
Abstract
Background:
Recently, the emergence of carbapenem-resistant hypermucoviscous Klebsiella pneumoniae has aroused increasing attention in China. We investigated the characteristics of a Klebsiella pneumoniae carbapenemase-2 (KPC-2) and New Delhi metallo-β-lactamase-5 (NDM-5) coproducing hypermucoviscous K. pneumoniae strain, named RJ-8061, which was isolated from the urine of an 86-year-old female patient with pneumonia.
Methods:
The RJ-8061 strain was investigated by string test, antimicrobial susceptibility testing, polymerase chain reaction for carbapenemase genes detection, capsular genotyping, multilocus sequence typing, whole-genome sequencing, and phylogenetics. A serum killing assay and a Galleria mellonella infection model were used to evaluate the virulence of RJ-8061 in vitro and in vivo.
Results:
RJ-8061 belonged to the sequence type 11 K64 serotype and showed high-level resistance to almost all frequently used antibiotics, only remaining susceptible to amikacin, colistin, and tigecycline. The complete genome size of RJ-8061 was 6,106,028 bp, including a 5,394,921 bp chromosome and seven circular plasmids. Plasmid pRJ-8061-hybrid is a 294,249 bp hybrid plasmid that co-harbored resistance genes [blaTEM-1B, mph(A), aac(3)-IId] and virulence genes (iucABCDiutA, rmpA2), whereas rmpA2 is a truncated version. In addition, blaKPC-2 and blaNDM-5 were located on plasmids 171,321 bp pRJ-8061-KPC-2 (IncFII/IncR) and 46,161 bp pRJ-8061-NDM-5 (IncX3), respectively. K-mer-based phylogenetic analysis grouped RJ-8061 into a carbapenem-resistant Klebsiella pneumoniae cluster. The G. mellonella infection model revealed that RJ-8061 showed relatively low virulence, with a 50% lethal dose of 106 cfu.
Conclusions:
To the best of our knowledge, this is the first report of a hypermucoviscous K. pneumoniae coproducing KPC-2 and NDM-5 carbapenemases.
Introduction
K
Until now, 126 KPC variants (KPC-1 to KPC-126) have been identified according to www.bldb.eu/BLDB.php?prot=A#KPC. In China, KPC-2 is the main genetic determinant of CRKP, 5 and these CRKP mainly belong to sequence type 11 (ST11). 5 Since NDM-1 was first detected in New Delhi in 2009, 6 42 NDM variants (NDM-1 to NDM-42) have been identified. Compared with NDM-1, NDM-5 has been shown to exhibit increased resistance to carbapenems. 7 Recently, the number of coproducing carbapenemase K. pneumoniae, which leads to higher level resistance to carbapenems, has been increasing.8–11
Compared with CRKP, hvKP generally displays a higher susceptibility to most antimicrobials. 12 However, hvKP strains often cause severe acute infection, and can even lead to metastatic spread in relatively healthy individuals with normal immunity due to their hypervirulence. 13 Virulence genes in hvKP include regulator of mucoid phenotype A (rmpA/rmpA2), aerobactin (iucABCD and iutA), salmochelin (iroBCDN), enterobactin (entABCDEFS, fepABCDG, and fes), and yersiniabactin (ybtAEPQSTUX, irp1/2, and fyuA). The hvKP strains predominantly belong to ST23, ST65, and ST86 clones, and most of them belong to K1 and K2 serotypes. 14
With the transmission of mobile genetic elements (MGEs), carbapenem-resistant hypervirulent (hypermucoviscous) Klebsiella pneumoniae (CR-hvKP/HMKP) that converges carbapenem resistance and hypervirulence, has emerged in recent years.15–19 Currently, CR-hvKP/HMKP strains have been progressively identified and reported in China, although they remain uncommon. 20 Furthermore, CR-hvKP/HMKP coproducing carbapenemases is even more rare.9,21,22 In this study, we report a KPC-2 and NDM-5 carbapenemases coproducing hypermucoviscous ST11 K. pneumoniae strain.
