
Abstract
Macrolides are commonly used to control respiratory tract infections in ruminants, but the susceptibility of Pasteurella multocida strains has shown a decrease to macrolide antibiotics in the last decade. In this work we assessed the prevalence of macrolide resistance of 100 P. multocida isolates from ruminant hosts and studied the resistance genotypes with newly designed PCRs. Susceptibility to erythromycin and tilmicosin was tested using minimal inhibitory concentration (MIC) test strips. A newly designed PCR was used for the detection of macAB genes, and a PCR plus restriction enzyme-based technique was developed for detecting a 23S rRNA gene mutation at position 2059. Five bovine isolates with notably increased MICs (≥256 μg/mL for erythromycin and ≥32 μg/mL for tilmicosin) carried resistance genes msr(E) and mph(E) or the A2059G point mutation in the 23S rRNA gene. Over 73% strains from small ruminants and all bovine isolates were MacAB PCR positive. Bovine strains were less sensitive to macrolide antibiotics than isolates from small ruminants, and an increase in the prevalence of macrolide resistance in bovine P. multocida isolates has also been observed over time.
Introduction
P
Antibiotic resistance in pathogenic bacteria is one of the most important challenges to public and veterinary health. 4 The high degree of resistance necessitates a constant search for new antimicrobial agents for the prevention and control of infections caused by resistant clones. Macrolides are commonly used to control respiratory tract infections in ruminants. 5 However, the susceptibility of P. multocida strains has shown a decrease to macrolide antibiotics, including erythromycin, tilmicosin, and tulathromycin, in the last decade.6–8 Resistance may occur by acquired resistance genes, which include rRNA methylases (erm(42)), macrolide transporters (msr(E)), and macrolide phosphotransferases (mph(E)).9–11 P. multocida can also acquire resistance to macrolides by a 23S rRNA mutation at nucleotide A2059G, resulting in highly resistant isolates (minimal inhibitory concentrations [MICs] >64 mg/L). 12
In addition to these, various efflux pumps have been reported to contribute to macrolide resistance in P. multocida and other respiratory pathogens. 13 Of these, the first antibiotic-specific drug transporter described in Gram-negative bacteria is MacAB, a macrolide-specific ATP-binding casette (ABC)-type efflux carrier. 14 It was demonstrated that plasmids carrying macA and B genes conferred resistance to 14- and 15-membered macrolides, and it was also described that erythromycin is a good substrate for the MacAB efflux pump. 15 The aim of our work was to investigate the prevalence of macrolide resistance in a collection of 100 P. multocida isolates from ruminant hosts and to study the macrolide resistance genotypes with newly designed PCRs.
Materials and Methods
P. multocida strains used in this study
A total of 100 isolates of P. multocida recovered from cattle (n = 58) and small ruminants (sheep, n = 30 and goat, n = 12) in Hungary from 2005 to 2021 were used in this study (Supplementary Table S1). Isolates were recovered from 71 herds representing 69 locations all over the country, with no known connection between any of the herds. All samples were collected from clinical cases, either from live animals (nasal swab and milk sample) or during post-mortem examinations (lung, spleen, and liver). The isolates were cultured on Columbia agar (LAB M Ltd., Bury, United Kingdom) plates supplemented with 5% sheep blood under aerobic conditions at 37°C for 24 hr. The identity of the isolates was confirmed by a species-specific PCR, 16 and reference strain X73 was used as a positive control.
Determination of MIC
To study macrolide resistance, the 14-membered erythromycin and the second-generation 16-membered tilmicosin, widely used in clinical practice, were chosen. Susceptibility to erythromycin and tilmicosin was tested using MIC test strips (Liofilchem, Roseto, Italy). The strains were cultured on Mueller–Hinton agar plates supplemented with 5% sheep blood at 37°C for 24 hr. Bacterial suspensions in phosphate-buffered saline, adjusted to a density of 0.5 McFarland, were spread onto Mueller–Hinton agar (LAB M Ltd.) plates supplemented with 5% sheep blood using a sterile swab. An MIC test strip was placed on each plate after ∼10 min and the plates were incubated at 37°C for 24 hr, following which the MIC values were read according to the manufacturer's instructions.
