
Abstract
Conjugative plasmids are a major contributor to the global spread of antibiotic resistance determinants, but the tracking of their evolutionary history is often neglected. Salmonella enterica serovar Typhimurium (S. Typhimurium) strain SRC27 was isolated from an equine infection in Australia in 1999. SRC27 was known to carry conjugative HI1 and I1 resistance plasmids. In this study, SRC27 was sequenced to determine the relationship between these HI1 and I1 resistance plasmids it was known to carry and HI1 and I1 resistance plasmids circulating worldwide. The resistance genes in the HI1 plasmid, pSRC27-H, are all located in a single complex 34.7 kb resistance region. The backbone sequence and location of the pSRC27-H resistance island were used to identify the most closely related HI1 plasmids among the >90 that have been sequenced since 2011. This defined a sublineage of 20 type 2 HI1 plasmids that have been circulating in Europe, Asia, North America, and Australia since at least 1993. The overall resistance gene content of these HI1 plasmids differs, indicating extensive evolution in situ through the acquisition of additional transposons and deletion or replacement of ancestral regions. The I1 plasmid contains a complete copy of Tn5393a, containing the strAB genes that confer resistance to streptomycin. The precise location of Tn5393a in the backbone also defined a globally disseminated sublineage of I1 plasmids, many of which have also acquired additional resistance determinants. The sequence revealed that SRC27 also carried two additional plasmids, the pSLT-type FIB(S):FII(S) virulence plasmid and a small cryptic theta-replicating Col156 plasmid.
Introduction
Nontyphoidal serovars (NTSs) of Salmonella enterica are a leading cause of foodborne illness leading to hospitalizations and deaths worldwide. S. enterica serovar Typhimurium (S. Typhimurium) is an NTS that causes inflammatory diarrhea that can lead to invasive systemic disease in vulnerable populations. 1 Multiple drug resistant (MDR) strains of S. Typhimurium first emerged in the 1980s and are now dominant in many regions. 2 A variety of different plasmid types have been reported in S. Typhimurium, although conjugative plasmids of the IncHI1 incompatibility group account for many of the resistance determinants seen in MDR S. Typhimurium in the United States. 2 R27, isolated in the United Kingdom in 1961 from a tetracycline-resistant S. enterica serovar Typhi (S. Typhi), is the archetypal IncHI1. It has been extensively studied,3–5 and in 2000, was the first IncHI1 plasmid to be sequenced. 6 The well-characterized backbone includes two regions required for conjugative transfer (Tra1 and Tra2), two partitioning modules (Par1 and Par2), two functional replication regions named repHI1A and repHI1B, and a nonfunctional repFIA.4,6 Recent studies have identified further regions of the IncHI1 backbone required for stability and replication. 7
By 2011, eight IncHI1 plasmids had been sequenced (R27, pHCM1, pAKU_1, pMAK-1, pO111_1, p9804_1, p7467_1, and p6979_1). 8 Two types of IncHI1 plasmids were later defined based on variable backbone regions and the location and structure of their resistance islands. 9 Type 1 includes the archetypal IncHI1 plasmid R27, while type 2 included pHCM1, pO111_1, and pMAK1.
S. Typhimurium strain SRC27 was isolated from an equine infection in Australia in 1999. 10 Using a combination of conjugation experiments and PCR-based replicon typing (PBRT), it was shown to carry the first known example of an IncHI1 plasmid from Australia, named pSRC27-H, and was also the first from an animal source. 10 pSRC27-H carried the aacC2 (gentamicin), aadA2 (streptomycin and spectinomycin), aphA1a (kanamycin and neomycin), blaTEM-1 (ampicillin), catA1 (chloramphenicol), dfrA12 (trimethoprim), strAB (streptomycin), sul2 (sulfamethoxazole), and tet(B) (tetracycline) resistance genes. At the time it was the only IncHI1 plasmid known to carry a gentamicin resistance determinant. 10 Linkage PCR combined with Sanger sequencing was used to show that all of the resistance determinants were located in a single complex multiple antibiotic resistance region (GenBank accession no. HQ840942). In addition, PBRT and conjugation experiments also demonstrated that SRC27 contained an I1 plasmid, pSRC27-I, that conferred resistance to streptomycin and spectinomycin (strAB). 10 Both pSRC27-H and pSRC27-I are conjugative, transferring at frequencies of 5 × 10−7 and 1 × 10−4, respectively, at 37°C. HI1 plasmids transfer poorly at 37°C, and conjugation at 26°C increased the transfer frequency of pSRC27-H over 1,000 fold. 10
Relative to other important contributors to the spread of antibiotic resistance, such as the IncC plasmids of which >350 have been sequenced and analyzed, IncHI1 plasmids remain relatively uncommon in sequenced genomes and the relationships between most of the sequenced IncHI1 plasmids have never been investigated. The absence of any other sequenced IncHI1 plasmid of Australian origin in the public database presents a unique opportunity to further our understanding of this important contributor to the global spread of antibiotic resistance. In this study, with the availability of a complete genome sequence of SRC27, I aimed to investigate the overall genetic composition of pSRC27-H and the other plasmids carried by SRC27 and to reexamine the evolutionary relatedness to other globally distributed epidemic MDR strains of S. Typhimurium using the larger set of genome data available today.
