
Abstract
Background:
Extensive use of carbapenems to treat multidrug-resistant (MDR) Gram-negative bacteria (GNB) facilitates the wide dissemination of carbapenemase-producing carbapenem-resistant GNB. Colistin was reintroduced into clinical settings to manage these GNB infections. However, there is currently an increase in the dissemination of mobile colistin resistance (mcr)-producing colistin-resistant GNB isolates in clinical settings. The epidemiology of carbapenemases and mcr in Pretoria was evaluated.
Methods:
Clinical MDR GNB were collected and screened for carbapenemases and mcr using polymerase chain reaction (PCR); their antibiotic susceptibility profiles were elucidated using the Vitek® 2 automated system (Biomerieux, France) and microbroth dilution (for colistin).
Results and Discussion:
A total of 306 isolates were collected; a majority of these were Klebsiella pneumoniae (n = 208) and were collected from males (n = 158). The isolates were retrieved from a variety of infection sites, including urine, blood cultures, and rectal swabs. The Vitek 2 system found that these isolates were largely resistant to β-lactams, where 217 (70.9%) had reduced susceptibility to at least one carbapenem (ertapenem, meropenem, or imipenem), and 81 isolates (26.5%) were resistant to colistin. PCR screening identified 201 (65.7%) isolates harboring carbapenemase genes consisting of blaOXA-48 (170, 84.2%), blaNDM (31, 15.4%), blaIMP (5, 2%), blaKPC (4, 1%), and blaVIM (5, 2%). Furthermore, 14 blaOXA-48–producing isolates were coharboring blaVIM (2), blaNDM (9), blaKPC (1), and blaIMP (2) genes. Only one isolate harbored the mobile colistin resistance (mcr)-1 gene, and this is the first report of an mcr-1–producing Acinetobacter baumannii isolate in South Africa.
Conclusion:
There is high endemicity of carbapenemase genes and a low prevalence of mcr genes in GNB, particularly in K. pneumoniae, in health care facilities in Pretoria and surrounding regions of South Africa.
Significance:
Health care facilities in Pretoria are becoming breeding grounds for MDR infections that threaten public health. Careful use of carbapenems and other antibiotics is necessary to prevent further escalation and outbreak of these MDR strains that can claim several lives.
Introduction
Antibiotic resistance is a global public health threat because it decreases therapeutic options for infectious diseases while stalling the progress of modern medicine.1–4 Resistance to all classes of antibiotics has been observed, including “last-resort” ones such as carbapenems and colistin (polymyxin E).1,5 Carbapenems are β-lactam antibiotics with broad bactericidal activity against both Gram-positive and Gram-negative pathogens in aerobic and anaerobic environments.6,7
Carbapenems were initially used in the clinical setting to treat fatal infections caused by Enterobacterales that produce extended spectrum β-lactamases (ESBLs).6,8 However, the extensive use of carbapenems resulted in the emergence of carbapenem-resistant Gram-negative bacteria (GNB).6,9 Carbapenem resistance may develop through a variety of cellular mechanisms, which include carbapenemase production, increased activity of efflux pumps, and by porin mutations. These mechanisms may either alone or accompanied by overexpression of AmpC or an EsβL, cause carbapenem resistance.9–11
Carbapenemases are a specific group of β-lactamases that hydrolyze carbapenems and render them inactive.6,8,9,12 Carbapenemase genes have been discovered worldwide, with carbapenem-resistant GNB infections constituting a 40% mortality rate.13,14 A study in South Africa found that 70% of carbapenem-resistant Enterobacteriaceae (CRE) infections were hospital acquired, and the in-hospital mortality rate was 38%. 15 These genes are usually associated with mobile genetic elements such as plasmids, allowing for the wide dissemination of these genes across GNB families.