For quality control, Staphylococcus aureus ATCC 25923 and Escherichia coli ATCC 25922 were used. The MICs of the quality control strains were within the acceptable Clinical and Laboratory Standards Institute (CLSI) range. We interpreted the breakpoints according to CLSI recommendations.17–19 The following susceptible, intermediate and resistant breakpoints were used: erythromycin S: ≤0.5, I: 1–4, R: ≥8 and tilmicosin S: ≤16, R: ≥32. The results were also analyzed before and after 2011–2012, which is approximately the middle of the study period, to assess trends in changes in antibiotic resistance.
PCRs and primer design
Acquired macrolide resistance genes (erm(42), msr(E), and mph(E)) were detected by a previously described multiplex PCR. 10 The sequences of the oligonucleotide primers designed in this study are listed in Table 1. Primers specific for macA and macB genes coding a macrolide-specific ABC-type efflux protein were designed from the complete genome sequence of NCTC10204 reference strain (GenBank accession no. LR134298).
Sequences of Oligonucleotides Designed in This Study
These sequences match the coding strand of the macA (primer F) and macB (primer R) gene of Pasteurella multocida NCTC10204 strain with GenBank NZ_ LR134298.
These sequences match the coding strand of the 23S rRNA gene of P. multocida Pm70 strain with GenBank NR_103956.
The binding site of the forward primer is located in the macA gene, while the binding site of the reverse primer is located in the macB gene, because these two genes are arranged in tandem in a single operon, which allows the two genes to be detected in one reaction. 20 Primers specific for the 23S rRNA gene of P. multocida were designed from the sequences of the P. multocida Pm70 strain (GenBank accession no. NR_103956). The primers were synthesized by Sigma-Aldrich (Haverhill, United Kingdom). Primers macAB F and macAB R were designed to produce a 1,009 bp PCR fragment from the macAB genes, whereas primers 23S F and 23S R amplified a 932 bp fragment from the 23S rRNA gene. Amplicon sizes were calculated from the DNA sequences of P. multocida reference strains, and our formerly characterized strain Pm238 was used as positive control. 21
DNA preparation
Lysates of pure cultures were prepared for all strains used in this study. A loopful of cultured bacterial growth was suspended in 100 μL of PCR grade water (VWR, Radnor, PA) and heated in a thermal cycler for 20 min at 99°C. Cellular debris was pelleted by centrifugation and the supernatant was used as DNA template for PCR amplification. The lysates were stored at −20°C until use.
23S rRNA PCR-restriction fragment length polymorphism
23S rRNA PCR was performed in a total volume of 25 μL containing 100 ng of template DNA, 2.5 μL of 10 × DreamTaq buffer, 2 μL 25 mM MgCl2, 0.5 μL of 10 mM dNTPs, 1 μL of forward and reverse primers (10 pmol/μL), and 1 U of DreamTaq polymerase (Thermo Scientific, Waltham). DNA was amplified for 25 cycles of denaturation at 95°C for 30 s, annealing at 64°C for 30 s, and extension at 72°C for 1 min. An initial denaturation step (95°C for 10 min) and a final extension step (72°C for 7 min) were also performed. Five microliters of PCR products was digested in a 30 μL volume containing 2 μL of 10 × FastDigest buffer and 1 μL of FastDigest Eco31I enzyme (Thermo Scientific). Reaction mixtures were incubated at 37°C for 15 min. Ten microliters of digested fragments from 23S rRNA gene was examined by electrophoresis.
MacAB PCR
MacAB PCR was performed in a total volume of 25 μL containing 100 ng of template DNA, 2.5 μL of 10 × DreamTaq buffer, 0.5 μL 25 mM MgCl2, 0.5 μL of 10 mM dNTPs, 1 μL of forward and reverse primers (10 pmol/μL), and 1 U of DreamTaq polymerase (Thermo Scientific). DNA was amplified for 25 cycles of denaturation at 95°C for 1 min, annealing at 54°C for 1 min, and extension at 72°C for 1 min. An initial denaturation step (95°C for 10 min) and a final extension step (72°C for 7 min) were also performed.
Analysis of amplification and digestion products
All reactions were performed in a Bio-Rad C1000 thermal cycler (Bio-Rad Laboratories, Inc., Berkeley, CA). To test the sensitivity of our assay, 10-fold dilutions of genomic DNA (106–100) of strain 4251 were used. DNA was quantified (NanoDrop spectrophotometer; Thermo Scientific) and template copy numbers per reaction were calculated using an online calculator.
