
Abstract
The populations of extended-spectrum β-lactamase-producing Escherichia coli (ESBL-EC) have increasingly disseminated in humans, animals, and the environment. This study aimed to determine the prevalence, antimicrobial susceptibilities, and molecular characteristics of ESBL-EC isolates obtained from vegetable farm soil. In total, 200 soil samples were collected from vegetable farms in Incheon, South Korea, between 2018 and 2019 and cultured on MacConkey screening plates supplemented with 2 μg/mL cefotaxime. Cefotaxime-resistant ESBL-EC isolates were recovered from 4.0% (8/200) of the soil samples. All eight isolates were nonsusceptible to ampicillin, piperacillin, cefazolin, cefotaxime, and cefepime and harbored blaCTX-M-type ESBL genes, including blaCTX-M-15 (50.0%), blaCTX-M-55 (25.0%), and blaCTX-M-14 (25.0%). Phylogenetic analysis showed that the B1 lineage was predominant (75.0%), followed by A (12.5%) and B2 (12.5%) lineages. Multilocus sequence typing revealed eight different E. coli sequence types (STs), including ST10, ST73, ST155, ST847, ST2521, ST3285, ST5173, and ST9479. Notably, ST10 and ST73 belong to the global extraintestinal pathogenic E. coli lineages. Our findings demonstrated that the farm soil environment may serve as a reservoir of human-associated multidrug-resistant ESBL-producing pathogens.
Introduction
The emergence and widespread of antimicrobial-resistant bacteria have become a barrier to infection control. Enterobacteriaceae can acquire resistance to oxyimino-cephalosporins, such as cefotaxime, by expressing extended-spectrum β-lactamases (ESBLs). In particular, ESBL-producing Escherichia coli strains (ESBL-ECs) are increasingly causing serious problems in both human and veterinary medicine. 1 ESBLs can be divided into three main types: TEM, SHV, and CTX-M. 2 Of these, CTX-M-type ESBLs are the most prevalent ESBL type worldwide,2,3 exceeding 230 variant counts (ftp://ftp.ncbi.nlm.nih.gov/pathogen/Antimicrobial_resistance; accessed October 3, 2020); these are further classified into five major groups according to the sequence similarity of amino acids: CTX-M-1, -2, -8, -9, and -25. 2
Most blaCTX-M genes are present in E. coli strains; of these the pandemic sequence type (ST) 131 lineage commonly harboring blaCTX-M-15 has played an important role in the global dissemination of ESBLs. 4 CTX-M-15 is the most common ESBL enzyme in most regions. 3 In South Korea, CTX-M-14 has been found to be most dominant since it was first discovered in clinical isolates of E. coli, Shigella sonnei, and Klebsiella pneumoniae in 2001.5–8 The fecal carriage rate of CTX-M-type ESBL-ECs in healthy Korean adults was 8.3% in 2014. 9 The South Korean national antimicrobial resistance surveillance system (Kor-GLASS) was established in 2016 and reported that the cefotaxime-resistance rate of clinical E. coli isolates reached 32.4% in 2017.10,11
Livestock is considered a major reservoir of ESBL producers that can cause infection and spread of antimicrobial resistance (AMR) in humans, possibly because of the selective pressure exerted by the use of antimicrobial agents in animal management. 12 In South Korea, the prevalence rates of ESBL-ECs were 94.1% in chickens, 69.5% in pigs, and 7.0% in cattle, with CTX-M-14, -55, and -65 being dominant in each livestock species in 2018. 13 CTX-M-65 and -55 have also been commonly detected in E. coli isolates from ducks and raw retail chickens.14,15 In addition to humans and livestock, ESBL-ECs have been broadly isolated from other potential reservoirs, including companion animals, wildlife, food products, and the natural and farm environments.3,16–19 Numerous studies performed in South Korea have reported the prevalence of ESBL-ECs in various sectors, including 44.7% in companion animals, 20 0.83% in raw vegetables, 21 3.3% in ready-to-eat sprouts, 22 and 2.5% in river water. 23 Based on the One Health approach that combines the intervention and prevention strategies across the three domains of humans, animals, and the environment, many AMR surveillances have been implemented locally, nationally, and globally24,25; however, there is still a lack of information about environmental resistance.