Materials and Methods
Source of the strain
Initially, we aimed to retrospectively investigate the molecular characteristics of K. pneumoniae isolated from Renji Hospital. One hundred twenty-one CRKP isolates were collected between January 1, 2019 and August 31, 2020. Among them, the RJ-8061 strain, isolated from the urine of an 86-year-old female patient with pneumonia and coproduced KPC and NDM carbapenemases, attracted our attention.
Antimicrobial susceptibility tests
Antimicrobial susceptibility testing was performed using the broth microdilution method for ceftazidime, cefepime, aztreonam, gentamicin, amikacin, levofloxacin, piperacillin/tazobactam, cefoperazone/sulbactam, imipenem, meropenem, ceftazidime/avibactam, colistin, and tigecycline. ATCC25922 was used as quality control. The minimum inhibitory concentration (MIC) results were interpreted according to the Clinical Laboratory Standards Institute (CLSI 2020) guidelines. The breakpoint of tigecycline was based on the standards of the European Committee for Antimicrobial Susceptibility Testing (EUCAST 2019).
Polymerase chain reaction screening of carbapenemase and virulence genes
RJ-8061 was screened for carbapenemase genes (blaKPC, blaNDM, blaOXA-48, blaIMP, and blaVIM) and several virulence genes (iucA, iutA, iroB, iroN, rmpA, and rmpA2) by polymerase chain reaction (PCR) and sequencing. The primers are listed in Supplementary Table S1.
Multilocus sequence typing
Multilocus sequence typing (MLST) was performed with seven housekeeping genes according to the protocol described on the K. pneumoniae MLST website. 23
DNA preparation, genome sequencing, and annotation
The total genomic DNA of RJ-8061 was extracted using TIANamp Bacteria DNA Kit (TIANGEN BIOTECH, BEIJING CO., LTD) and was used for whole-genome sequencing (WGS). The genome was sequenced using a combination of Illumina Hiseq X Ten (2 × 150 bp) sequencing and PacBio RS II Single MoleculeReal Time (SMRT) platforms. A statistic of quality information was applied for quality trimming, which was used to remove the low-quality data to form clean data. Final error correction of the PacBio assembly results was performed using the Illumina reads by Pilon (version 1.22). Glimmer (version 3.02) was used for coding sequence (CDS) prediction. The predicted CDSs were annotated from Non-Redundant Protein Database, Swiss-Prot, Pfam, Gene Ontology, Clusters of Orthologous Groups of Proteins, and Kyoto Encyclopedia of Genes and Genomes databases using sequence alignment tool Basic Local Alignment Search Tool (BLAST+) (version 2.3.0), Diamond (version 0.8.35), and HMMER (version 3.2.1).
Clean data were assembled in CLC Genomics Workbench (version 21.0.3; QIAGEN, Aarhus, Denmark); then the generated de novo assembled contigs were carried on BLAST using Virulence Factors Database (VFDB, 2021-05) and the comprehensive antibiotic resistance database (CARD) to verify the presence of virulence and resistant genes. The assembled plasmid contigs were submitted to PlasmidFinder 2.1 to investigate the incompatibility replicon type. The MGEs in the genomic sequences were identified by insertion sequences (IS) finder for insertion sequences. Capsular type was determined using the assembled contigs to query the wzi allele databases.
