
Abstract
The opportunistic pathogen, Pseudomonas aeruginosa, uses different mechanisms as well as biofilm production to acquire antibiotic resistance. The polysaccharide synthesis locus (psl) genes play an important role in P. aeruginosa biofilm formation. Therefore, targeting the expression of psl genes can be a suitable strategy to prevent the formation of biofilms by antibiotic-resistant strains. Today, advances in nanotechnology provide a novel potential strategy to combat antibiotic-resistant bacteria. In this study, the silver nanoparticles (Ag NPs) synthesized using a chemical co-precipitation method and, after conjugation with thiosemicarbazide, their effect on the biofilm-forming ability are studied in P. aeruginosa isolates. Chemical properties of synthesized nanoparticles were determined by scanning and transmission electron microscopy, Fourier transform infrared spectroscopy, diffuse reflectance spectroscopy, ultraviolet–visible spectroscopy, X-ray diffraction, and energy dispersive X-ray spectroscopy. The results confirmed the spherical/cubic morphology, solution stability, and good dispersion of Ag@Glu–TSC NPs with an average size of 40–60 nm. In addition, minimum inhibitory concentration values of functionalized Ag NPs were at least twofold lower than the Ag NPs (alone). The quantitative PCR data analysis showed a decrease in the expression of the pslA gene in the presence of Ag@Glu–TSC NPs, up to 60%, which was associated with a reduction of biofilm formation compared to control. In conclusion, the Ag@Glu–TSC NPs can be considered a new inhibitor of biofilm production in antibiotic-resistant bacteria.
Introduction
Every day, humans are exposed to a variety of bacterial infections, some of which, despite the development of public health immunization programs, pose a serious threat to people's health. Meanwhile, the emergence of antibiotic-resistant strains due to overuse and misuse of antibiotics has led to the formation of a global crisis about pathogenic bacteria. Currently, at least 700,000 people die each year due to drug-resistant diseases and according to the World Health Organization (WHO), drug-resistant diseases could cause 10 million deaths each year by 2050. 1
At present, with the entrance of nanotechnology in biomedical applications, the antimicrobial properties of nanoparticles have been more considered an antibiotic alternative to combat multidrug-resistant bacteria. Nanoparticles have a higher surface-to-volume ratio, which leads to a more efficient reaction. The exact mechanism of nanoparticle action against microorganisms has not been well understood; however, it seems that silver nanoparticles (Ag NPs) with a size range of 10 to 100 nm can be efficiently taken by living organisms such as bacteria and affect their normal cellular function that is important for their survival. 2
There are several reports for the antibacterial potential of metal nanoparticles such as copper, titanium, magnesium, zinc, gold, and silver. 3 Among these, Ag NPs have attracted the special attention of researchers because of their antimicrobial and anticancer potentials.4,5 Antibacterial properties of Ag NPs alone and in conjugation with chemical antibacterial agents have been studied in many researches. Loo et al. indicated a strong antimicrobial activity for Ag NPs against a diverse range of Gram-negative foodborne pathogens, including Escherichia coli, Klebsiella pneumoniae, Salmonella Typhimurium, and Salmonella Enteritidis. 6
The antibacterial potential of Ag NPs against common human pathogens such as Gram-negative bacteria (E. coli, Pseudomonas aeruginosa, and Salmonella enterica) and Gram-positive bacteria (Bacillus subtilis and Staphylococcus aureus) was analyzed and confirmed by Yousaf et al. 7 Mohsen et al. exhibited higher antibacterial activity for Ag NPs that functionalized with ciprofloxacin against two common Gram-positive (S. aureus) and Gram-negative (E. coli) human pathogens. 8
P. aeruginosa is an opportunistic Gram-negative pathogen that is rarely identified as normal microbial flora in healthy persons. This pathogen can cause a wide range of diseases, including urinary tract infections, respiratory infections, toxic sepsis, and bacteremia, which can even lead to death in people with weakened immune systems. In recent years, the overuse of antibiotic treatment for P. aeruginosa infections and the emergence of multidrug-resistant (MDR) strains have caused serious problems in the treatment of nosocomial infections worldwide. For this reason, P. aeruginosa is on the WHO priority 1 (critical) list of antibiotic-resistant bacteria for research and development of new antibiotics. 9
P. aeruginosa can use a variety of the well-known and characterized mechanisms, such as decrease intracellular drug accumulation by the efflux pumps, changes in target sites, use of alternative metabolic pathways, and drug inactivation or modification to acquired antibiotic resistance. 10 In addition to antibiotic resistance mechanisms, some bacteria, including P. aeruginosa, can form biofilms that reduced their susceptibility to host immune response and conventional antibiotics. 11
Biofilms comprise a community of bacterial cells living within an extracellular polymeric substance (EPS) matrix that attaches to the surface of a biological or abiotic structure. The matrix acts as a protective shield and is mainly composed of water, proteins, polysaccharides, lipids, and extracellular DNA. 12 Also, the EPS matrix helps the bacterial populations to maintain close contact and act as a community through cell-cell communication pathways using quorum-sensing products.13,14
Currently, the most important concern about biofilm-related infections is this sensitivity reduction to antibiotic treatment, which resulted in human infections, particularly chronic infections because of failure to fully eradicate the pathogens. 15 The inability of antimicrobial agents, such as antibiotics, to eradicate biofilm bacteria is due to the physical and chemical properties of the extracellular matrix, which greatly reduce the ability of agents to penetrate microbial communities. Therefore, it is necessary to first know the biofilm structure and its composition. It seems, combined with antibiotic treatment, preventing biofilm formation could be suggested as a good strategy for fighting bacterial infections.