Each amplification or digestion product was analyzed by electrophoresis in a 1.5% agarose gel (SeaKem, Lonza, Basel, Switzerland) stained with GelRed (Biotium, Inc., Hayward) and visualized under UV light. The results were confirmed by sequencing of PCR products. Sequencing was performed by Macrogen Europe (Amsterdam, The Netherlands). Nucleotide sequences were analyzed using Geneious Prime software (version 2020.2.4) and confirmed by BLAST (www.ncbi.nlm.nih.gov/blast/Blast.cgi). The GenBank accession numbers for sequences obtained in this study are OL441367–OL441370 and OM162018 (macA), OL441371–OL441374 and OM162019 (macB), and OL413486–OL413491 (23S rRNA).
Results
The identity of all P. multocida isolates was confirmed by the species-specific PCR. The erythromycin and tilmicosin MICs are shown in Fig. 1 and Tables 2 and 3. Only one of the bovine strains (and none of the strains from small ruminants) was sensitive to erythromycin (MIC 0.5 μg/mL) according to the available breakpoint. 17 MICs were 1 to 4 μg/mL (intermediate resistance) for 53.5% (n = 31) of bovine strains, and 76.2% (n = 32) of isolates from small ruminants.
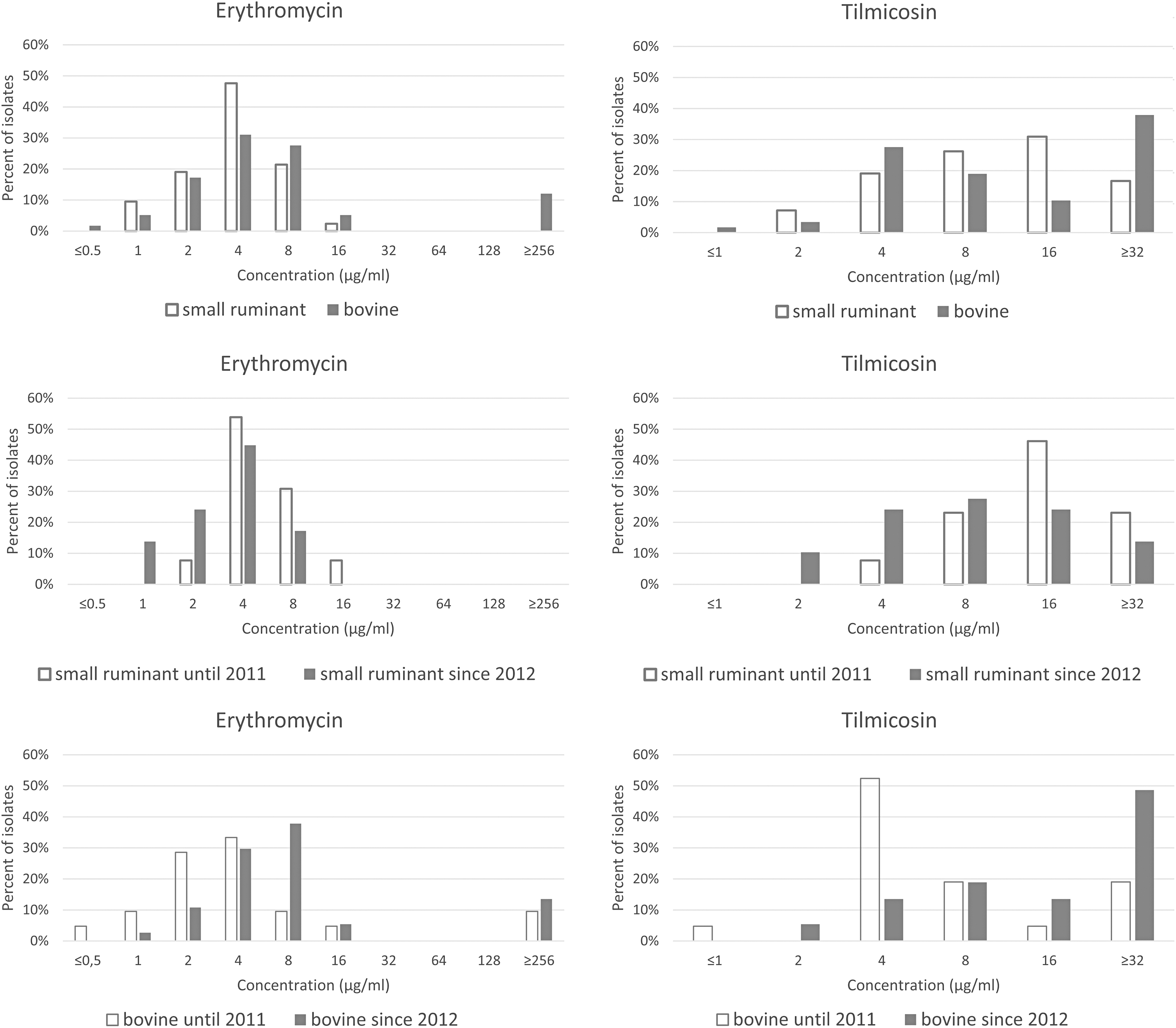
MIC distributions for the 100 Hungarian Pasteurella multocida isolates tested in this study comparing the type of antibiotic with host species and period of isolation. MIC, minimal inhibitory concentration.