Comparative genomic and phylogenetic analyses
Sequence comparison was performed by BLASTn and a circular map was generated using the BLAST Ring Image Generator (version 0.95). The program Easyfig (2.2.3) was applied to comparative schematic diagrams. The phylogenetic K-mer tree was generated by aligning the whole genome of RJ-8061 with that of 9 CRKP and 9 hvKP mentioned in the literature published in the past 5 years (Supplementary Table S2 in Supplementary Data) using CLC Genomics Workbench version 21 (CLC; QIAGEN) as described previously. 24
String test, serum killing assay, and Galleria mellonella infection model
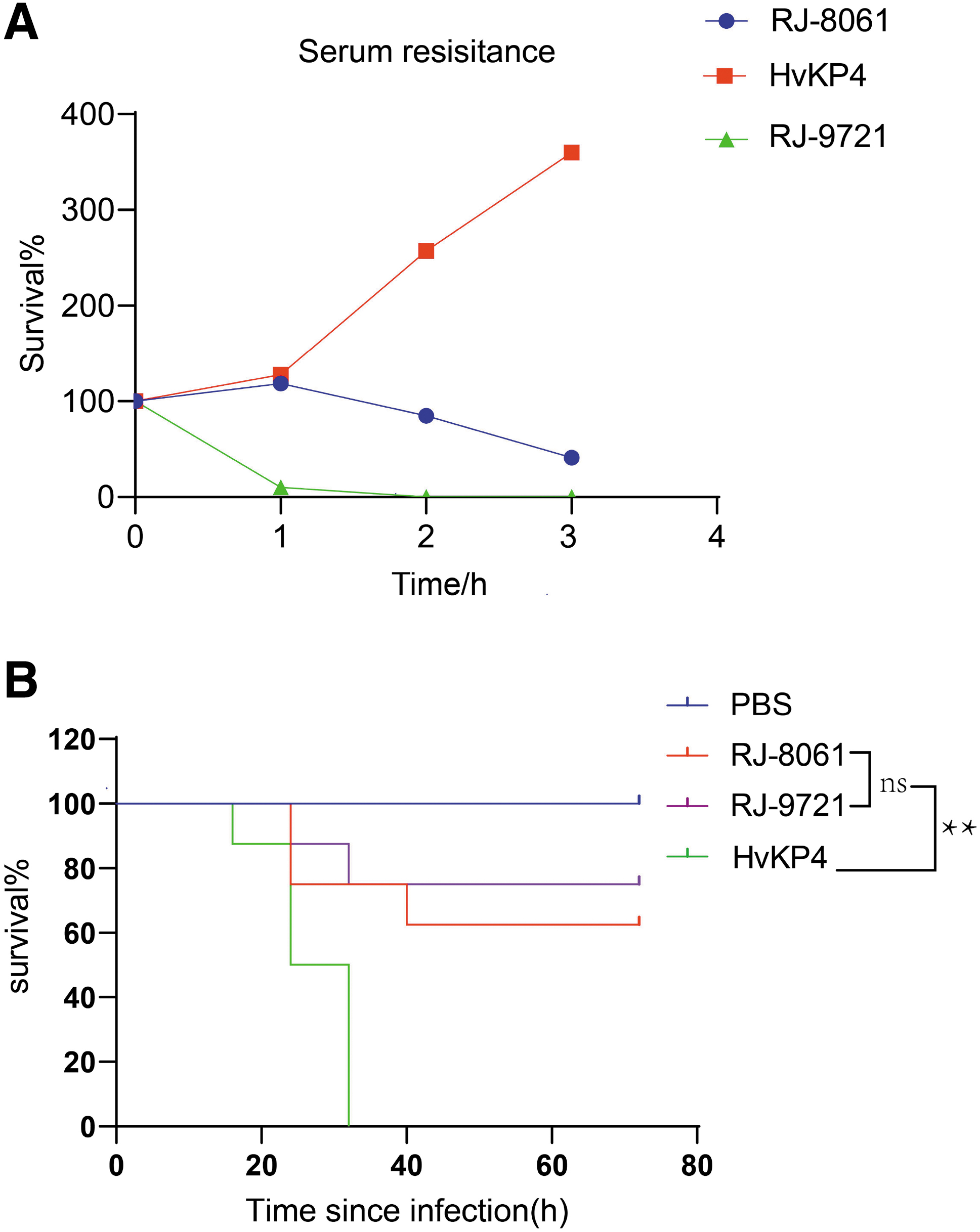
The hypermucoviscous phenotype was identified using string test, 13 which was deemed positive when a viscous string >5 mm could be generated by touching and pulling a single colony upward with a standard inoculation loop on an agar plate. The hypermucoviscous hvKP strain CG43 and an ST11 classic Klebsiella pneumoniae (cKP) RJ-7994 were used as positive and negative controls, respectively. Serum killing was used to estimate in vitro the virulence of RJ-8061 as previously described. 25 In brief, 106 cfu/mL bacterial suspension and pooled human serum (1:3) were mixed and incubated at 37°C. Viable counts were obtained by plating serial dilutions on Mueller–Hinton agar at 0, 1, 2, and 3 hours. The survival rate (SV) was represented as SV1h/0h, SV2h/0h, and SV3h/0h. The response to serum killing in terms of SV was scored using six grades. 26
An ST11 cKP (RJ-9721 50% lethal dose [LD50]: 1 × 107 cfu) and an ST23 hvKP (hvKP4 LD50: 1 × 104 cfu) were used as the low-virulence control and hypervirulence control, respectively. Subsequently, a Galleria mellonella pathogen-free (purchased from Tianjin Huiyude Biotech Company, Tianjin, China) infection model was used to evaluate the virulence of RJ-8061 as described previously. 27 Eight G. mellonella in each group were injected with 10 μL of a bacterial suspension (with concentrations of 105, 106, and 107 cfu/mL) and placed in petri dishes at 31°C. The viability of the G. mellonella was recorded every 8 hours until 72 hours. The exact inoculation dose was confirmed on Luria-Bertani agar, and the LD50 was calculated using the SigmaPlot program (version 11.0). The survival curve was assessed by Kaplan–Meier analysis and log-rank test. p-Values <0.05 were considered statistically significant.