It has been shown that both genetic characterizations of the isolates and environmental conditions can affect biofilm development by P. aeruginosa.16,17 The stability of biofilm structure in P. aeruginosa, is determined by at least three polysaccharides (alginate, Pel, and Psl).18,19 In the initial stages of biofilm formation, Pel and Psl polysaccharides contributed as a primary structural scaffold for matrix development. 20 However, mutational analysis demonstrates that only Psl is a key component for surface attachment of most P. aeruginosa isolates.
There are at least 12 genes in the polysaccharide synthesis locus (psl), 11 of which (pslACDEFGHIJKL) are essential for Psl synthesis and export. 21 Due to their significant role in biofilm development and maintenance, it is possible to increase the susceptibility of P. aeruginosa to antimicrobial agents by repression of psl gene expression.
In recent years, the effect of nanoparticles on gene expression has been confirmed in some studies. Ag NPs, 22 zinc oxide nanoparticles, 23 titanium oxide nanoparticles, 24 and gold nanoparticles 25 were observed to upregulate and downregulate genes in Arabidopsis. Huang et al. showed Ag NPs exposure, could alter gene expression in mouse neural cells. 26 In this study, according to the identification of antibiotic-resistant P. aeruginosa as a health threat and called for the development of new antimicrobial therapies for this pathogen, we investigated the impact of magnetic Ag NPs conjugated with thiosemicarbazide on the expression of psl gene and the biofilm formation capacity of clinical strains of P. aeruginosa. The choice of thiosemicarbazide for conjugation with Ag NPs is due to its antimicrobial activity against Gram-negative and Gram-positive bacteria.27,28
Materials and Methods
All the chemicals, culture media, and reagents used in this study were of analytical grade and purchased from E. Merck KG (Darmstadt, Germany).
Synthesis of the Ag@Glu–TSC nanoparticles
First, Ag NPs synthesized, and functionalized using glutamic acid (Glu), glutamic acid acts as a linker for the conjugation of Ag NPs with thiosemicarbazides (TSC). Then Tsc was conjugated to Ag@Glu NPs through covalent bond formation between carboxylic groups of Glu and amino-terminal groups of thiosemicarbazide. Briefly, 100 mg glutamic acid was added to the AgNO3 solution (0.85 mg/mL) and was heated for 30 min to get a clear solution. The solution was added drop by drop to a solution that contains 0.4 mg/mL NaBH4 and was mixed vigorously for 3 hr at room temperature.
Ag@Glu NPs were separated by centrifugation at 2,500 g for 15 min, washed using distilled water, and dried in a vacuum oven at 70°C for 3 hr. 23 For the preparation of Ag@Glu–TSC NPs, 0.50 g of Ag@Glu NPs and 0.20 g thiosemicarbazide were added to 100 mL 96% ethanol. After sonication for 30 min (Misonix Sonicator 3000, 30 W), the mixture was stirred overnight at 40°C. The prepared nanoparticles were collected by centrifugation at 2,500 g for 15 min and then washed three times with distilled water and 96% ethanol, respectively. Finally, Ag@Glu–TSC NPs dried at 60°C for 8 hr and stored at room temperature before testing their antibacterial properties. 29
Characterization of Ag@Glu–TSC NPs
Electron microscopy and diffraction methods combined to demonstrate the synthesis of Ag@Glu–TSC NPs. The scanning and transmission electron microscopy (SEM and TEM) images were taken using a Zeiss EM10C 80 KV and KYKY-EM3200 electron microscopes, respectively. The colloidal stability of the nanoparticles was characterized by the zeta potential analysis. The PerkinElmer®LAMBDA™ was used to monitor the differential reflectance spectroscopy (DRS) in a wavelength range of 200–1,000 nm.
The infrared spectra of samples in the form of potassium bromide pellets were obtained by a Shimadzu 8900 FT-IR spectrophotometer. For elemental analysis of functionalized nanoparticles, an energy dispersive X-ray spectrometer, MIRA3 FEG-SEM series, was used. Powder X-ray diffraction (XRD) patterns of Ag NPs and Ag@Glu–TSC NPs were recorded using a Philips® X-Pert MPD diffractometer with Cu Kα radiation (λ = 0.1542 nm).
Bacterial strains and resistance profiling
P. aeruginosa isolates were collected from skin ulcers and urine of infected patients referred to some of the Rasht laboratories, in 2016 and 2017. The patients have not participated directly in sampling. The isolates were identified using Gram staining, cetrimide agar combined with biochemical tests such as fermentation of mannitol, oxidase, catalase, Methyl Red/Voges-Proskauer test, Simmon's Citrate Agar, Urease, and hydrogen sulfide production test. The antibiotic resistance profiles of the P. aeruginosa isolates were determined in triplicate against ciprofloxacin (5 μg), gentamicin (10 μg), amikacin (30 μg), tobramycin (10 μg), piperacillin (100 μg), imipenem (10 μg), and ceftazidime (10 μg) by Kirby-Bauer's disk diffusion method.
