
Abstract
Objectives:
To characterize the antimicrobial resistance and virulence of pathogenic Escherichia coli isolated from diseased captive giant pandas.
Methods:
Antimicrobial susceptibility and minimum inhibitory concentration (MIC) were determined by the broth dilution method. Whole-genome sequencing was used to characterize the phylogeny, serotype, virulence, resistome, plasmids, and genetic structures of the cefotaxime (CTX)-M genes.
Results:
Four extended-spectrum beta-lactamase (ESBL)-producing E. coli strains were identified and the MICs against 11 antibiotics in vitro were determined. All ESBL-producing E. coli strains were resistant to more than eight antibiotics and carried the blaCTX-M-55 or blaCTX-M-105 gene in different sizes of replicon-type plasmids (pAMSH1-IncHI2, 257 kb; pAMPD2-IncFII, 89 kb; pAMPD02-IncFIB, 129 kb; and pAMSC4-IncN, 47 kb). Distinct insertional sequences and transposases were identified up-/downstream of blaCTX-Ms, including IS26, ISEcp1, ISKpn72, IS903B, and Tn2. These strains also possessed at least three virulence genes of pathogenic E. coli and originated in four different evolutionary branches. One strain carried the complete locus of the enterocyte effacement pathogenicity island, but lacked the virulence genes stx and bfpA, indicating atypical enteropathogenic E. coli, whereas the other strains were considered to be extraintestinal pathogenic E. coli.
Conclusions:
The emergence of ESBL-producing pathogenic E. coli strains from diseased captive giant pandas warrants greater attention. The findings of this study will help to prevent the spread of these strains among captive giant pandas as well as from wild animals to humans.
Introduction
The giant panda (Ailuropoda melanoleuca) is an endangered and rare animal unique to China and is featured as the logo of the World Wildlife Fund to symbolize all endangered species. Beginning in the 16th century, the natural mountain habitat of the giant panda has become encroached by human settlements and is now limited to the provinces of Sichuan, Shaanxi, and Gansu. 1
In addition, the low reproductive rate and various infectious diseases have also negatively impacted survival. At the end of 2019, there were 600 artificially bred giant pandas (The 2019 Annual Conference of the Chinese Committee of Giant Panda Breeding Techniques held in Chengdu) and about 170 living in zoos throughout China. The giant panda prefers a vegetarian diet with bamboo as the principal staple food, 2 whereas captive giant pandas are also fed apples, carrots, and milk as well as other nutritional supplements.
Infiltration of the intestinal tract by pathogenic bacteria poses a significant threat to the health of the giant panda. 3 Among these pathogenic bacteria, diarrheagenic Escherichia coli is a zoonotic pathogen that can cause diarrhea and even death to both humans and giant pandas.4,5 Escherichia coli is a Gram-negative commensal bacterium commonly found in the digestive tracts of humans and various animals.
However, infection with pathogenic E. coli is a serious public health concern. Pathogenic E. coli are mainly divided into two groups depending on the disease location: (1) extraintestinal pathogenic E. coli, which include neonatal meningitis E. coli, uropathogenic E. coli, sepsis-associated E. coli, and avian pathogenic E. coli, 6 and (2) intestinal pathogenic E. coli, including enteropathogenic E. coli, enterohemorrhagic (Shiga toxin-producing) E. coli, enteroaggregative E. coli, enterotoxigenic E. coli, and enteroinvasive E. coli, which are causative agents of diarrheal disease, but differ regarding preferred host colonization sites, virulence mechanisms, and ensuing clinical symptoms.
Pathogenic E. coli strains isolated from various animal species have been extensively investigated. 4 However, little is known about the pathogenicity and characteristics of pathogenic E. coli in giant pandas. In 1992, enterohemorrhagic E. coli was isolated from captive giant pandas in China, but was undetectable after recovery. 7
Although carefully raised, giant pandas in zoo settings frequently acquire infections from common areas shared with tourists, birds, and mice. Antimicrobial treatment has resulted in the emergence of E. coli strains resistant to beta-lactams, tetracyclines, sulfonamides, and other antimicrobials.8–10 In 2015, a multidrug-resistant E. coli strain was isolated from a 3-year-old male giant panda in the Shanghai Wild Animal Park. 11 Although a common occurrence, intestinal diseases threaten the health of captive giant pandas and can result in death.