MIC50 and MIC90 Values of the 100 Hungarian Pasteurella multocida Isolates Tested in This Study
MIC, minimal inhibitory concentration.
Distribution of Macrolide Resistance Determinants and Minimal Inhibitory Concentrations of Erythromycin and Tilmicosin for Pasteurella multocida Isolates Investigated in This Study
According to CLSI, 17 isolates with MICs ≤0.5 μg/mL were considered susceptible, isolates with MICs between 1 and 4 μg/mL were classified as intermediate, and isolates with MICs ≥8 μg/mL were considered resistant.
According to CLSI, 18 isolates with MICs ≤16 μg/mL were considered susceptible and isolates with an MIC of 32 μg/mL were considered resistant.
MICs were ≥8 μg/mL (considered resistant) for 44.8% (n = 26) of bovine isolates and 23.8% (n = 10) of strains from small ruminants. Seven out of the 58 isolates (12.1%) from cattle exhibited an MIC ≥256 μg/mL, suggesting a high level of resistance against this antibiotic. These strains were collected from six herds located in different regions of the country at considerable distances from each other, excluding any possible connection between them. The majority of strains from cattle and from small ruminants was inhibited by concentrations of ≤16 μg/mL of tilmicosin (62.1%, n = 30; and 83.3%, n = 22, respectively), and was considered susceptible. 18 The rest of the isolates were resistant with MICs ≥32 μg/mL.
Five out of seven bovine P. multocida strains with remarkably increased MICs (≥256 μg/mL for erythromycin and ≥32 μg/mL for tilmicosin) carried resistance genes msr(E) and mph(E) or the A2059G point mutation in the 23S rRNA gene, in addition to harboring macA and macB genes.
Among macrolide resistance genes, both msr(E) and mph(E) were detected together in two bovine strains (3.4%), while erm(42) could not be identified in any isolate. The PCR plus restriction enzyme-based technique developed for detecting a 23S rRNA gene mutation at position 2059 was able to identify three mutant strains (5.2%) of bovine P. multocida isolates (Fig. 2). Strains from small ruminants were negative in the above reactions. MacAB PCR was positive in all bovine isolates, and in 73.8% (n = 31) of isolates from small ruminants (Fig. 2). The sensitivities of assays developed in this study were 105 genomic equivalents per reaction.

Discussion
In this study, 100 isolates of P. multocida of bovine and small ruminant origin were characterized by determining their macrolide resistance genotypes and MICs to erythromycin and tilmicosin. A PCR for the detection of macA and macB genes, and a PCR plus restriction enzyme-based technique for detecting a 23S rRNA gene mutation at position 2059 were also developed, which were suitable for the investigation of macrolide resistance genotypes.
Macrolide resistance determinants erm(42) and/or msr(E)-mph(E) were first reported in a small number in P. multocida and Mannheimia haemolytica isolates from bovine respiratory disease (BRD) cases.6,9 A recent study found a correlation between the presence of erm(42) and/or msr(E)-mph(E) genes and elevated gamithromycin and tildipirosin MIC values in P. multocida and M. haemolytica field isolates from BRD cases. 8 In another study, a multiplex PCR to detect erm(42), msr(E), and mph(E) genes in P. multocida and M. haemolytica isolates also showed that the presence of these genes correlated with higher MIC values to veterinary macrolides (tilmicosin, tulathromycin, gamithromycin, and tildipirosin). 10
Furthermore, this study confirmed that strains with msr(E) or mph(E) always possessed both genes as a tandem pair, as previously observed.6,8 Therefore, the presence of msr(E)-mph(E) in our strains was not surprising. On the other hand, only two of our P. multocida isolates carried resistance genes msr(E)-mph(E) while erm(42) was not detected in any of the resistant strains. A recent study detected no antimicrobial resistance genes in P. multocida isolates with phenotypic resistance to erythromycin, tilmicosin, and tulathromycin, suggesting the existence of point mutations in the bacterial ribosome or other resistance mechanisms. 13
Macrolide resistance may develop by 23S rRNA mutations in P. multocida strains where acquired resistance determinants are absent. 12 In these usually highly resistant P. multocida isolates, an adenosine to guanosine mutation at nucleotide 2059 has been found. 12 This mechanism was also reported in P. multocida isolates that lacked integrative and conjugative elements, as well as erm(42), msr(E), and mph(E) genes. 22 This A2059G mutation was detectable in a small percentage of strains (n = 3) tested in this study, and these isolates were highly resistant to erythromycin and tilmicosin, showing remarkably high MIC values.