In this study, we investigated the prevalence, antimicrobial susceptibilities, and β-lactamase gene profile of ESBL-ECs isolated from vegetable farm soil in South Korea to determine their epidemiological relatedness to human-associated strains according to the national monitoring program.
Materials and Methods
Screening for ESBL-ECs from vegetable farm soil
A total of 200 farming soil samples (100 each between May and September of 2018 and 2019) were collected from the top soil layer (0 − 20 cm) of 200 different vegetable farms in Incheon, South Korea. Twenty grams of each soil sample was incubated in 180 mL of E. coli (EC) selective enrichment broth (Difco Laboratories, Detroit, MI) at 37°C for 24 hr. Subsequently, 1 mL of enriched medium was added to 9 mL of tryptone soya broth (Oxoid, Basingstoke, United Kingdom) supplemented with 0.4 g/mL vancomycin (Wako Pure Chemical Industries, Osaka, Japan), followed by incubation at 37°C for 4 hr. A loopful of the enrichment was streaked on a MacConkey agar plate containing 2 μg/mL cefotaxime and incubated at 37°C for 24 hr. One pink or reddish colony suspected of comprising E. coli was randomly picked (one for each plate where the colony growth was observed) using a sterile platinum loop, and further cultured on CHROMagar ESBL (CHROMagar, Paris, France) plate at 37°C for 24 hr. A single colony with dark pink to reddish hue from each sample was grown on tryptone soya agar (Oxoid) at 37°C for 4 hr and the nonduplicate isolate was further characterized. E. coli species of the pure isolates was confirmed by matrix-assisted laser desorption ionization–time-of-flight mass spectrometry (Bruker Daltonik GmbH, Bremen, Germany) based on a score of ≥2.0.
Antimicrobial susceptibility testing
Antimicrobial susceptibilities were tested for 22 agents from 15 antimicrobial classes. The susceptibilities for 21 agents except for colistin were tested by the disk diffusion method on Mueller–Hinton agar (Difco Laboratories) using 6-mm antibiotic disks purchased from Oxoid, as follows: amikacin (30 μg), amoxicillin−clavulanic acid (20/10 μg), ampicillin (10 μg), ampicillin−sulbactam (10/10 μg), aztreonam (30 μg), cefazolin (30 μg), cefepime (30 μg), cefoxitin (30 μg), ceftazidime (30 μg), chloramphenicol (30 μg), ciprofloxacin (5 μg), cefotaxime (30 μg), ertapenem (10 μg), gentamicin (10 μg), imipenem (10 μg), meropenem (10 μg), nalidixic acid (30 μg), piperacillin (100 μg), tetracycline (30 μg), tigecycline (15 μg), and trimethoprim−sulfamethoxazole (1.25/23.75 μg). The susceptibility to colistin was determined through the minimum inhibitory concentration by the broth microdilution method using the Sensititre system (Thermo Fisher Scientific, MA). The interpretation of resistance, intermediate resistance, and susceptibility complied with the Clinical and Laboratory Standards Institute M100-S27, 26 except for tigecycline, which complied with the European Committee on Antimicrobial Susceptibility Testing breakpoint version 7.1. 27 Multidrug resistance (MDR) was determined by nonsusceptibility (resistance or intermediate resistance) to at least one antimicrobial agent of three or more classes. 28 E. coli ATCC 25922 strain was used as a control for the testing.