However, relatively few studies have investigated the prevalence of extended-spectrum beta-lactamase (ESBL)-producing E. coli and pathogenic E. coli in the intestine of the giant panda. Therefore, the aim of this study was to clarify the phenotypic, genetic, and pathogenic characteristics of E. coli in the intestine of captive giant pandas. In addition, the incidence of antibiotic resistance of four E. coli isolates from fecal samples collected in recent years from diseased giant pandas was investigated.
Materials and Methods
Bacterial strains
Four ESBL-positive E. coli strains were isolated from the fecal samples of four different diseased captive giant pandas from Sichuan Province (2008), Shaanxi Province (2012), and the Shanghai Wild Animal Park (2015), China. Two strains (AMSHJX03 and AMSHJX04) were isolated from the fecal samples of pandas housed in the same facility in Shaanxi Province. The strain AMSHJX01 (in this study) and the strain SH-YH-DH (accession no. KX129949.1) were isolated from different tissues of the same panda in the Shanghai Wild Animal Park.
Each of the pandas became anorexic and lassitude, with some developing fever and/or diarrhea (Table 1). The samples were resuspended in physiological saline and plated on MacConkey agar (Qingdao Hope Bio-Technology Co., Ltd., Qingdao, China) and incubated overnight at 37°C. One suspected E. coli colony was selected from each plate and recultured on McConkey agar for subsequent analysis.
Characteristics of Escherichia coli Strains from Diseased Captive Giant Pandas
Antimicrobial susceptibility in vitro
Antibiotic susceptibility analysis was conducted using the BD Phoenix-100 Automated Microbiology System (BD Biosciences, San Jose, CA) with drug sensitivity test plates containing 21 different antibiotics (i.e., amikacin [AMK], gentamicin [GEN], imipenem [IPM], meropenem [MEM], cefazolin [CFZ], ceftazidime [CAZ], cefotaxime [CTX], cefepime [FEP], aztreonam [ATM], ampicillin [AMP], piperacillin [PIP], amoxicillin-clavulanic acid [AMC], ampicillin-sulbactam [SAM], piperacillin-tazobactam [TZP], colistin [CST], trimethoprim-sulfamethoxazole [SXT], chloramphenicol [CHL], ciprofloxacin [CIP], levofloxacin [LVX], moxifloxacin [MXF], and tetracycline [TET]).
The ESBL phenotype was also recognized by the system. The minimum inhibitory concentration (MIC) was determined using the broth dilution method (Clinical and Laboratory Standards Institute 2018). The antibiotic resistance patterns of different strains were clarified with the use of distinct antibiotics, which included CFZ, CAZ, CTX, FEP, ATM, GEN, CHL, CST, and TET.
Antimicrobial resistance genes detection
Twenty-six pairs of primers were synthesized to detect target genes that conferred antibiotics resistance to CTX (blaCTX-M, blaCTX-M Group 1, blaCTX-M Group 2, blaCTX-M Group 9, blaSHV, and blaTEM),12–14 TET (tetA, tetB, tetC, tetD, tetW, and tetM),15–17 CST (mcr-1), 18 CHL (cat1, cmlA, and floR,), aminoglycosides (aph(3′)-IIa, aac(3)-IIa, aac(6′)-Ib, and ant(3″)-Ia) 8 and quinolone (qnrA, qnrB, qnrC, qnrVC, qnrD, and qnrS).19,20
Plasmid replicon typing
The E. coli strains and conjugants were examined for the presence of 18 plasmid replicons using a polymerase chain reaction (PCR)-based replicon typing method. All primers, annealing temperatures, and expected amplicon sizes are reported elsewhere. 21 The target genes were amplified with the use of a multiplex panel and confirmed by PCR. The positive amplicon sequences were compared with those in the National Center for Biotechnology Information (NCBI) database using the Basic Local Alignment Search Tool (BLAST). The plasmid replicon type of the strain and multilocus sequence typing (MLST) of the plasmid were identified using the PlasmidFinder and pMLST databases. The cutoff threshold for identity was set at 95% to determine the existence of a particular plasmid.