We could only explain the genetic background of macrolide resistance by the presence of resistance genes or 23S rRNA gene mutation in a small percentage of our isolates. A previous study also hypothesized that the explanation for this phenomenon may be an increased expression of an intrinsic gene, probably an efflux pump.7,13 To clarify this issue, we developed a PCR detecting macA and macB genes, and to the best of our knowledge, our study is the first in which the investigation and detection of macAB genes in a large number of P. multocida isolates were performed. MacAB PCR was positive in all bovine strains; thus, we were able to detect genes underlying the genetic background of macrolide resistance in each isolate from this host species.
MacAB PCR was also positive in the majority of isolates from small ruminants, further confirming that the macAB efflux pump may be a major contributing factor to macrolide resistance in P. multocida. At the same time, these strains were negative in all other reactions performed in the study, while macAB genes were also undetectable in 23.2% (n = 11) of these isolates. This observation suggests the existence of other efflux proteins or mutations in the L4 and L22 ribosomal protein genes involved in the development of macrolide resistance.6,13 It was surprising that the only bovine isolate that was sensitive to erythromycin (MIC = 0.5 μg/mL) carried the macA and macB genes.
However, sequence analysis of macAB genes of this strain revealed an amino acid substitution (A43T) in the macB gene, which might affect the structure and function of the cytoplasmic ATP binding domain.23,24 We also found that five (71.4%) out of the seven isolates with high erythromycin and tilmicosin MIC values carried msr(E) and mph(E) genes or a 23S rRNA gene mutation, in addition to macAB genes.
These erythromycin and tilmicosin MIC data suggest a somehow increasing tendency in the incidence of macrolide resistance in bovine P. multocida isolates over time (Fig. 1 and Table 2). A similar trend has been reported in Australia to tilmicosin, tulathromycin, and gamithromycin, and the resistance phenotype correlated with the carriage of msr(E) and mph(E) genes in all, but one macrolide-resistant isolate. 11
This time-dependent increase in resistance was not detected in isolates from small ruminants in Hungary, and based on MIC50 and MIC90 data, we found that bovine strains were less sensitive to macrolides than the isolates from small ruminants (Table 2). This may indicate that there is a lower selection pressure on P. multocida strains infecting small ruminants, presumably due to the lower use of antibiotics compared to that used in the cattle farming industry. This confirms and complements the previous finding that ovine isolates are resistant to fewer antibiotic agents and have lower MIC values than strains from pigs. 25
Conclusions
The newly designed molecular methods reported in this study enabled us to conduct a comprehensive analysis of macrolide resistance in isolates of P. multocida. More generally, these techniques may contribute to the rapid and accurate detection of resistance genes, which may help better understanding the mechanisms of bacterial resistance in P. multocida. The time-dependent increase in macrolide resistance in P. multocida underlines the importance of prudent and responsible use of antibiotics to reduce the selection pressure that promotes the development and spread of highly resistant clones.
Footnotes
Acknowledgments
The outstanding technical assistance of Éva Hegedűs and Katalin Oryszcsák is highly appreciated. The authors thank Dr. A.J. Lax for the useful comments on the article.
Authors' Contributions
B.U. designed the research, performed the PCRs and data analysis, wrote the article, contributed to discussion, and reviewed and edited the article. T.M. wrote the article, contributed to discussion, and reviewed and edited the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
Our study was supported by the National Research, Development and Innovation Office (grant NKFIH K124457) and the SA-27/2021 grant of the Eötvös Loránd Research Network.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.