After an 18-hr incubation at 37°C, strains with resistance to at least three antibiotics of different classes were identified as MDR isolates.
Antibacterial assay
The MIC (minimum inhibitory concentration) values of thiosemicarbazide, Ag NPs, and Ag@Glu–TSC NPs were determined by the broth microdilution testing method in 96-well plates. First, the pathogenic isolates plated on Mueller Hinton agar and incubated for 24 hr at 37°C. The overnight-grown bacteria were swabbed into the nutrient broth medium to prepare a suspension with 0.5 McFarland turbidity (1.5 × 108 CFU/mL). Subsequently, 100 μL of bacterial suspension was added to the wells containing different concentrations of Ag NPs, TSC, and Ag@Glu–TSC NPs (32, 64, 128, 256, 512, and 1,024 μg/well) and incubated at 37°C for 24 hr. The diameters of inhibition growth zones are measured to determine the antibacterial efficacy of different agents.
Ciprofloxacin (5 μg/disc) and dimethyl sulfoxide were used as positive and negative controls, respectively. For each antibacterial agent, the following equation was used for calculating the mean surface area of the inhibition zone and assessment of the increase in fold area (a and b are the zones of inhibitions recorded for different antimicrobial agents).
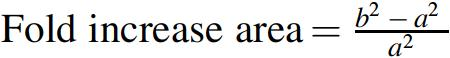 Also, the sub-MICs of antimicrobial agents were determined as the noninhibitory concentration that is still effective in altering bacterial physicochemical properties and morphology to investigate its effects on biofilm formation ability. All experiments were done in triplicates with an interpretation in agreement with the Clinical Laboratories Standard Institution (CLSI) guideline.
30
Also, the sub-MICs of antimicrobial agents were determined as the noninhibitory concentration that is still effective in altering bacterial physicochemical properties and morphology to investigate its effects on biofilm formation ability. All experiments were done in triplicates with an interpretation in agreement with the Clinical Laboratories Standard Institution (CLSI) guideline.
30
Biofilm formation assay by microtiter plate method
Biofilm formation occurs when bacteria switch from a planktonic (free swimming) state to a surface-attached state. In this study, we used microtiter plate assay (also called 96-well plate) by the method proposed by Tendolkar et al., 31 with some modifications to evaluate biofilm formation ability and inhibition in P. aeruginosa. In brief, the clinical isolates were grown for 24 hr at 37°C in trypticase soy broth (TSB), and then the cultures were adjusted equivalent to 0.5 McFarland's standard for biofilm assays. A volume of 200 μL of bacterial suspensions was used to inoculate each well of sterile U-bottom shape 96-well polystyrene microtiter plates (Greiner-Bio One, German).
For each isolate, four wells were used separately and repeated in triplicate. Four wells were assigned to negative control (containing 200 μL of pure TSB without bacteria). The plate incubated overnight at 37°C. After incubation, the bacterial suspension of each well was gently removed and then the wells washed with 0.2 mL of phosphate-buffered saline (pH7.2) three times to remove planktonic cells. Adherent bacteria were stained with 200 μL of crystal violet solution in water (0.1%, w/v). Microtiter plates were washed with deionized water to remove excess stain and kept upside down overnight for drying. After drying, each well was incubated in 200 μL of acetic acid solution (30% in water) at room temperature for 10 min.
The optical densities (OD) of solution were determined as the amounts of biofilm formation by using an ELISA reader (Awareness Technology, Inc.) at 595 nm. Acetic acid solution (30% in water) was used as the blank and the experiment was performed in triplicates. Finally, based on the criteria described by Stepanovic et al., 32 the clinical isolates were classified as no biofilm producer, moderate biofilm producer, and strong biofilm producer.
Biofilm inhibition assay
For the preparation of bacterial suspensions, the same protocol was used. Each well of a microtiter plate was pipetted with 100 μL of bacterial suspension and incubated overnight at 37°C with sub-MICs of Ag NPs, TSC, and Ag@Glu–TSC NPs. For each antibacterial agent, the four wells were used separately and repeated in triplicate. The formation of biofilms was measured using the microtiter plate method as defined in the biofilm formation assay section. The following equation was used to calculate the percentage of biofilm inhibition:

where OD control refers to the absorbance of the bacterial growth at 595 nm without antibacterial agent treatment.
RNA extraction and complementary DNA production
After treatment of pathogenic isolates with sub-MIC of thiosemicarbazide, Ag, and Ag@Glu–TSC NPs, total RNA was extracted using the High Pure RNA Isolation kit (Roche Co., Germany). The RNA was treated with DNase I to remove genomic DNA contaminations. The quantity and quality of RNAs were evaluated by agarose gel electrophoresis and ultraviolet (UV) spectroscopy methods, and reverse transcription was performed with the complementary DNA (cDNA) synthesis kit using 1 μg RNA.
Quantitative PCR
The expression pattern of the pslA gene during biofilm formation in P. aeruginosa isolates was studied before and after treatment with subinhibitory concentrations of TSC, Ag, and Ag@Glu–TSC NPs by a one-step ABI Real-time PCR machine (Applied Biosystems, Foster City, CA). The quantitative PCR (qPCR) amplification was performed in triplicate with primer pairs described in Table 1. The total reaction volume was 20 μL, including 0.5 μL of the cDNA (100 μg/mL) as a template, 0.5 μL of each primer (2 pmol), and 10 μL Power SYBR®Green PCR Master Mix.