Materials and Methods
Bacterial strain and preparation of DNA
S. enterica serovar Typhimurium strain SRC27 was isolated in 1999 from an equine infection in Australia. The antibiotic resistance profile of SRC27 was confirmed using a disc diffusion method, as described previously, 10 with an annular radius of <6 mm indicative of resistance. The resistance profile of SRC27 was further confirmed by patching on plates containing a suitable level of the antibiotic as described previously, 10 using Escherichia coli UB1637 as a control.
Genome sequencing
The complete genome of strain SRC27 was determined by sequencing using long-read MinION (Oxford Nanopore) and short-read Illumina MiSeq technologies. Genomic DNA was prepared as described previously. 11 The MinION run (195 × depth) generated a total of 52,338 reads, and the Illumina MiSeq run (32 × depth) generated a total of 456,791 150-bp paired-end reads. The MinION reads were filtered using Filtlong (version 0.2.0) to remove any reads <1,000 bp, followed by removal of the lowest 10% of reads by quality. The long and short reads were combined in a hybrid assembly using Unicycler (version 0.4.0) 12 with default assembly parameters.
Protein coding, rRNA, and tRNA genes were annotated using a combination of Prokka (version 1.12) 13 and RAST. 14 Plasmid replicons and antibiotic resistance genes were identified using ABRicate (version 0.8.13) and were annotated manually. Insertion sequences (IS) were identified using the ISfinder database 15 and annotated manually. The PubMLST database was used to determine the sequence type of SRC27 and to subtype the plasmid replicons where possible. The pSRC27-H sequence was opened in the same location as the R27 sequence (GenBank accession no. AF250878), and the pSRC27-I sequence was opened in the same location as the R64 sequence (GenBank accession no. AP005147). Related plasmids were identified using a standard BLAST search and filtering manually for complete plasmid sequences. Plasmid copy number was determined by comparing the coverage of the individual plasmid molecules, as reported in the Unicycler assembly, with the coverage of the circular chromosome.
Nucleotide accession numbers
The SRC27 chromosome and plasmids pSRC27-Col156, pSRC27-F, pSRC27-H, and pSRC27-I have been deposited in GenBank under accession numbers CP058807, CP058808, CP058809, CP058810, and CP058811, respectively. The existing GenBank entry for the pSRC27-H resistance region (HQ840942) was updated to incorporate the complete manually annotated plasmid backbone. The Illumina HiSeq and Oxford MinIon reads for this project have been deposited in the Short Read Archive under accession nos. SRX8685365 and SRX8685366, respectively.
Results
SRC27
The hybrid assembly of strain SRC27 from short- and long-read sequence data generated five circular molecules (Table 1) representing the SRC27 chromosome (4,865,373 bp) and four plasmids ranging in size from 3,752 to 205,772 bp. SRC27 was typed as ST19 [aroC (10), dnaN (7), hemD (12), hisD (9), purE (5)] using the S. enterica MLST scheme in the PubMLST database. ST19 is the most prevalent S. Typhimurium sequence type (ST) internationally. 16 The two previously identified HI1 (pSRC27-H) and I1 (pSRC27-I) plasmids were present. 10 The ABRicate output identified two additional plasmids that had not been detected previously; an FIB(S):FII(S) plasmid and a small high-copy number Col156 plasmid (Table 1).