Colistin was designated as the “highest critically important antimicrobial for human medicine” by the World Health Organization.16,17 It is part of the polymyxin antibiotic class of cationic cyclic polypeptide antibiotics that attack the lipopolysaccharide (LPS) layer found on the outer membrane of GNB.13,18 Colistin was reintroduced into the clinical setting to treat critically ill patients suspected of infections due to multidrug-resistant (MDR) bacteria.7,12,16,19 Acquired resistance to colistin is achieved through various mechanisms, one of which is the modification of the LPS, which reduces the bactericidal activity of colistin.13,20
LPS modifications can be achieved through various chromosomal mutations within genes that are within the two-component systems pmrA/pmrB and phoP/phoQ; that is, mgrB, crrb, phoQ, pmrB, etc.10,11 Resistance can also develop through the acquisition of mobile colistin resistance (mcr) genes that encode MCR enzymes that mediate LPS modifications.11,21 Mcr genes have been identified in multiple hospitals across South Africa, mostly in Escherichia coli and Klebsiella pneumoniae, and have also been identified in livestock and in effluents of wastewater treatment plants.22–24
Newton-Foot et al. screened for colistin-resistant E. coli and K. pneumoniae isolates collected between January and August 2016 by the National Health Laboratory Service (NHLS) at Tygerberg Hospital, and found that 83% (15 of 18) of them were MCR-1 producing. 24 This highlights an increase in mcr genes in the Tygerberg Academic hospital and surrounding regional and district hospitals. 24
Carbapenemase and mcr genes are commonly associated with mobile genetic elements such as plasmids, insertion sequences, and transposons.5,24–27 This association allows for the rapid dissemination of these resistance genes worldwide, thus posing a threat to public health, 26 because they endanger the efficacy of carbapenems and colistin during treatment.1,3
Resistance to the last-resort antibiotics, carbapenems and colistin, should be carefully monitored to prevent further spread of bacterial resistance and to inform treatment options. 13 Molecular tools such as polymerase chain reaction (PCR) and whole-genome sequencing have been shown to be useful in the identification of resistance genes 28 and for screening of known resistance genes. 6 This study describes the molecular epidemiology of carbapenemase and mcr genes in MDR GNB collected from the medical microbiology laboratory of the Tshwane academic division of the NHLS (Tshwane Academic Division) culture biobank.
Methods
A total of 306 MDR clinical Gram-negative isolates were collected from NHLS/UP. All the clinical isolates were subjected to identification and antibiotic susceptibility testing using the Vitek-2 automated system (BioMérieux, France) according to the manufacturer's instructions. Further, the epidemiological data such as sex, age, and sample source were retrieved from the NHLS TrakCare system (Supplementary Table S1).
Antimicrobial susceptibility test of Gram-negative isolates
Identification and antimicrobial susceptibility testing were performed on the isolates using the Vitek® 2 automated system (BioMérieux) against 17 antibiotics. These included the following: amikacin, gentamicin, ampicillin, amoxicillin-clavulanate, piperacillin tazobactam, cefuroxime, cefotaxime/ceftriaxone, ceftazidime, ertapenem, imipenem, meropenem, colistin, ciprofloxacin, tigecycline, trimethoprim sulfamethoxazole, cefoxitin, and cefepime.
Isolates identified to be colistin resistant were selected for broth microdilution (BMD) to determine their exact minimum inhibition concentration (MIC) values (Supplementary Table S1). The BMD was performed according to the CLSI standards 29 using E. coli ATCC® 25922™ and/or Pseudomonas aeruginosa ATCC 27853™ for quality control.
PCR-based screening of carbapenemase and mcr genes
All 306 isolates were screened for the presence of carbapenem and colistin resistance genes, regardless of their susceptibility profile. The PCR screening included two multiplex PCRs: one was made up of all five mcr primers seen in Table 1, and the second was made up of blaVIM, blaOXA, and blaNDM primers. The PCR further consisted of two singleplex PCRs for both blaKPC and blaIMP. The PCRs were performed according to Supplementary Table S2. Amplicons and a 100 bp DNA ladder (Promega) were viewed using 2% agarose gel electrophoresis.
Primer Sequences for mcr and Carbapenemase Polymerase Chain Reaction Screening
mcr, mobile colistin resistance; KPC, Klebsiella pneumoniae carbapenemase.
In brief, colonies of overnight culture were placed in Eppendorf tubes with 1 mL of 1 × phosphate buffer saline and boiled for 10 minutes in a water bath. The tubes were transferred to an ice bath for 5 minutes, and then were centrifuged for 5 minutes at 1,000 rpm (123 g). Three microliters of supernatant were used for PCR. The PCR premixes were made up of 12.5 μL GoTaq Green Master Mix (Promega), 3 μL of genomic DNA, 1.5 μL for each primer, and nuclease-free water to a final volume of 25 μL. The PCRs were conducted according to Supplementary Table S2.
Ethical approval
Ethics approval was received from the Faculty of Health Sciences: Research Ethics Committee at the University of Pretoria under the reference 581/2020.
Results
A total of 306 MDR clinical GNB isolates were collected from NHLS bacterial culture biobank. These isolates comprised of 208 K. pneumoniae, 29 E. coli, 20 Acinetobacter baumannii, 16 Enterobacter cloacae complex, 10 P. aeruginosa, 9 Salmonella Group D, 3 Klebsiella oxytoca, 2 Serratia marcescens, 2 Morganella morganii, and single isolates each of Citrobacter freundii, Citrobacter koseri, Shigella flexneri, Proteus mirabilis, Proteus vulgaris, and Providencia stuartii. The bacterial strains were isolated from a variety of infection sites, which includes the following: urine (n = 84), blood culture (n = 68), rectal swab (RecSw) (n = 31), aspirate (fluid/tracheal, treated; n = 24), tissue (n = 18), pus (aspirate/swab; n = 15), catheter tip (arterial/intravenous; n = 14), sputum (n = 13), catheter urine (9), central spinal fluid (9), swab (5), stool (4), bronchial alveolar lavage (1), and ventriculoperitoneal shunt (1).