Primers Used for Quantitative Polymerase Chain Reaction Amplification
After the initial denaturation step at 95°C for 10 min, the reactions were run for 40 cycles of amplification at 95°C for 20 sec, 60°C for 20 sec, 72°C for 30 sec, and a final extension at 72°C for 5 min. Nuclease-free water was used as a negative control sample. The relative expression levels of the pslA gene were analyzed using ΔΔCт method and the 16s rRNA gene was used as an endogenous control for normalization of the results.
Statistical analysis
The statistical package for social science (SPSS) version 21 statistical software (SPSS, Inc., Chicago, IL) was used for statistical analysis and all data were expressed as mean ± standard deviation (ANOVA, Tukey's HSD test as post hoc test). All experiments performed in triplicate and the differences were considered statistically significant if the p-value was ≤0.05.
Results
Characterization of Ag@Glu–TSC nanoparticles
The UV-visible DRS of Ag@Glu–TSC NPs in the range of 200–1,000 nm is presented in Fig. 1. The strong and broad peaks with a center at 317 nm are attributed to surface plasmon resonance effect of Ag NPs with different particles sizes.

Based on the SEM and TEM images (Fig. 2a, b), Ag@Glu–TSC NPs have approximately spherical morphology in the size range of 40–60 nm, which are suitable for the cell penetration.

SEM
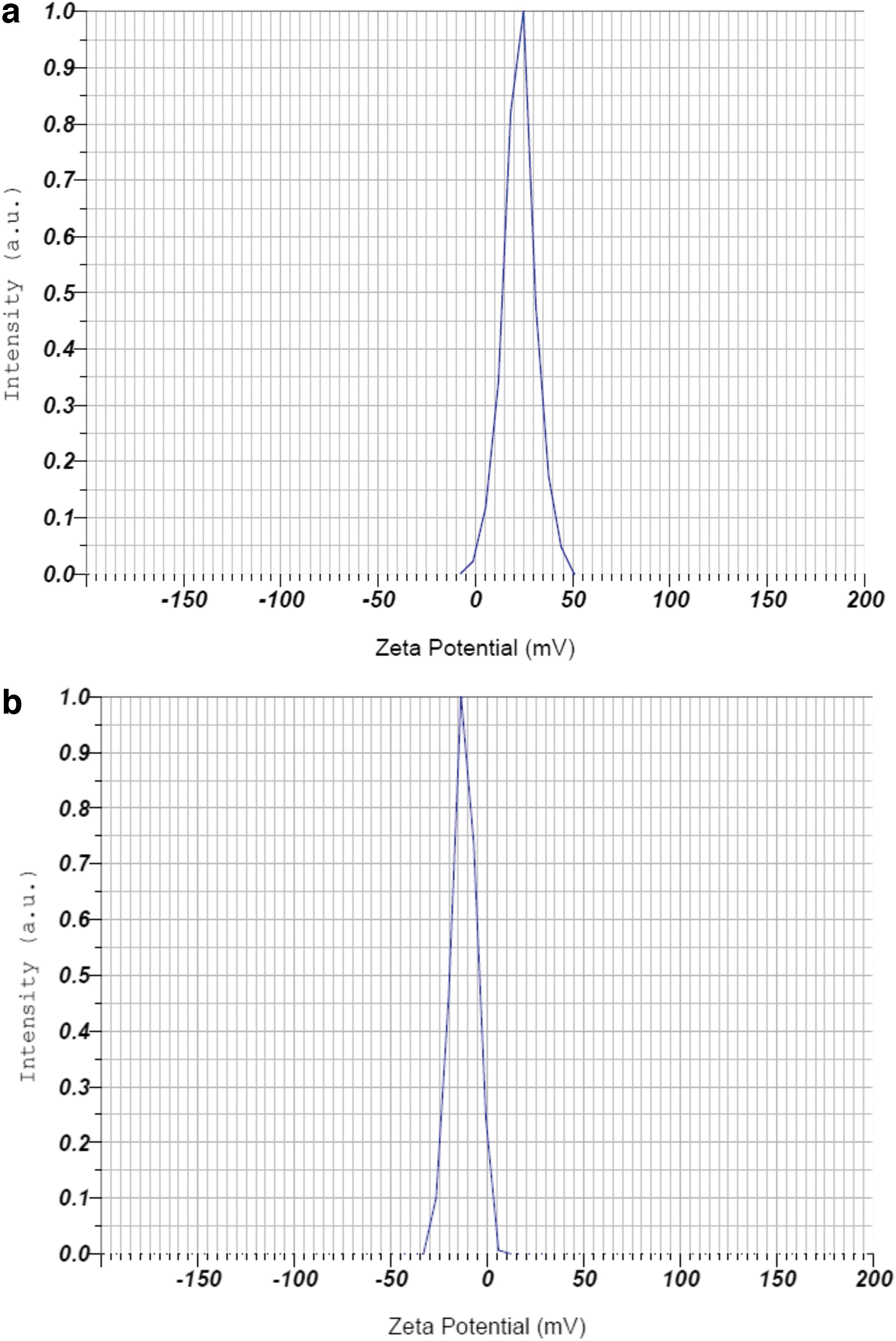
The zeta potential analysis was performed to determine the long-term stability of Ag and Ag@Glu–TSC NPs. The particles with zeta potential values higher than +30 mV or lower than −30 mV are colloidally stable. 35 In contrast, at the isoelectric point, the zeta potential is zero and the colloids are least stable. Herein, the zeta potential value of Ag@Glu–TSC NPs was +32.6 mV, which confirmed the stability of nanoparticles in solution (Fig. 3).