Genetic Content of SRC27
Copy number determined relative to the SRC27 chromosome.
The 93,853 bp FIB(S):FII(S) plasmid did not contain any antibiotic resistance genes and was identified as the pSLT-type (also previously called MP10, pRQ28, or pSTV) virulence plasmid17,18 commonly found in S. Typhimurium strains from around the world. 19
pSRC27-H
pSRC27-H is a low-copy (∼2 copies per cell, Table 1) 205,772 bp conjugative IncHI1 plasmid (Fig. 1A) and was further subtyped as ST2 plasmid using the PubMLST database. The assembly of plasmid pSRC27-H confirmed that the catA1 (chloramphenicol), sul2 (sulfonamides), strA/strB (streptomycin), aphA1a (kanamycin and neomycin), aacC2 (gentamicin), dfrA12 (trimethoprim), aadA2 (streptomycin and spectinomycin), tet(B) (tetracycline), and two copies of blaTEM-1 (ampicillin) resistance genes are contained in a single 34,730 bp complex resistance region that is bounded at one end by IS1 and at the other end by IS10 (Fig. 1B), as reported previously. 10
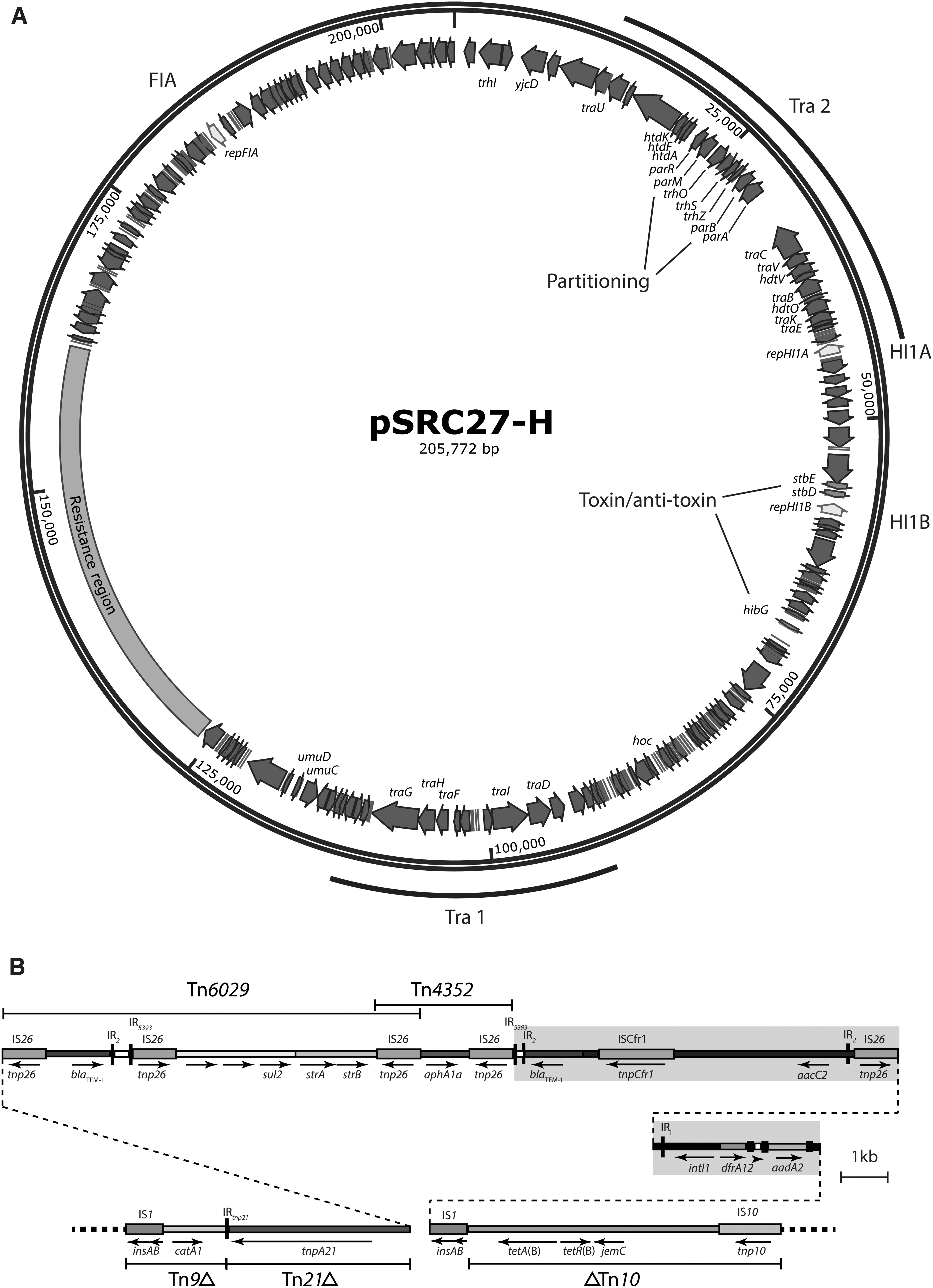
The backbone of pSRC27-H was derived by removing the complex resistance island (Fig. 1B), generating a backbone of 171,042 bp. The pSRC27-H backbone contains 198 open reading frames (ORFs) encoding proteins of >50 amino acids (aa). This includes the repHI1A and repHI1B replication initiation genes and 20 genes involved in plasmid maintenance, including the parA and parB partitioning genes and the higB toxin gene. pSRC27-H also includes the repFIA gene, found in most IncHI1 plasmids surrounded by two copies of IS1. While this gene, derived from FIA F plasmids, is nonfunctional for replication of IncHI1 plasmids, it has been previously shown to confer incompatibility with FIA plasmids. 20 Both the Tra 1 and Tra 2 transfer regions are present in pSRC27-H, and all of the genes previously shown to be essential for conjugative transfer of IncHI1 plasmids are accounted for.5,6 Although over 90 IncHI1 plasmids have been sequenced and deposited in the GenBank nonredundant database (Supplementary Table S1), pSRC27-H is still the only IncHI1 plasmid from Australia.
Related IncHI1 plasmids