Nucleotide sequence accession number
The complete genome sequence of Klebsiella pneumoniae RJ-8061 was deposited in the National Center for Biotechnology Information (NCBI) GenBank database under the accession number PRJNA793126.
Approval statement
This study was approved by the Ethics Committee of Renji Hospital, School of Medicine, Shanghai Jiaotong University, Shanghai, China. All individual patients or their legal guardians provided informed consent.
Results
Antimicrobial susceptibility testing and molecular characteristics
Klebsiella pneumoniae RJ-8061 exhibited high-level resistance to ceftazidime (MIC, ≥128 μg/mL), cefotaxime (MIC, ≥128 μg/mL), cefepime (MIC, ≥128 μg/mL), aztreonam (MIC, ≥128 μg/mL), gentamicin (MIC, 64 μg/mL), levofloxacin (MIC, 16 μg/mL), piperacillin/tazobactam (MIC, ≥128 μg/mL), cefoperazone/sulbactam (MIC, ≥128 μg/mL), imipenem (MIC, ≥128 μg/mL), and meropenem (MIC, ≥256 μg/mL), whereas only remaining susceptible to amikacin (MIC, 4 μg/mL), colistin (MIC, 0.75 μg/mL), and tigecycline (MIC, 0.5 μg/mL). In addition, RJ-8061 displayed high-level resistance to ceftazidime/avibactam (MIC, ≥128 μg/mL), suggesting metallo-β-lactamase (MBL) production. The typical hypermucoviscous phenotype of RJ-8061 was determined by the positive string test. PCR screening and MLST revealed that RJ-8061 co-harbored KPC-2 and NDM-5 carbapenemases and belonged to ST11 K64 serotype.
Genome and phylogenetic analyses