Determination of
Five strong Bragg's reflections peaks at 2θ values of XRD patterns (Fig. 4) are corresponding to [111], [200], [220], [311], and [222] planes that can be indexed to the face-centered cubic phase of Ag in accordance with Joint Committee on Powder Diffraction Standards (JCPDS No. 04-0783). The high intensity and sharpness of XRD characteristic peaks showed the good-quality crystalline structure of Ag@Glu–TSC. In addition, the crystallite size of the Ag@Glu–TSC NPs was estimated according to XRD data using the Debye-Scherer equation (D = K λ/β cosθ). The average size was estimated to be about 45 nm, which is consistent with the results of electron microscopy.

The XRD patterns of the
Energy dispersive X-ray analysis was used to identify the elemental composition of the prepared Ag@Glu–TSC NPs (Table 2). The sharp optical absorption peak approximately at 3 keV confirmed the presence of Ag NPs. Generally, the results clearly identify the presence of Ag, S, N, and O signals in the spectrum (Fig. 5), and confirmed the conjugation of Ag NPs with thiosemicarbazide. The higher intensity of S peak compared with Ag peaks indicates the high surface to volume ratio of Ag NPs and their coating by chemical compounds.

The EDX spectra of
Elemental Composition of Ag@Glu–TSC Nanoparticles Based on the Energy Dispersive X-Ray Analysis Results
The Fourier transform infrared (FT-IR) analysis was performed to determine the chemical composition and functional groups present on the surface of the synthesized Ag@Glu–TSC NPs (Fig. 6). The weak peak situated at 2,962 cm−1 belonged to the O-H stretching and shows the interaction of water with the surface of Ag NPs. The bands around the 1,550–1,650 and 3,250–3,400 cm−1 are corresponding to N–H stretch of 1°, 2° amine, and amide of thiosemicarbazide and glutamic acid. The spectral band around 1,618 and 1,643 cm−1 (C = O vibration of -COOH groups for glutamic acid) indicates the binding of glutamic acid to the surface of Ag NPs.
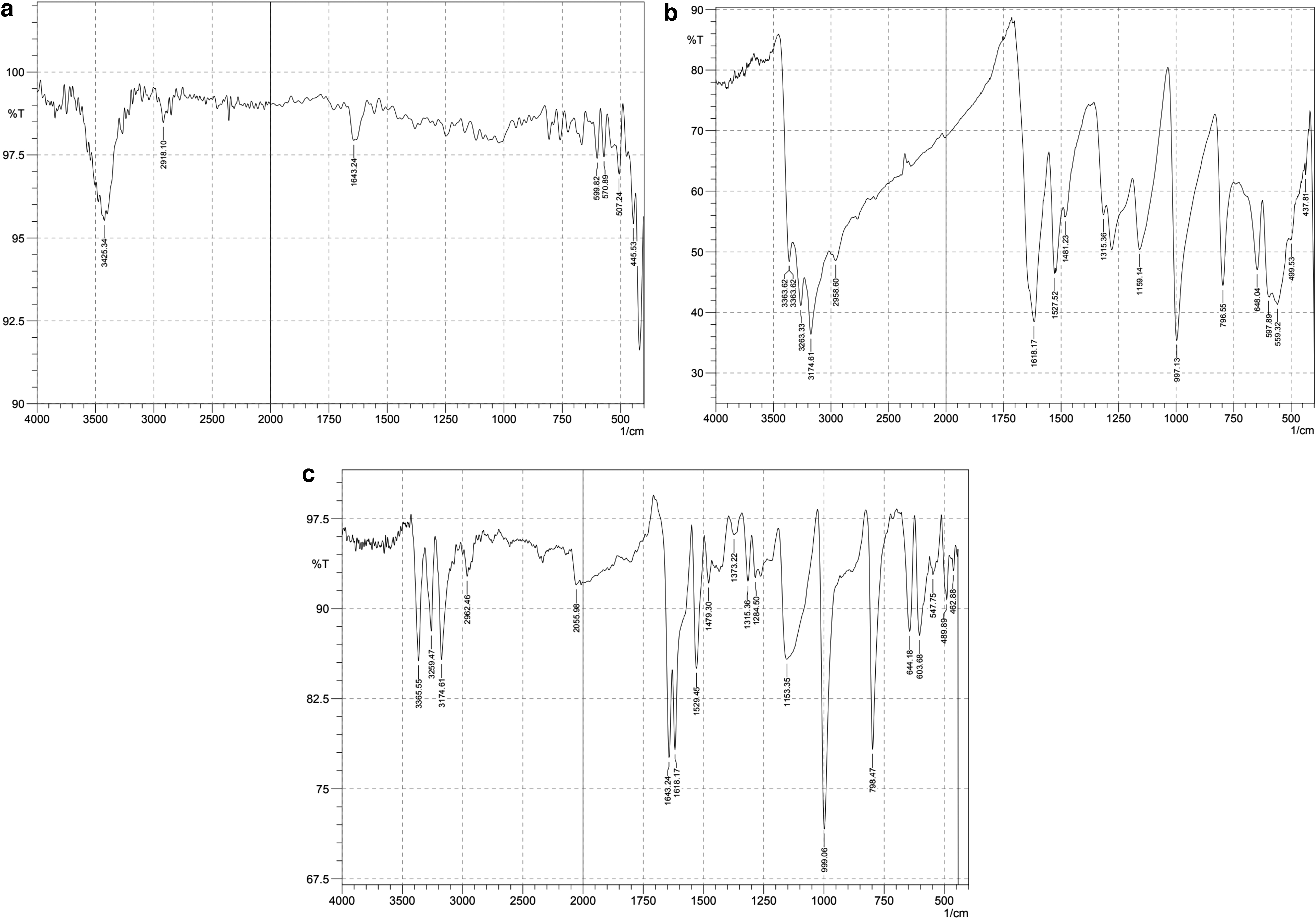
The FT-IR spectra of
The N–O asymmetric stretch at 1,529 cm−1 is corresponding to nitro compounds, which confirmed the presence of glutamic acid and thiosemicarbazide on the surface of Ag NPs. The sharp absorption peaks centered at 1,284 cm−1 is corresponding to the stretching vibrations of the carboxylic acid groups of glutamic acid. In general, the results of FT-IR confirmed the conjugation of thiosemicarbazide to Ag NPs using glutamic acid as a linker.