IncHI1 plasmids have previously been grouped into two types based on three distinct backbone segments (“Region C,” “Region D,” and “Region E”) and the location and structure of the resistance island (“Region A” and “Region B”). 9 However, even as early as 2012, it was noted that some IncHI1 plasmids were emerging that did not strictly fit within the groups defined by these features. 9 In this study, the repHI1A and repHI1B genes of pSRC27-H were used to query the GenBank nonredundant database to recover all of the IncHI1 plasmids sequenced to date. Ninety other IncHI1 plasmids were found, and the metadata for them was collated (Supplementary Table S1). A backbone comparison using the pSRC27-H backbone as the query showed that among the 90 sequenced plasmids, 24 others (Table 2), including the plasmids previously classified as type 2 IncHI1 (pHCM1, pO111_1, pMAK1), 9 share a conserved backbone (>99.5% nucleotide identity and >98% coverage) with pSRC27-H. Twenty of these 24 plasmids also include a resistance island in the same location as the one in pSRC27-H (see below). The type 2 IncHI1 group had previously been predicted largely based on the presence of Region C, Region D, and Region E. 9 This grouping is now further enhanced by a highly conserved backbone sequence. However, some minor backbone variations result in some different sequence types using the IncHI1 pMLST scheme (Table 2). The vast majority share ST2 with pSRC27-H, but the oldest plasmid pHCM1 is ST1 and three other plasmids are ST9. This highlights the importance of not relying solely on ST to group plasmids, as close evolutionary relationships may be missed.
Plasmids Related to pSRC27-H
ST determined using the IncHI1 pMLST scheme in the PubMLST database.
NT, nontypeable; ST, sequence type.
The related plasmids were recovered from E. coli and various S. enterica serovars in human, animal, and environmental sources across a broad geographic range, including Australia, North America, Europe, Asia, and South-East Asia (Table 2). This is indicative of a global spread of type 2 IncHI1 plasmids. There is also a clear epidemic persistence of this plasmid type, as it has been circulating since pHCM1 was first recovered 27 years ago (Table 2). The four plasmids with a conserved type 2 backbone, but without the related resistance island (pT16R-1, p1919D62-1, p2019XSD11-TC2-284, and p2019XSD11-190), were all recovered from animal sources in China in 2019 and may represent another sublineage or an ancestral lineage of type 2 IncHI1 plasmids.