Characterization of ESBL genes
DNAs were purified from the isolates using the G-spin™ Total DNA Extraction Kit (iNtRON Biotechnology, Seoul, Korea) in accordance with the manufacturer's instructions. Polymerase chain reaction (PCR) and sequencing were conducted using the specific primers for blaCTX-M-1, blaCTX-M-2, blaCTX-M-9, blaCTX-M-25, blaTEM, and blaSHV groups, respectively. 13 The amplicon sequences were analyzed using the NCBI BLAST program (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Phylogenetic analyses
The DNA extracts were subjected to multiplex PCR for the detection of chuA, yjaA, and TspE4.C2, and the isolates were subsequently assigned to phylogenetic groups A (chuA−, TspE4C2−), B1 (chuA−, yjaA−, TspE4.C2+), B2 (chuA+, yjaA+), and D (chuA+, yjaA−), as reported previously. 29 E. coli STs were determined by multilocus sequence typing (MLST) using the internal fragment sequences of seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) in accordance with the EnteroBase protocol and database. MEGA version X software was used to construct the maximum-likelihood phylogenetic tree with a bootstrap analysis of 1000 replicates based on the seven housekeeping gene sequences.
Results
Prevalence and antimicrobial susceptibility
In total, 8 cefotaxime-resistant ESBL-ECs (8/200, 4.0%) were recovered from 200 soil samples. Antimicrobial susceptibility testing showed that all these isolates were nonsusceptible (were resistant or intermediate resistant) to ampicillin, piperacillin, cefazoline, cefotaxime, and cefepime; however, they were susceptible to amikacin, ertapenem, imipenem, meropenem, cefoxitin, trimethoprim–sulfamethoxazole, tigecycline, and colistin (Fig. 1). The nonsusceptibility rates to aztreonam, tetracycline, nalidixic acid, ampicillin–sulbactam, ceftazidime, gentamicin, ciprofloxacin, amoxicillin–clavulanic acid, and chloramphenicol were 87.5%, 62.5%, 50.0%, 37.5%, 37.5%, 25.0%, 25.0%, 25.0%, and 25.0%, respectively. All the isolates showed MDR phenotypes nonsusceptible to three to nine antimicrobial classes.
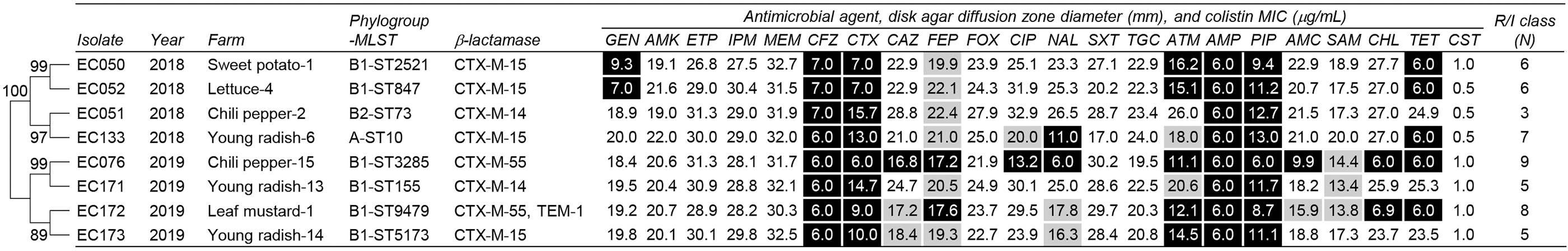
Characteristics of eight cefotaxime-resistant Escherichia coli isolated from vegetable farm soil. Antimicrobial susceptibility was determined using the disk agar diffusion assay, with exception that antimicrobial susceptibility to colistin was determined on the basis of the MIC using the broth microdilution method. Resistance and intermediate resistance are indicated with black and gray, respectively. The numbers in the branches of the phylogenetic tree indicated bootstrap support percentages of 1,000 replicates. Antimicrobial classes were categorized as follows: aminoglycosides (GEN, gentamicin; AMK, amikacin), carbapenems (ETP, ertapenem; IPM, imipenem; MEM, meropenem), nonextended-spectrum cephalosporins (CFZ, cefazolin), extended-spectrum cephalosporins (CTX, cefotaxime; CAZ, ceftazidime; FEP, cefepime), cephamycins (FOX, cefoxitin), fluoroquinolones (CIP, ciprofloxacin; NAL, nalidixic acid), folate pathway inhibitors (SXT, trimethoprim–sulfamethoxazole), glycylcyclines (TGC, tigecycline), monobactams (ATM, aztreonam), penicillins (AMP, ampicillin; PIP, piperacillin), penicillins plus β-lactamase inhibitors (AMC, amoxicillin–clavulanic acid; SAM, ampicillin–sulbactam), phenicols (CHL, chloramphenicol), tetracyclines (TET, tetracycline), and polymyxins (CST, colistin). MIC, minimum inhibitory concentration; MLST, multilocus sequence typing; N, number; R/I, resistant or intermediate-resistant; ST, sequence type.