Antibacterial activity
Resistance profile of P. aeruginosa isolates
Based on the standard zone of inhibition, 100 bacterial samples are classified as sensitive, intermediate, and resistant (Table 3). Of the 100 P. aeruginosa isolates evaluated in this study, almost all isolates were resistant to at least 1 or more antibiotics, 84 isolates exhibited MDR phenotype with resistance to at least 3 antibiotics of different classes, and 29 isolates were resistant to all antibiotics.
Antibiotic Resistance Profile of Pseudomonas aeruginosa Against a Different Group of Antibiotics
Values shown are the percentage of the total (n = 100).
Determination of MIC and sub-MIC
The antibacterial activity of ciprofloxacin, TSC, Ag, and Ag@Glu–TSC NPs was evaluated against multiple antibiotic-resistant isolates of P. aeruginosa by a broth microdilution MIC method. The results showed a dose-dependent inhibitory effect of antibacterial agents against the P. aeruginosa isolates. The MICs for most resistant isolate against ciprofloxacin, TSC, Ag, and Ag@Glu–TSC NPs were 32, 256, 256, and 128 μg/mL, respectively (p < 0.01). A notable observation was greater killing effects of Ag@Glu–TSC NPs than TSC and Ag NPs separately, which demonstrated better antibacterial activity for the combination of both agents (p < 0.01).
Based on our results, the sub-MICs of thiosemicarbazide, Ag, and Ag@Glu–TSC NPs were 1/4 MICs. The biofilm grown on the plates exhibited the highest inhibition of adherence for Ag@Glu–TSC NPs (57%) compared to TSC (17%) and Ag NPs alone (48%). All experiments were performed in triplicate and antibacterial free broth media were used as negative control (p < 0.01).
Biofilm formation and antibiofilm assay
The biofilm production ability in a total number of 100 clinical isolates of P. aeruginosa was evaluated in vitro and the results are shown in (Table 4).
Classification of Pseudomonas aeruginosa Isolates Based on the Biofilm Production Ability
The results of microtiter plate method showed that for all the tested clinical isolates of P. aeruginosa, the sub-MICs of Ag NPs, TSC, and Ag@Glu–TSC NPs inhibited the activity of biofilm formation. The Ag NPs and thiosemicarbazide alone reduce the biofilm activity by ∼45% and 22% in P. aeruginosa isolates, respectively. Combination of Ag NPs and thiosemicarbazide showed more effective biofilm inhibition activity by 67% (Table 5), which revealed the synergistic effect of Ag NPs and TSC.
Mean Percentage of Inhibition of Biofilm Formation in the Presence of Subminimum Inhibitory Concentration of Different Antibacterial Agents
Ag NP, silver nanoparticle.
qPCR of pslA gene
The relative expression of the pslA biofilm gene was quantitatively determined by real-time PCR after treating with sub-MIC (1/4 MIC) of TSC, Ag, and Ag@Glu–TSC NPs. As shown in Fig. 7 the most downregulation of the pslA gene expression was determined for exposure to Ag@Glu–TSC NPs (61%). Antibacterial-free broth media were used as a negative control.