The pSRC27-H resistance region and related plasmids
The 20 other IncHI1 plasmids that contain a resistance region in the same backbone location as the resistance region in pSRC27-H (Table 2) include some of the earliest IncHI1 plasmids to be sequenced (pHCM1, pO111_1, and p109/9). The resistance gene content and structure of the resistance islands are variable (Table 3), but they are all based on the same general transposon scaffold and all-but-one contains the tet(B) tetracycline resistance gene derived from Tn10. Fifteen of the plasmids share the same left and right resistance island boundaries with pSRC27-H, and the boundaries in the remaining five differ only due to IS26-mediated deletions extending left (relative to Fig. 1B) from within the resistance island into the plasmid backbone. pEQ1, pEQ2, and p3498 share the same 187 bp IS26-mediated backbone deletion, while RCS78 and pP59A-CTX-M-55 have lost 1,032 and 3,369 bp from their backbones, respectively, due to IS26-mediated deletions. The complex internal resistance gene rearrangements seen in some of the resistance islands are the result of evolution in situ through the acquisition of various transposons and other mobile genetic elements.
Resistance Gene Content of Plasmids Related to pSRC27-H
Left boundary of resistance island missing due to IS26-mediated deletion.
Two separate resistance islands. Second contains tet(M) and the second copy of blaTEM-1.
p109/9 (GenBank accession no. KP899805) and pO111_1 (GenBank accession no. AP010961) have an identical 29,474 bp resistance island, containing the catA1, blaTEM-1, sul2, strA, strB, aphA1, and tet(B) resistance genes, that is most closely related to the island in pSRC27-H. The segments of difference between pSRC27-H, p109/9, and pO111_1 are shaded in Fig. 1B. In pSRC27-H, the complex mosaic structure between Tn4352 and IS1 replaces the mer end of Tn21 in p109/9 and pO111_1 (see Fig. 1 in Cain and Hall 10 ).
In 2012, pSRC27-H was the only IncHI1 plasmid known to confer gentamicin resistance, owing to the presence of the aacC2 gentamicin resistance gene, 10 but there are now six other related IncHI1 plasmids that contain aacC2 (RCS78_p, pEQ1, pEQ2, p014ODMR, pP59A-CTX-M-55, and p3498, Table 3) in a similar configuration to the one seen in pSRC27-H. All six were recovered from E. coli from animal hosts (three from horses, two from pigs, and one from a dog) in Europe and South-East Asia, indicative of global spread of this sublineage of IncHI1 plasmid containing the aacC2 gentamicin resistance determinant. SRC27 was recovered from an equine infection, and while gentamicin is not usually used in other animals, it is used in combination with ampicillin as a first-choice treatment for Salmonella infections in horses due to their high economic value 21 and the prevalence of MDR S. enterica strains that are already resistant to many other veterinary antibiotics. Five of the eight aacC2-containing plasmids have also acquired the blaCTX-M gene (Table 3). Although different alleles are present, blaCTX-M is located in the same position in the resistance island in the five plasmids, indicative of a single acquisition event followed by the acquisition of single nucleotide polymorphisms (SNPs) to give new alleles.
A sublineage of six plasmids (pPNCS015054_S2, pPNCS014881.1, pPNCS007087.1, p10-3184.1, Unnamed1, and p08-4425.1) related to pSRC27-H was all recovered from human sources in Canada between 2008 and 2011 (Table 2) and contain only the tet(B) resistance determinant (Table 3). Analysis of the structure of their resistance islands showed that recombination between the two directly-oriented copies of IS1 (Fig. 1B) had removed everything between the two IS. This is another example of an evolution in situ event that can be used to define a sublineage of related plasmids circulating in clinically-important isolates that could not have been differentiated from the rest of this lineage based solely on ST.
pSRC27-I
pSRC27-I is a 90,483 bp I1 plasmid that was further subtyped as ST302 (repI1 (1), ardA (3), trbA (3), sogS (6), pilL (10)) using the I1 MLST scheme 22 in the PubMLST database. pSRC27-I is a low-copy plasmid, existing at ∼3 copies per chromosome (Table 1). pSRC27-I contained the strA and strB streptomycin resistance genes, consistent with previous linkage PCR that had shown that the strAB genes in pSRC27-I were linked to IS1133 (incorrectly named as IS1331) on one side and to the tnpR gene of Tn5393 on the other, 10 although the overall genetic context of the strAB genes had not been determined.