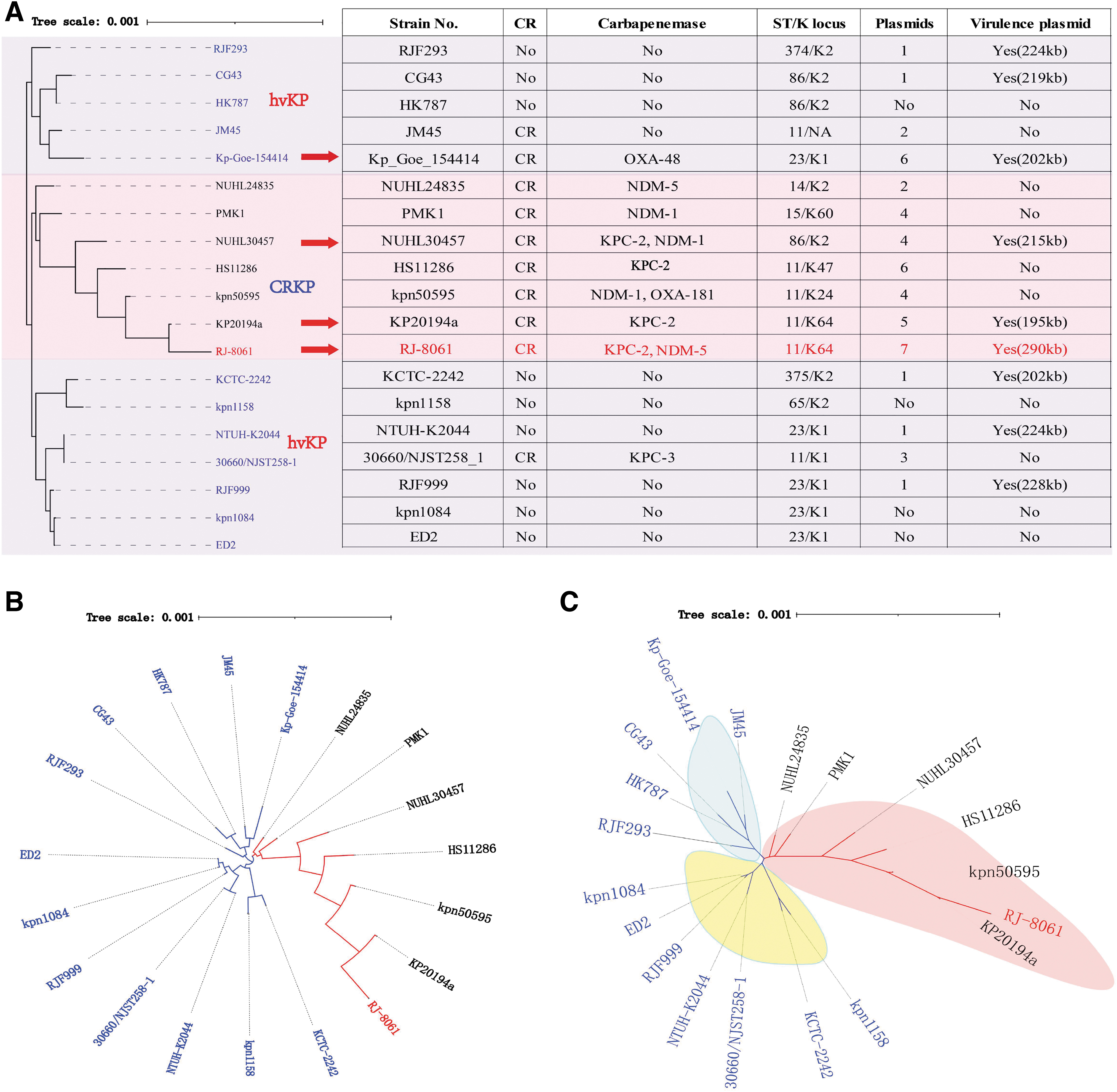
The sequencing depth of RJ-8061 was 231 × coverage of the genome using Illumina Hiseq X Ten (2 × 150 bp) sequencing and 229 × coverage of the genome using PacBio RS II SMRT platforms. WGS showed a total of 6,106,028 bp for RJ-8061, including a chromosome and seven circular plasmids, named pRJ-8061-hybrid, pRJ-8061-KPC-2, pRJ-8061-NDM-5, pRJ-8061-A, pRJ-8061-B, pRJ-8061-C, and pRJ-8061-D. RJ-8061 chromosome carried several resistance genes, including blaSHV-187, aadA2, sul1, qacE, fosA6, ant(3″)-Ia and aadA13, and numerous virulence genes, including fim(ABCDEFGHIK), mrk(ABCDFHIJ), ent(A∼F), fep(ABCDG), iroE, tle1, and tli1 (Table 1).
Genomic Information of the Klebsiella pneumoniae RJ-8061
Genomic analysis revealed that the plasmid pRJ-8061-hybrid (294,249 bp) belonging to IncHI1B/IncFIB type was a hybrid plasmid that simultaneously harbored resistance and virulence genes with an average ratio of guanine to cytosine (GC) content of 47.48%. VFDB and CARD analysis showed that the plasmid pRJ-8061-hybrid not only contained a range of virulence genes (iucABCD, iutA, and rmpA2) but also β-lactamase gene (blaTEM-1B), macrolide resistance gene (mphA), and aminoglycoside resistance gene [aac(3)-IId]. As shown in Fig. 1A, some IS were scattered across the complete sequence of the plasmid pRJ-8061-hybrid, which indicated that the aforementioned genes might insert into the plasmid pRJ-8061-hybrid through ISs.