Conjugation
Conjugation experiments were conducted with ESBL phenotype strains by the filter method using an azide-resistant mutant of E. coli strain J53 as the recipient. Transconjugants were selected on tryptone soya agar containing CTX (4 mg/L) and sodium azide (200 mg/L). The antimicrobial susceptibility and ESBL phenotype of selected transconjugants were further characterized using the BD PhoenixTM-100 System. The presence of resistance genes and the replicon types of the plasmids from the transconjugants were determined using PCR, as described earlier.
Phylogenetic grouping and MLST
All strains were assigned to one of the eight main phylogenetic groups of E. coli (A, B1, B2, D, C, E, F, and clade I) by quadruplex PCR targeting of chuA, yjaA, arpA, and the DNA fragment TSPE4.C2. 22 MLST was also performed in accordance with Achtmans's scheme. Seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) were amplified by PCR and sequenced. Then, the sequences were compared with other known sequences using BLAST.
Whole-genome sequencing
Genomic DNA of the original strains was extracted with the sodium dodecyl sulfate method. The harvested DNA was detected by agarose gel electrophoresis and quantified using a Qubit fluorometer (Invitrogen Corporation, Carlsbad, CA). The genome of each strain was sequenced with Single Molecule Real-Time (SMRT) technology6,23 by Beijing Novogene Bioinformatics Technology Co., Ltd. (Beijing, China). The low-quality reads were filtered using SMRT Link v5.0.1 and the filtered reads were assembled to generate a single gap-free contig. At last the chromosome and plasmid sequences were screened by BLAST with plasmid database.
Pathogenicity and drug resistance gene function analysis
The secretory proteins were predicted using the SignalP-5.0 Server and type I–VII proteins secreted by the pathogenic bacteria were predicted with EffectiveT3 software. Predictions of virulence genes, antimicrobial resistance genes, and mobile elements were based on the VFDB (Virulence Factors of Pathogenic Bacteria) database, BLASTX, and CARD (Comprehensive Antibiotic Research Database). The O and H serotypes of the strains were predicted using the SerotypeFinder tool.
Comparative genomics analysis
To generate a phylogenetic tree depicting the positions of the aEPEC strain AMSHJX01 and strains AMSHJX02, AMSHJX03, and AMSHJX04 in this study, the genomes were compared with 28 published E. coli genomes using the neighbor-joining method. An evolutionary tree of the locus of enterocyte effacement (LEE) pathogenicity island in the genome of the aEPEC strain AMSHJX01 was constructed using the same method and the sequence of the whole island was compared with those of 24 published E. coli LEE islands and an E. albertii LEE island. ALL evolutionary analyses were conducted using Molecular Evolutionary Genetics Analysis 7 software.
Nucleotide sequence accession numbers
The whole-genome sequence data of strains AMSHJX01, AMSHJX02, AMSHJX03, and AMSHJX04 reported in this study have been deposited in the GenBank database under accession numbers CP030939, CP031105, CP058355, and CP058308, respectively. The accession numbers of the corresponding plasmids are CP030940 for strain AMSHJX01 (pAMSH1), CP031106–CP031111 for strain AMSHJX02 (pAMSC1- pAMSC6), CP058356 and CP058357 for strain AMSHJX03 (pAMPD1 and pAMPD2), and CP058309–CP058311 for strain AMSHJX04 (pAMPD01–pAMPD03).
Results
In vitro antimicrobial susceptibility testing of E. coli strains
A total of four strains were multidrug resistant and ESBL-positive. All four were resistant to six beta-lactam antibiotics (CAZ, CTX, FEP, ATM, AMX, and PIP) with the highest MICs obtained for CAZ (>1,024 mg/L), CTX (512 mg/L), FEP (512 mg/L), ATM (128 mg/L), AMP (>1,024 mg/L), and PIP (>1,024 mg/L). Only strain AMSHJX01 was resistant to the high-level MIC of CAZ (1,024 mg/L) and strain AMSHJX04 was resistant to the high-level MIC of CL (32 mg/L). The results are presented in Table 2.
Antimicrobial Characteristics of Escherichia coli Strains and Transconjugants
AMP, ampicillin; ATM, aztreonam; CAZ, ceftazidime; CFZ, cefazolin; CHL, chloramphenicol; CIP, ciprofloxacin; CST, colistin; CTX, cefotaxime; ESBL, extended-spectrum beta-lactamase; FEP, cefepime; GEN, gentamicin; LVX, levofloxacin; MIC, minimum inhibitory concentration; MXF, moxifloxacin; PIP, piperacillin; SAM, ampicillin-sulbactam; SXT, trimethoprim-sulfamethoxazole; TET, tetracycline.