ESBL gene profiling
Next, we assessed the profiles of ESBL genes belonging to blaCTX-M-1, blaCTX-M-2, blaCTX-M-9, blaCTX-M-25, blaTEM, and blaSHV groups. All eight isolates harbored one of the blaCTX-M group genes. The most predominant group was blaCTX-M-1 (6/8, 75.0%); blaCTX-M-15 and blaCTX-M-55 were detected in four and two isolates, respectively (Fig. 1). One of the blaCTX-M-55-positive isolates cocarried non-ESBL blaTEM-1. Moreover, blaCTX-M-14 belonging to the blaCTX-M-9 group was identified in two samples (2/8, 25.0%). The bla genes belonging to blaCTX-M-2, blaCTX-M-25, and blaSHV groups were not detected in this study.
Phylogenetic characterization
Phylogenetic analysis using a PCR-based assay showed that the eight isolates belonged to B1 (6/8, 75.0%), A (1/8, 12.5%), and B2 (1/8, 12.5%) (Fig. 1). MLST revealed eight different E. coli STs: ST10, ST73, ST155, ST847, ST2521, ST3285, ST5173, and ST9479. The isolates were divided into two major distinct branches of the phylogenetic tree, which were correlated with the sampling years.
Discussion
The recovery rate of ESBL-ECs in vegetable farm soil in Incheon, South Korea, was 4.0% in both 2018 and 2019; this value was lower than that reported previously in China (17.4%) and Tunisia (7.3%).30,31 However, since the samples in this study were collected only at a single point on each farm, further investigation using samples at multiple points is needed to accurately estimate the true prevalence of ESBL-ECs in farm soil. All eight isolates in this study had MDR phenotypes; the average number of nonsusceptible classes increased from 5.5 in 2018 to 6.8 in 2019. ESBLs can usually hydrolyze extended-spectrum cephalosporins and monobactams but not carbapenems and cephamycins. 2 In addition, most of the clinical ESBL-ECs are susceptible to amikacin and tigecycline. 32 Consistent with these characteristics, all the isolates in this study were susceptible to ertapenem, imipenem, meropenem, cefoxitin, amikacin, and tigecycline and showed a high resistance rate of 87.5% to aztreonam.
CTX-M-15 and CTX-M-14 are the most common ESBL variants worldwide. 3 In the past decade, the incidence of ESBL-EC–producing CTX-M-55 has been increasingly reported in humans and animals in Asian countries, including China33,34 and Vietnam. 35 In South Korea, CTX-M-55 has been predominantly detected in pigs (53.7%), 13 retail raw chickens (25.4%), 14 and companion animals (17.2%) 20 ; it is also showing an increasing occurrence in humans. 5 In this study, we isolated two CTX-M-55–producing ESBL-ECs from farm soil collected in 2019. Given that the blaCTX-M-55 gene encodes the A77V variant of CTX-M-15 by a single nucleotide substitution, 36 it is likely that CTX-M-55 is replacing CTX-M-15 in multiple sectors in Asia.