Comparative analysis showed that the pRJ-8061-hybrid shared 87% to 89% coverage and 99.5% to 99.67% identity with pKpvST147B (339 kb, CP040726.1) from KpvST147B isolated in east England; pEBS1036 (347 kb, MT648512.1) from EBS1036 isolated in Guangdong, China; and pKpvST383L (372 kb, CP034201.2) from KpvST383 isolated in London. In addition, a ∼16-kb region in the pRJ-8061-hybrid was highly similar to the known virulence plasmid pLVPK (AY378100.1) from Klebsiella pneumoniae CG43, with 99.34% identity (Fig. 1A), and the covered region containing the virulence genes iucABCD and iutA (Fig. 1B).
The pRJ-8061-KPC-2 plasmid belonged to an IncFII/IncR incompatibility type and exhibited an average GC content of 53.83%. In addition, a BLAST comparison showed that pRJ-8061-KPC-2 had 91%–93% coverage and 99.9% nucleotide identity with pBS1066 (MT269843) from BS1066 isolated in Guangdong, China; pKPC2_020037 (CP036372) from WCHKP020037 isolated in Chengdu, China; pJX2-2 (CP064248) from JX-CR-hvKP-2 isolated in Nanchang, China; and plasmid unnamed2 (CP023942.1) from FDAARGOS-444 isolated in Canada (Fig. 1C). Analysis of the genetic environment around blaKPC-2 showed that it was identical to pJX2-2 and pBS1066 (Fig. 1D).
The downstream region of blaKPC-2 was a segment (-KPC-ISKpn6-IS26-Tral-); its upstream region contained a set of transposon elements (-TniA-IS26-ΔTn3-ISKpn27-KPC-), seven mer operons, which were separated from transposon elements mentioned earlier by one open reading frame (ORF), and two truncated transposons, IS5075 (family IS110 [1,327 bp]) and IS903B (family IS5 [1,057 bp]), which were located upstream region of those mer operons (Fig. 2B). Surprisingly, when compared with plasmid unnamed2, the surrounding sequence of blaKPC-2 in pRJ-8061-KPC-2 was identical to an inverted sequence in plasmid unnamed2. Otherwise, compared with pKPC2_020037, their upstream region was identical, but the downstream region was distinct. We found an inserted fragment (-KPC-ISKpn27-ΔTn3-IS26-ISKpn6-) between ISKpn6 and blaKPC-2 in the downstream region of blaKPC-2 in pKPC2_020037 (Fig. 1D).
The blaNDM-5 gene was located on an IncX3-type plasmid of 46,161 bp, designated pRJ-8061-NDM-5, which shared 100% coverage and 100% identity with plasmid pP768-NDM-5 (MF547510) from Escherichia coli strain p768MEM-011 isolated in China (Fig. 1E).
The whole-genome K-mer-based phylogenetic analysis indicated that RJ-8061 was clustered into CRKP clade (Fig. 2A). The unrooted tree also clearly displayed that RJ-8061 was closely related to CRKP strains, such as KP20194a (Hunan, China), HS11286 (Shanghai, China), NUHL30457 (Jiangxi, China), NUHL24835 (Jiangxi, China), PMK1 (the United Kingdom), and kpn50595 (the Czech Republic), but distant from the clades grouped by hvKP strains (Fig. 2B, C). Therefore, RJ-8061 might have evolved from a CRKP strain by acquiring a virulence plasmid.
In vitro and in vivo virulence of RJ-8061
A serum killing test showed that RJ-8061 was intermediately sensitive (SV1h/0h > 100% but SV2h/0h < 100%, SV3h/0h < 100%) and was designated as Grade 3 (Fig. 3A). The G. mellonella infection experiment indicated that the LD50 of RJ-8061 was 1 × 106 cfu. In addition, the Kaplan–Meier survival estimate for Klebsiella pneumoniae RJ-8061-infected G. mellonella revealed that RJ-8061 showed similar virulence to the low-virulence control RJ-9721 at 72 hours postinfection (Fig. 3B). The aforementioned results indicate that the virulence of strain RJ-8061 was relatively low. In contrast to the hypervirulent control strain hvKP4, virulence genes rmpA and yersiniabactin genes (ybtX, ybtQ, ybtP, ybtU, ybtS, ybtP, ybtA, ybtT, ybtE, irp2, irp1, and fyuA) were not present in the RJ-8061 strain. This might be one of the reasons for the difference in their virulence.