Escherichia coli transconjugants and plasmid typing
All plasmid types of the E. coli strains were detected using PlasmidFinder. Also, the plasmid replicon type of the strain and transconjugant were confirmed using PCR-based replicon typing. The plasmid types IncHI2, IncN, IncFII, and IncFIB carried the CTX-M genes, which differed from each other. The four transconjugants were all ESBL-positive. Six β-lactam-resistant phenotypes (CFZ, CTX, FEP, ATM, AMP, and PIP) were transferred with plasmids. Strains AMSHJX01-t and AMSHJX02-t were multidrug resistant. The MICs of the transconjugants to most of the antibiotics were half of those of the strains, with the exception of CFZ (2 mg/L) and ATM (<0.25 mg/L) in the transconjugant AMSHJX01-t. The results are presented in Table 2.
E. coli strains resistome analysis
Identification of resistance genes was conducted using the whole-genome sequences. The antibiotic resistance genotypes of the strains were mostly due to efflux pump genes. Both ESBL (CTX-M type) and AmpC (CMY-62 type) were detected in two strains (AMSHJX01 and AMSHJX02). More antibiotic categories were detected in plasmids. The CTX-M genes of the plasmids from different strains were identified as blaCTX-M-55 and blaCTX-M-105.
One GEN-resistant strain (MIC = 256 mg/L) carried four aminoglycoside resistance genes in a large plasmid (257 kb), which included aph (6)-Id, aph (3″)-Ib, aac(6′)-Ib-cr, and aac (3)-IIa. One strain with a CST MIC of 32 mg/L possessed the mcr-1 gene. Two strains contained the intl1 gene, which encodes a class 1 integron. The antibiotic resistance genes carried by the plasmids are presented in Table 2. The distribution of blaCTX-M genes and related mobile elements are shown in Fig. 1.

Genetic context of mobile elements found in association with blaCTX-M genes. CTX-M genes are indicated with red arrows. Insertion sequences and transposons are indicated with blue and violet arrows, respectively. CTX, cefotaxime.
Virulence gene analysis
The virulence genes of four E. coli strains were analyzed using the VFDB database. Strain AMSHJX01 was identified as an aEPEC that possessed the entire 35-kb LEE pathogenicity island sequence, but lacked the BFP plasmid and stx gene, while carrying other important virulence factors, including Nle protein A and Paa (required for attaching/effacing activity), which are located outside of the LEE region.
Although the other three strains did not carry specific virulence genes, the genomes did encode some virulence-related genes. For example, strain AMSHJX02 carried the tsh gene encoding hemoglobin protease, whereas the genome of the lethal avian pathogenic E. coli carried the kps genes encoding the K1 capsule, which confers invasiveness. Meanwhile, strains AMSHJX03 and AMSHJX04 carried the eaeH gene, which encodes a protein that facilitates bacterial adhesin and invasion. In addition, strain AMSHJX04 carried the virulence gene cfa, which encodes CFA/I fimbriae. These three E. coli strains exhibited at least three important virulence markers of extraintestinal pathogenic E. coli and were, therefore, classified as members of this group.
Phylogenetic analysis of whole LEE pathogenicity island
The LEE of strain AMSHJX01, which is about 34,175 bp in length (nt 780253–814427), encodes a type III secretion system, intimin protein and translocated receptor, as well as translocons, chaperones, regulators, and secreted effector proteins that have been linked to virulence. The LEE sequence of strain AMSHJX01 was compared with that of 25 other strains retrieved from the NCBI database, which showed that the LEE sequence of strain AMSHJX01 was located within the same evolutionary branch (Fig. 2A) as those of strains CAU16175 (GenBank accession no. CP047378.1) and SMN013SH2 (GenBank accession no. CP023673.1), indicating a possible common ancestor, whereas the LEE islands of other strains may have evolved from another ancestor.