Analysis of the complete pSRC27-I sequence showed that the strAB genes are located in a complete copy of Tn5393a (Fig. 2A), a 6,706 bp variant of Tn5393 that includes a copy of the IS3 family member IS1133.23,24 Tn5393a in pSRC27-I is flanked by a 5 bp (TATAA) target side duplication (TSD), indicative of a clean insertion into the pSRC27-I backbone with no adjacent deletions. No other insertion sequences or other mobile genetic elements were identified in pSRC27-I. Removal of Tn5393a and one copy of the 5 bp TSD generated a backbone of 83,772 bp with a complete transfer region (Tra in Fig. 2A) that contains all of the genes previously shown to be essential for conjugative transfer of I1 plasmids. 25 This is consistent with the previously reported ability of pSRC27-I to transfer from S. Typhimurium to E. coli at a frequency of 1.4 × 10−4 transconjugants per donor. 10

Circular maps of
Searches using backbone of pSRC27-I to other sequences in the GenBank nonredundant database revealed at least 250 related I1 plasmids of various sizes and resistance gene contents. I1 plasmids exhibit high levels of mosaicism, which can make evolutionary lineages difficult to deduce.26,27 For example, regions in pSRC27-I and the archetypal I1 plasmid R64 share 86.2–99.8% nucleotide identity. As the acquisition of a resistance island represents a distinct ancestral evolutionary step, the position of a resistance island can be used as a marker of a lineage. Six other I1 plasmids (Table 4) have Tn5393a, or a variant of Tn5393, in the same backbone location as Tn5393a in pSRC27-I. In one instance (pCV839-15-2, GenBank accession no. CP024976), the strA and strB belonging to Tn5393 have been removed by a deletion. The I1 plasmids containing Tn5393a in this specific backbone location represent a globally distributed sublineage of I1 plasmids found in S. enterica and E. coli from Australia, Asia, Europe, North America, and the Middle East over a three-decade period (Table 4). However, the six plasmids with Tn5393 also all contain additional antibiotic resistance genes located in other backbone locations (Table 4). These plasmids also all exhibit significant mosaicism in their backbones, with regions that share 95–99% nucleotide identity, which would make this lineage difficult to identify without using the resistance island as an epidemiological marker.
Plasmids Related to pSRC27-I
Additional resistance genes located elsewhere.
Hybrid I1 and FIB:FII:FIC plasmid.
A deletion has removed the strA and strB genes belonging to Tn5393.
A small theta-replicating plasmid in SRC27
The smallest plasmid identified in SRC27 was a high copy number (38 × , Table 1) 3,752 bp Col156 plasmid (Fig. 2B). Although Col156 plasmids have previously been recognized as important vehicles for the transmission of antibiotic resistance determinants,28,29 pSRC27-Col156 does not contain any antibiotic resistance genes. pSRC27-Col156 contains three open reading frames. The first is predicted to encode a plasmid replication protein (Pfam PF03090), consistent with the prediction that Col156 plasmids are mobilizable theta-replicating high copy plasmids that use a ColE2-like replication mechanism, involving post-transcriptional control of the plasmid rep gene by plasmid-encoded RNA (RNAI). 30 The other two are predicted to encode a MobA cis-acting relaxase (Pfam PF03389) of the MOBQ4 family 30 and a MobC relaxase accessory protein.
pSRC27-Col156 is not identical to any other sequenced plasmid in the GenBank nonredundant database, and the five closest relatives (pTB505, GenBank accession no. CP034824; pSA20011914.2, GenBank accession no. CP029994; p931-3904, GenBank accession no. CP016386; pSTY2-1899, GenBank accession no. CP019895; pCROD3, GenBank accession no. FN543505) only share 96.1–99.85% nucleotide identity with pSRC27-Col156 and are between 152 and 158 bp larger. The related plasmids were recovered from a number of S. enterica serovars and a Citrobacter rodentium.