Discussion
CR-hvKP, which converged carbapenem resistance and hypervirulence, has posed a great challenge to public health. 28 In line with this, carbapenemase coproduction in a CR-hvKP strain makes the situation worse. In this study, we report a KPC-2 and NDM-5 coproducing ST11 K. pneumoniae, which also displayed a hypermucoviscous phenotype. The coexistence of blaKPC-2 and blaNDM-5 resistance genes was consistent with the multidrug resistance (MDR) phenotype of RJ-8061. Particularly, RJ-8061 was resistant to ceftazidime/avibactam, suggesting the production of MBL.29,30 Further analysis confirmed that RJ-8061 carried blaNDM-5 gene. In addition, we identified blaSHV-187 in the chromosome of RJ-8061.
Unfortunately, virulence plasmids carrying antimicrobial resistance genes is becoming increasingly common. Hybrid plasmids containing both virulence and resistance genes have recently been reported in various STs of K. pneumoniae strains, including the hypervirulence clone ST23 and MDR ST11 clone. 14 The pRJ-8061-hybrid plasmid might be generated from a virulence plasmid that acquired resistance genes or a resistant plasmid that acquired virulence genes by using recombination or IS-mediated transfer.
As for virulence, RJ-8061 contained some hvKP-specific virulence-associated genes, including aerobactin (iucABCD, iutA), the regulator of mucoid phenotype A2 (rmpA2), type I fimbriae (fim ABCDEFGHIK), and type 3 fimbriae (mrkABCDFHIJ). Although numerous virulence genes were carried by RJ-8061, the strain was intermediately resistant in the serum killing test (Grade 3). In this study, we speculated that the relatively low virulence of RJ-8061 is partially attributed to the nonfunction of the rmpA2 gene and the missing of virulence genes, such as yersiniabactin (ybtX, ybtQ, ybtP, ybtU, ybtS, ybtP, ybtA, ybtT, ybtE, irp2, irp1, and fyuA) and the regulator of mucoid phenotype (rmpA). Generally, an intact rmpA2 gene is 639 bp in length. Nevertheless, we found a truncated rmpA2 gene in plasmid pRJ-8061-hybrid that was 618 bp in length, which might be nonfunctional. The discovery of such rmpA2 variants has appeared in more than 40 other K. pneumoniae strains and has been recorded in the NCBI database.31,32
Together, RJ-8061 possesses an extraordinary number of virulence genes and a high diversity of resistance factors. Our research underlines the importance of raising public awareness of the presence of resistance and virulence genes in the same plasmid or strain. Therefore, it is of great significance to strictly supervise and manage the carbapenem-resistant hypermucoviscous Klebsiella pneumoniae (CR-HMKP) strains coproducing carbapenemases. Further studies are needed to evaluate whether silent transmission has occurred in China. Overall, further analysis of carbapenemase-encoding and virulence plasmids may provide insight into the ongoing recombination and evolution of virulence and MDR in K. pneumoniae.
Conclusions
In this study, we identified an ST11 K64 serotype blaKPC-2 and blaNDM-5 coproducing CR-HMKP strain, namely, RJ-8061. RJ-8061 contained seven plasmids, including a hybrid plasmid pRJ-8061-hybrid, KPC-2 harboring plasmid pRJ-8061-KPC-2, NDM-5 harboring plasmid pRJ-8061-NDM-5, and four other plasmids. Coproduction of KPC-2 and NDM-5 carbapenemase in CR-hvKP is very uncommon. Despite the string test being positive, the strain did not show high virulence. Thus, there is an urgent need for detailed molecular studies of the superbugs, which could provide comprehensive insights into the evolution process of these strains.
Footnotes
Authors' Contributions
J.Z. designed this project; X.J. and L.Z. performed experiments; X.J., J.Z., and Z.S. analyzed the data and prepared figures; X.J. and J.Z. drafted the article. All authors contributed to data interpretation and read and commented on the article.
Disclosure Statement
All authors report no potential conflict of interest.
Funding Information
This study was supported by the National Natural Science Foundation of China (81702062) and the Scientific Research Project of Shanghai Municipal Health Commission (20204Y0288).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.