsusceptible. In addition, strains for which the MIC of vancomycin was 2 μg/mL were found more frequently in SCVs than WT. Therefore, the selection of SCVs may reduce the effect of treatment with vancomycin. Otherwise, an increase of the bacterial counts was observed in WT and SCVs after exposure to daptomycin. That was probably due to decreased susceptibility to daptomycin. Actually, 17 out of 64 strains were nonsusceptible after being exposed to daptomycin and the MIC of vancomycin for them was also 2 μg/mL.
Rifampicin and minocycline decreased bacterial counts of WT and SCVs by ∼4 logs. These two drugs are known to show antimicrobial activity because they permeate the biofilm. 38 So it is thought that these antimicrobials would show activity against SCVs having a high biofilm-forming ability. However, these results showed an increase of bacterial counts of WT and SCVs cultured in the presence of either rifampicin or minocycline for 24 hours. It is known that S. aureus easily acquires resistance when rifampicin and minocycline are used as single agents.41,42 Actually, after exposure to rifampicin or minocycline, most survival strains will have acquired resistance to these drugs. The reason for the increased bacterial counts was acquired resistance. The rifampicin-resistant strain detected in this study carried at least one RpoB mutation point. Furthermore, one mechanism of tetracycline resistance is high expression of Tet38, which is a drug efflux pump encoded in the chromosome. 43 The expression of tet38 mRNA in strains that acquired resistance to minocycline was 10-fold that found in WT. Therefore, the use of rifampicin or minocycline alone for S. aureus infection may increase the isolation frequency of resistant strains with or without SCVs. In S. aureus infections that encountered the orthopedic field, SCVs are known not only to form biofilm but also to survive a long time by invading osteoblasts. 44 Rifampicin and minocycline have good transferability to osteoblasts, but β-lactams or glycopeptides are not good. Therefore, SCVs are exposed to rifampicin or minocycline alone when these combined with β-lactams or glycopeptides in osteoblast cell infection. S. aureus acquires resistance to these drugs with high frequency and relatively early after treatment.
Conclusion
SCVs emerge after exposure for 5 days or more to trimethoprim/sulfamethoxazole in areas hard to be sterilized by various antimicrobial agents due to a high biofilm organization potency. Furthermore, these SCVs easily acquire drug resistance. Therefore, administration for 5 days or more of trimethoprim/sulfamethoxazole for S. aureus infections promotes biofilm formation, which would weaken the effect of treatment with the antimicrobial agent.
Footnotes
Acknowledgment
We thank Dr. Akira Watanabe of Tohoku Infectious Disease Society for offering S. aureus clinical isolates.
Disclosure Statement
All authors have no conflict of interests to declare. All authors meet the ICMJE authorship criteria.
Funding Information
No funding was received for this article.