Discussion
In the genomic era, the study of older strains is often neglected in favor of large-scale mass sequencing of contemporary antibiotic-resistant isolates. This is the case with S. Typhimurium, where the vast majority of sequences in the public databases are from isolates recovered over the past decade and a clear sequencing bias toward sequencing multiple isolates from very few geographic locations and sources (Supplementary Table S1). In Australia, almost all of the sequenced S. Typhimurium strains were recovered after 2005, 16 and the plasmids carried by Australian S. Typhimurium strains have been largely ignored in favor of core-genome analysis and prophage profiling. 31 Here, the complete sequence of S. Typhimurium strain SRC27, recovered from an infected horse in Australia in 1999, provides a unique perspective into S. Typhimurium from Australia. SRC27 belongs to ST19, the most dominant globally distributed S. Typhimurium ST, 16 and the carriage of four plasmids belonging to the IncHI1, I1, FIB(S):FII(S), and Col156 groups makes SRC27 one of the more prolific carriers of plasmids among S. Typhimurium worldwide.
pSRC27-H belongs to a specific sublineage of IncHI1 plasmids that contains some of the earliest-recovered multiresistant IncHI1 plasmids. pSRC27-H alone can account for all of the resistance phenotypes detected for SRC27, with a second redundant copy of strA/strB located on pSRC27-I. The antibiotic resistance regions carried by plasmids are often complex mosaics, composed of various transposons, insertion sequences, integrons, and other mobile genetic elements carrying a suite of antibiotic resistance genes. Many of these regions have arisen as a result of ongoing evolution in situ32,33, in which a single ancestral insertion of a mobile genetic element at a specific location provides a scaffold for the acquisition and/or loss of additional mobile genetic elements and resistance genes. The evolutionary relationship between the antibiotic resistance island of pSRC27-H and related plasmids was first noted in 2012 when only two related plasmids had been sequenced. 10 Since then, the availability of additional sequences (Tables 2 and 3) demonstrates that evolution in situ within the resistance island has resulted in the recruitment of additional resistance genes, including those conferring resistance to third-generation cephalosporins (blaCTX-M-1 or blaCTX-M-55) and erythromycin [mph(A)] in the last decade. These plasmids are now epidemic, having been circulating worldwide for multiple decades among bacteria of human and animal origin. The specific location of the resistance island is an important epidemiological marker, as it serves to identify this lineage of related plasmids that may otherwise be missed by simply comparing the sequence of the conserved backbones. The ability to identify and track IncHI1 plasmids from this epidemic lineage is of significant clinical importance, as they also account for many of the resistance determinants seen in globally-distributed MDR Salmonella Typhi, 8 the causative agent of Typhoid fever.
The mobilization of pSRC27-Col156 could not be tested owing to it being a cryptic plasmid with no selectable marker. However, an analysis of strains containing Col156 plasmids in the GenBank nonredundant database shows that in >90% of cases, the Col156 plasmid is found with a coresident F plasmid. In some cases the coresident F plasmid is the pSLT-type FIB(S):FII(S) virulence plasmid, as seen in SRC27, and in other cases it is a different F type plasmid. In contrast, Col156 plasmids are rarely found with coresident I1 or HI1 plasmids. Hence, it is possible that it may be mobilized by the pSLT FIB(S):FII(S) virulence plasmid or other related F plasmids.
It is surprising, given the importance of MDR plasmids, that pSRC27-H is still the only IncHI1 plasmid sequenced from Australia in the GenBank nonredundant database. The largest comparative genomic analysis of Australian S. Typhimurium isolates to date 16 unfortunately only mapped reads for specific plasmid types, namely the pSLT virulence plasmid, an I1 plasmid, and a pRSF1010-like plasmid, rather than examining the broader plasmid content of the 39 isolates sequenced. The sequencing and detailed analysis of the overall genetic content of additional historic and contemporary Australian S. Typhimurium isolates will be an important step in understanding the local and international phylogeny of this important pathogen. It will also help to balance the sequencing bias that exists in the public database, which is currently dominated by S. Typhimurium isolates from the United States and Asia.
Footnotes
Acknowledgments
The author thanks Prof. Ruth Hall (The University of Sydney) for supplying isolate SRC27 and for providing support with the preparation of this article. The author also thanks Dr. Mohammad Hamidian (University of Technology Sydney) for bioinformatic assistance.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this work.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.