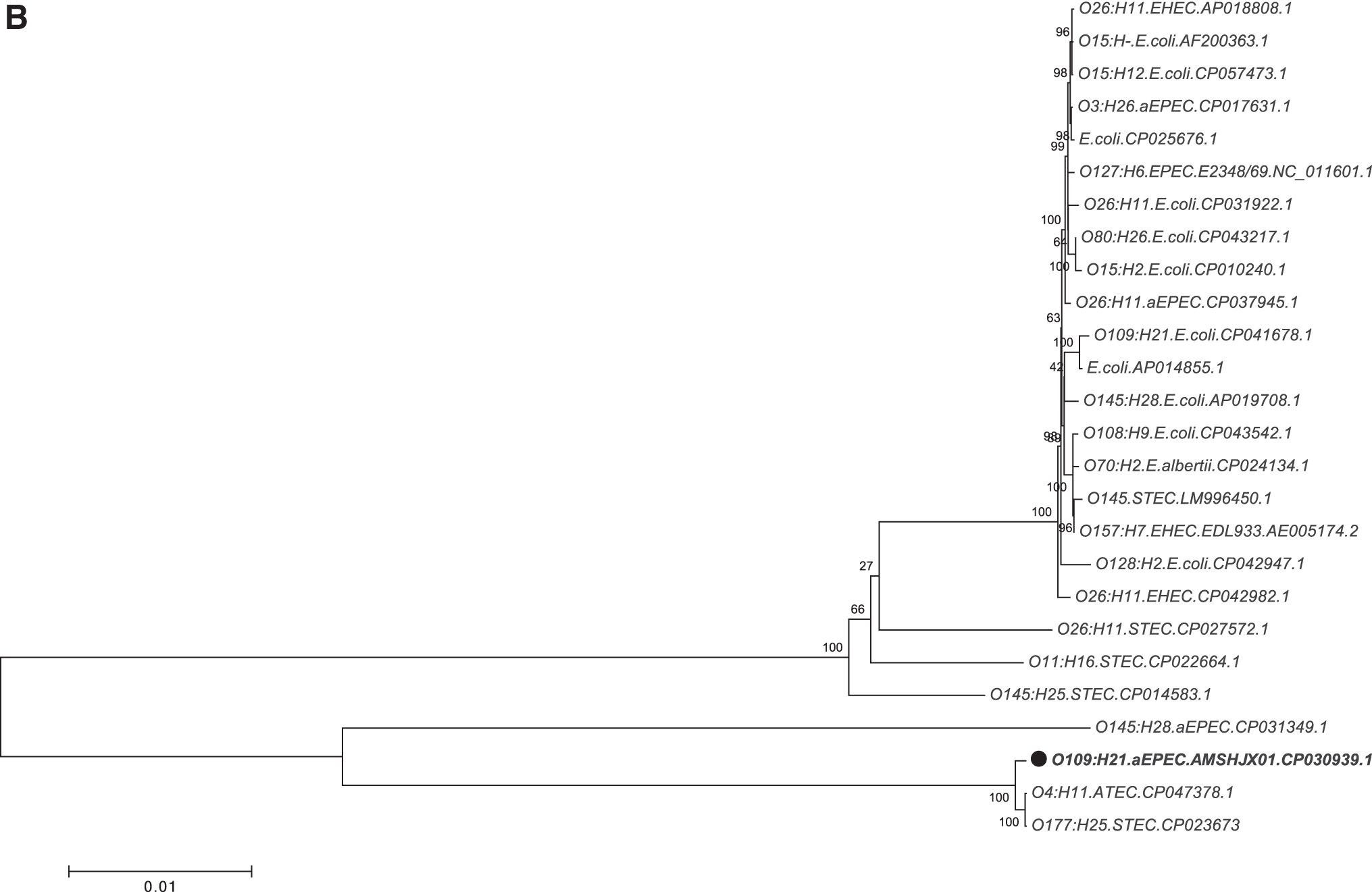
Phylogenetic analysis of completely sequenced Escherichia coli strains.
Phylogenetic analysis of whole genome
The evolutionary distances of the four strains were compared with 28 publicly available E. coli genomes of diverse pathotypes. The analysis showed that the strains were located on different evolutionary branches (Fig. 2B), but were closely related to strains isolated from domestic animals. Moreover, different Shiga-toxigenic E. coli strains were located on the evolutionary branches of strains AMSHJX01, AMSHJX02, and AMSHJX04. Nucleotide sequence accession numbers of the E. coli strains in this study were presented in Supplementary Table S1.