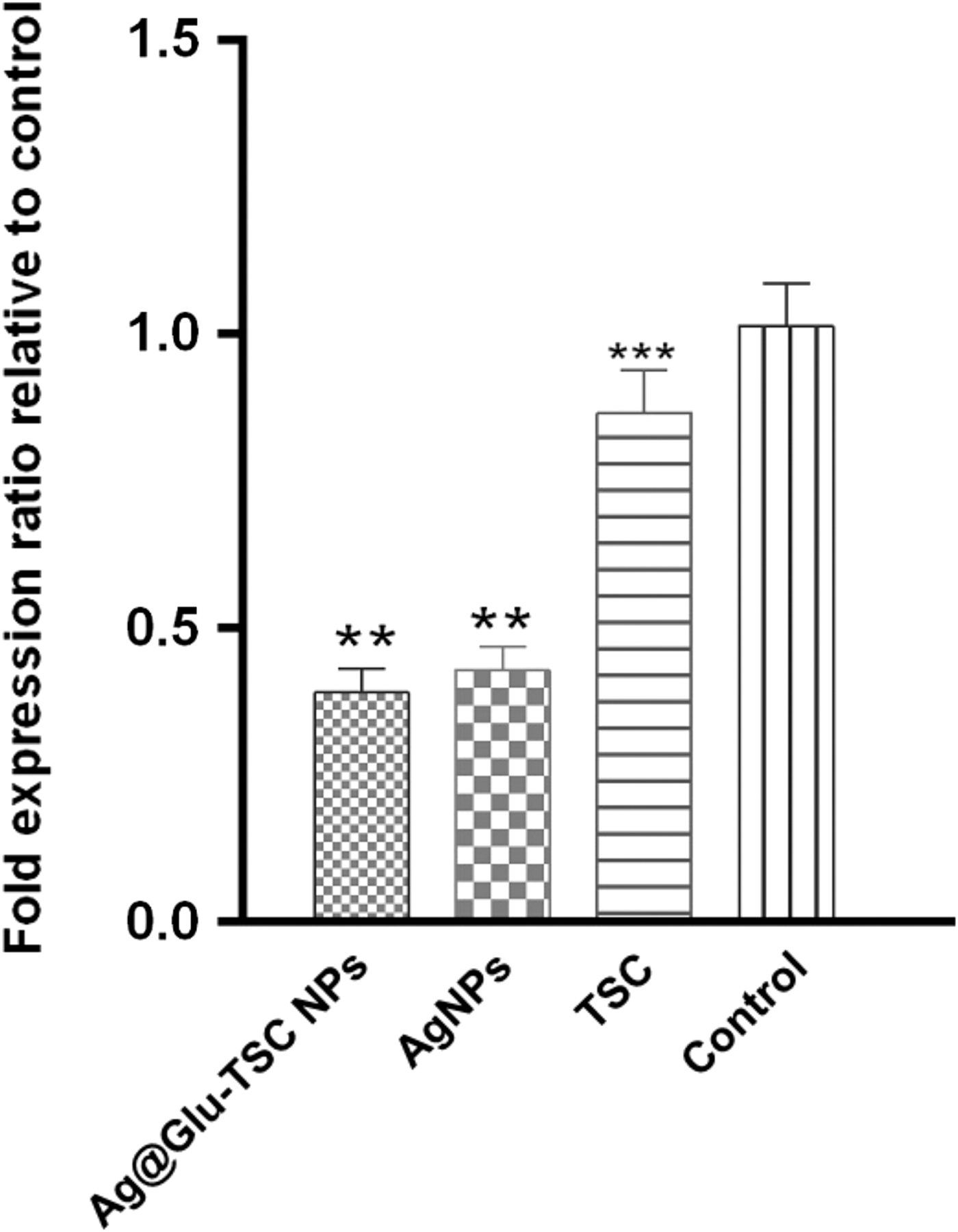
The effect of Ag@Glu–TSC NPs, Ag NPs, and thiosemicarbazide on the relative expression of pslA biofilm gene in Pseudomonas aeruginosa isolates (n = 3, *p < 0.05, **p < 0.01, ***p < 0.001).
Discussion
P. aeruginosa as an opportunistic pathogen exhibits resistance to different classes of antibiotics such as the β-lactam and penem group. Emergence of MDR isolates of P. aeruginosa has been reported, especially in many developing countries worldwide.36–38 Therefore, one of the most important goals of researchers is to find suitable antibiotic alternatives to fight this pathogen. Antibacterial activity of Ag NPs against the planktonic cells and biofilms of Gram-negative bacteria is reported in some articles. For examples, Radzig et al. reported the antibiofilm effects of stabilized Ag NPs on P. aeruginosa PAO1, Serratia proteamaculans 94, and E. coli AB1157. 39
Liao et al. showed the disruption of morphology and structure of MDR P. aeruginosa after entrance of Ag NPs into the cells. 40 Furthermore, the antibacterial activity of green synthesized Ag NPs against a diverse range of Gram-negative foodborne pathogens such as K. pneumoniae, Salmonella Enteritidis, Salmonella Typhimurium, and E. coli was determined by Loo et al. using disk diffusion method. 6
In addition to antibiotics, there are chemicals such as thiosemicarbazide, which can be used to fight bacterial infections. The antibacterial, cytotoxic, and antiproliferative activities of thiosemicarbazide and its derivatives were evaluated and reported in some studies. The results of Pitucha et al. demonstrated good antibacterial properties of some thiosemicarbazide derivatives against Streptococcus mutans, Staphylococcus epidermidis, and Streptococcus sanguinis with only slightly cytotoxic effects on host cells. 41 Antibacterial activity of thiosemicarbazide derivatives has been also reported by researchers such as Kalhor et al. and Wujec et al.27,42
In our previous studies, we evaluated the antibacterial properties of some metal nanoparticles alone and in combination with thiosemicarbazide to find the best combination against antibiotic-resistant bacteria. In this study, the antibacterial properties of Ag NPs functionalized with thiosemicarbazide were examined to study their synergistic effects against biofilm formation in antibiotic-resistant isolates of P. aeruginosa collected from infected patients.