Among E. coli phylogenetic groups, commensal strains are mainly included in A and B1 lineages, whereas more virulent strains are included in B2 and D lineages. 37 In this study, most isolates belonged to commensal B1 (6/8, 75.0%) and A (1/8, 12.5%) lineages, whereas one isolate belonged to the virulent B2 (1/8, 12.5%). Eight different E. coli STs were identified by MLST analysis in this study. Notably, two of them, namely ST10 and ST73, were among the major hyperepidemic extraintestinal pathogenic E. coli (ExPEC) lineages, 38 which are associated with urinary tract infections and bacteremia. 39 Along with the pandemic E. coli ST131, the ST10 lineage is responsible for the rapid dissemination of ESBL genes worldwide. 40 Moreover, ESBL production by this lineage is ubiquitously encountered in various ecological areas, including humans, livestock, vegetables, and the environment.41–45 In South Korea, ESBL-EC ST10 has been detected in chickens,13,46 pigs, 13 raw vegetables,21,47 healthy adults, 9 and patients. 48 We recently found that E. coli ST10 harboring blaCTX-M-55 isolated from lettuce in South Korea cocarried the mcr-1 gene conferring colistin resistance. 47 E. coli strains harboring mcr-1 and mcr-3 have emerged in agricultural soil. 49 However, colistin resistance was not observed in this study. E. coli ST73 is common in humans and companion animals; however, as it is rarely associated with ESBLs,38,50,51 few studies have reported ESBL production by ST73. In clinical ST73 isolates, CTX-M-15, CTX-M-1, CTX-M-9, and OXA-1 have been reported in the United Kingdom,52,53 CTX-M-15 has been reported in Egypt, 54 and CTX-M-14 has been reported in Japan. 55 In South Korea, ESBL production by ST73 has been observed in children with urinary tract infections. 56 Among nonhuman ST73 isolates, CTX-M-15 and SHV-12 have been found in sewage in the United Kingdom 53 and companion animals in the United States, 57 respectively. To the best of our knowledge, this is the first study to identify CTX-M-14–producing ESBL-EC ST73 in nonhuman sectors. The ST73 isolate was found to have a narrower spectrum of resistance (nonsusceptible to three classes) than the other STs (nonsusceptible to five to nine classes). Clinical studies have also revealed ST73 to have a low degree of resistance among ExPEC lineages.38,58
The same ESBL genes, plasmids, and strains have often been shared among ESBL-ECs in humans, animals, food produce, and the environment,41,43,59–61 suggesting their ecological circulation. Irrigation water and manure are the major sources of soil contamination with resistant bacteria.62,63 Irrigation water and surface water for agriculture have a high prevalence of ESBL-ECs.63,64 Similarly, the manure of animal litter has been widely used as organic fertilizer in agricultural farms 16 ; the abundance of blaCTX-M genes has been found to be higher in manured soil than in inorganic fertilized soil. 65 Thus, ESBL-ECs can spread to soil through agricultural environments, such as irrigation water and animal manure, and be transmitted to fresh produce and potentially to humans through the food chain. In conclusion, we demonstrated that soil may serve as a reservoir of ESBL-producing MDR pathogens that can pose a high risk to human health. Our findings highlight the importance of strict monitoring and control strategies in the One Health perspective for combating AMR.
Footnotes
Authors' Contributions
Study conception and design were carried out by J.S.; sample collection, data acquisition, and analysis were taken care of by J.H.S., J.K., S.-S.O., and J.S.; article writing was done by J.S. All authors reviewed the article.
Ethical Approval
Not required.
Data Statement
Available upon request.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Korea Centers for Disease Control and Prevention [2017ER540302], the Basic Science Research Program through the National Research Foundation (NRF) of Korea funded by the Ministry of Education [2018R1A6A1A03025523], and the Inha University Research Grant [2020].