Discussion
The giant panda is a unique endangered species in China and chosen as the logo of the World Wildlife Fund. By January 2021, the wild population of giant pandas in China had increased to 1864. Infectious disease has become an emerging factor that may pose significant risk to endangered species. Owing to the relatively small population and geographical limitations, few studies have investigated infectious bacterial diseases of the giant panda and most previous reports were published in Chinese, which limited global dissemination. More recently, there have been an increase in English language reports of antibiotic-resistant bacteria, especially E. coli, isolated from giant pandas.1,9,10
Metagenomic analyses have found that E. coli accounts for 40.8% of the intestinal bacteria of the giant panda. However, the abundance and diversity of bacteria in the gut of wild giant pandas significantly differ from those of captive and semiwild giant pandas. 24 Notably, E. coli can acquire resistance and virulence genes from other strains or species through horizontal genetic exchange, which promotes antibiotic resistance of pathogenic strains. 25 ESBLs are a group of enzymes with the ability to hydrolyze oxyimino-cephalosporin and monobactams.
The spread of beta-lactamases is frequently linked with plasmid-mediated ESBLs, specifically the CTX-M family. 26 The data obtained in this study indicated that three CTX-M-type ESBL E. coli strains carried blaCTX-M-55 in various plasmids (pAMSH1-257kb, pAMPD2-89kb, and pAMPD02-129kb) belonging to diverse sequence types (i.e., ST40, ST716, and ST155). The replicon-type plasmids encoding blaCTX-M-55 included IncHI2, IncFII, and IncFIB. Distinct mobile elements, including IS26, ISEcp1, ISKpn72, IS903B, and Tn2, were identified up-/downstream of blaCTX-M-55 and blaCTX-M-105 (pAMSC4-47kb, IncN).
These elements have also been identified in other E.coli strains isolated from captive giant pandas. 27 The complicated genetic environment of the strains indicates that the spread of CTX-M genes was associated with various mobile elements as previously reported.28,29 The transconjugant ESBL-positive strain AMSHJX01-t carried both blaCTX-M-55 and blaTEM-1, but was more susceptible to CFZ (MIC, 2 mg/L) and ATM (MIC, 0.25 mg/L) than the donor strain (MIC, >1,024 and 128 mg/L, respectively) and two other CTX-M-55 transconjugants (MIC, 1,024 and >32 mg/L, respectively).
Meanwhile, comparisons with the use of nucleotide BLAST revealed that the sequences around blaCTX-M-55 of the three plasmids were similar. The difference in activity of the same beta-lactamase suggests that some transcriptional regulatory elements carried by the plasmid pAMSH1 contributed to this phenotype, although further investigations are needed.
These ESBL-positive E. coli strains also carrying virulence genes associated with invasion (e.g., eae, eaeH, and kps) or virulence-associated secretory systems (e.g., espL1, L4, R1, X1, X4, and X5) were predicted as human pathogens by the PathogenFinder web server. The aEPEC strain AMSHJX01 carried a LEE pathogenicity island. For more than a decade, aEPEC has been described as an emerging pathogen.
The LEE pathogenicity island is the only known virulence determinant of aEPEC. The genes located in the LEE pathogenicity island have accumulated extensive genetic diversity in the evolution process. 30 In this study, the LEE sequence of strain AMSHJX01 was compared with 25 others reported in the NCBI database. The results highlight the existence of globally disseminated aEPEC strains that have acquired different LEE lineages in their evolutionary histories, which will be critical for future studies of the emergence of new aEPEC strains.
The presence of resistance genes and virulence-associated genes critically reduce the efficacy of clinical treatment, resulting in poorer recovery of captive giant pandas from infections. Meanwhile, these strains act as reservoirs of resistance genes that could spread to animal keepers, veterinarians, and zoo visitors, among others. Hence, further understanding of the structural and genetic backgrounds of these ESBL-producing pathogenic E. coli will help to protect endangered and rare animals, such as the giant panda.
To strengthen protections of the giant panda, artificial breeding programs are ongoing in China. Notably, some captive giant pandas have been released into natural habitats. However, because of close contact with humans, captive giant pandas are prone to infections by various human pathogens. The reintroduction of giant pandas into natural environments also increases the risk of infection with unknown pathogens. Hence, ongoing etiological surveillance of captive giant pandas is critical.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.