Based on the results of the antimicrobial susceptibility testing, 84% of the P. aeruginosa isolates were resistance to at least three antibiotics of different classes (MDR isolates). The MICs of Ag NPs as well as thiosemicarbazide (up to 256 μg/mL) against control and clinical P. aeruginosa isolates, were higher than Ag@Glu–TSC NPs (128 μg/mL). This means that Ag@Glu–TSC NPs are more useful for fighting MDR strains compared to Ag NPs and thiosemicarbazide (alone). Previously, our research team evaluated the antibacterial activity of Ag@Glu–TSC NPs against methicillin-resistant Staphylococcus aureus (MRSA) strains as a Gram-positive pathogen. The MIC value of TSC and Ag NPs against MRSA determined 1,024 μg/mL, while the MIC value of Ag@Glu–TSC against clinical strains determined in the range of 256–512 μg/mL. 43
Currently, there are some reports on the antibiofilm potential of Ag NPs and nanocomposites. Kalishwaralal et al. showed more than 95% inhibition of biofilm formation in P. aeruginosa and S. epidermidis strains after treatment with Ag NPs. 44 In another research, Ramalingam et al. confirmed the antibiofilm activity of Ag NPs against P. aeruginosa and S. aureus. 45 Gurunathan et al. tested the inhibitory ability of Ag NPs against biofilm formation by the human pathogens P. aeruginosa, S. aureus, Streptococcus pneumoniae, and Shigella flexneri under in vitro conditions. They presented the ability of Ag NPs to inhibit biofilm activity in a dose-dependent action and at concentrations slightly lower than those that affected host cell viability. 46
We have also previously shown that biofilm formation in MRSA can be inhibited up to 76.7% in the presence of subinhibitory concentrations of Ag@Glu–TSC NPs. 43 In agreement with these reports, in this study, our finding showed a decrease in biofilm formation after treatment with Ag NPs, and this reduction was enhanced by the conjugation of Ag NPs to thiosemicarbazide up to 70% compared with the control.
In P. aeruginosa, the psl gene operon comprises 12 genes, which encode PslA-PslO proteins to synthesize Psl exopolysaccharide that plays an important role in biofilm formation and maintenance.18,19 Based on the results of Billings et al., during the early stages of biofilm development, the Psl polysaccharide provides a first-line defense toward antibiotics. 47 The contribution of Psl exopolysaccharide in the tolerance and resistance to antibiotic treatment in P. aeruginosa was reported in some other studies.48–50 Furthermore, Mishra et al. showed that the Psl polysaccharide could protect P. aeruginosa cells against neutrophil-mediated phagocytosis and oxidative stress. 51 These studies clearly confirmed the functions of Psl as a scaffold for biofilm development and a barrier against host immune defense and antibiotic attacks. Therefore, preventing the expression of Psl operon genes can provide an effective way to inhibit biofilm production in P. aeruginosa.
The results of qPCR showed the downregulation of pslA gene in the presence of Ag@Glu–TSC NPs by an average of 61%, compared to the control sample. The precise inhibitory mechanisms of nanoparticles toward the gene expression have not been fully characterized, but the decrease in pslA gene expression could be due to transcription blocking by reactive oxygen species 52 or direct interaction of the nanoparticles with the transcription factors.53,54 Before this, the effect of thiosemicarbazide, Ag NPs, and Ag@Glu–TSC nanoparticles on the expression of biofilm-associated genes icaA and icaD was investigated in S. aureus and highest reduction was observed in the presence of Ag@Glu–TSC NPs. 43
The agreement of results obtained in two studies shows the effective performance of Ag@Glu–TSC nanoparticles in the reduction of the expression of biofilm formation genes in both Gram-negative and Gram-negative bacteria compared to thiosemicarbazide and Ag NPs alone. According to these findings, the enhanced synergistic effect of Ag NP nanoparticles in combination with thiosemicarbazide against pathogenic bacteria may be used as a potent antibiotic alternative to fight a variety of bacterial infections.
Conclusion
The antibacterial properties of Ag NPs have been known for many years. Furthermore, the conjugation of Ag NPs with antimicrobial agents such as antibiotics and other organic compounds such as thiosemicarbazide has been considered to improve their activity in clinical applications. Accordingly, in this study, we have synthesized Ag NPs functionalized by glutamic acid and conjugated with thiosemicarbazide to evaluate their effects on the expression of pslA biofilm genes in multidrug-resistant P. aeruginosa isolates. In conclusion, we found that 40–60 nm Ag@Glu–TSC NPs can penetrate the bacterial cell and decrease the pslA gene expression that results in the inhibition of biofilm formation.
Therefore, the Ag@Glu–TSC NPs can be considered a suitable candidate for fighting multidrug-resistant microbial pathogens. However, during the use of these substances, like antibiotics, environmental points must be considered and observed. Nevertheless, more investigations are required to determine the side effects of these nanoparticles on the host cells.
Footnotes
Authors' Contributions
Mrs. Honarmand and Dr. Salehzadeh are from Islamic Azad University (Rasht, Iran), Dr. Jalali is from Arak University (Arak, Iran), Dr. Nikokar is from Guilan University of Medical Sciences (Rasht, Iran), and Dr. Panahi Sharif from Ondokuz Mayis University (Samsun, Turkey), all where education and research are the primary functions.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.