Urine was the predominant sample collected, where the majority (96.4%) of the isolates isolated were of the Enterobacteriaceae family, which are common etiological agents of urine tract infections. Klebsiella pneumoniae was isolated from 21 different infection sites, with urine, blood cultures, and RecSws being the predominant sites (Fig. 1). Thereafter, A. baumannii and E. coli were isolated from nine and eight sites, respectively.
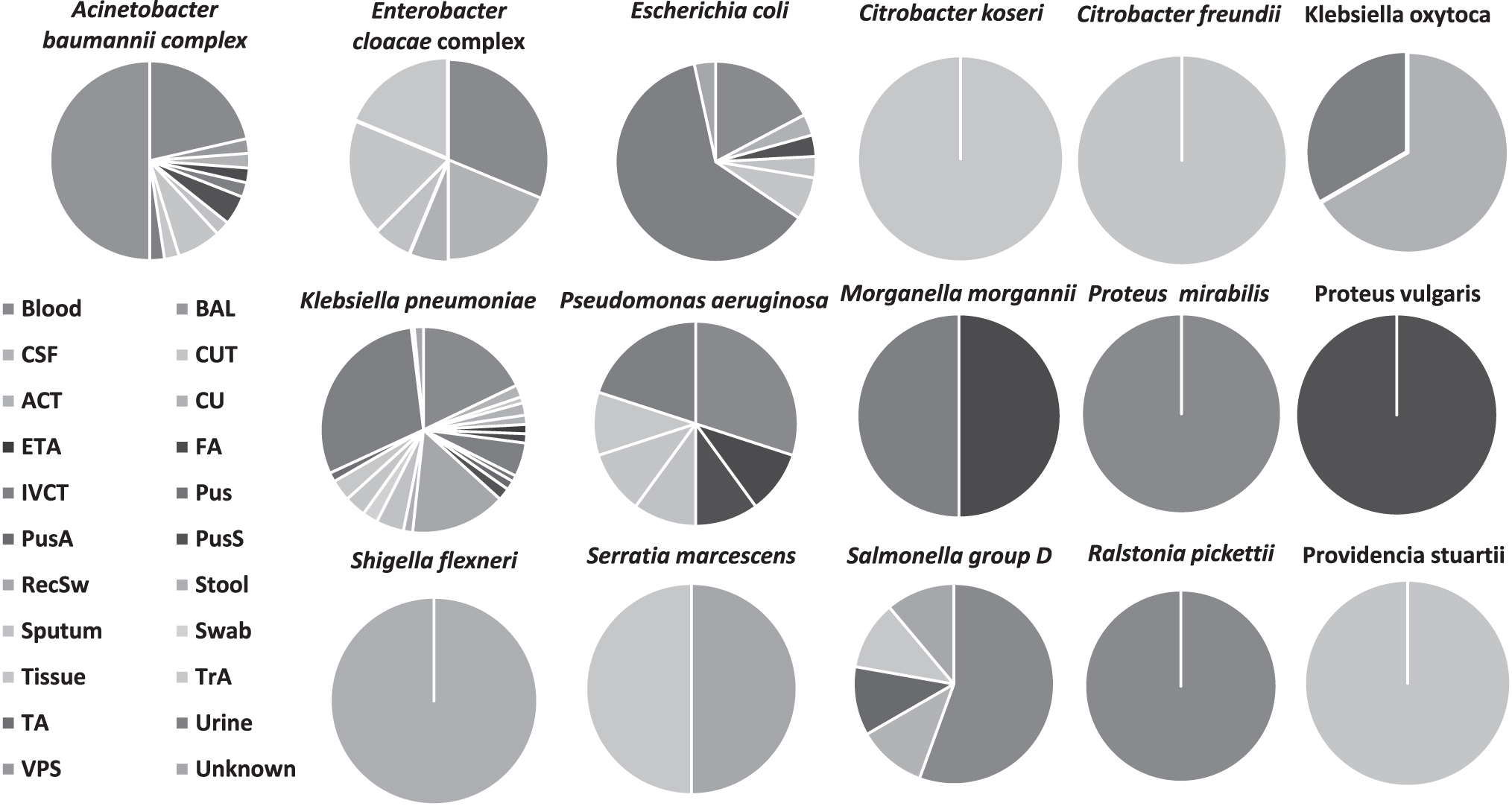
The distribution of sample sources for each MDR clinical Gram-negative isolate. The sample sources are made up of blood culture, CSF, ACT, ETA, IVCT, PusA, RecSw, sputum, tissue, TA, VPS, BAL, CUT, CU, FA, Pus, PusS, stool, swab, TA, and urine. ACT, arterial catheter tip; BAL, bronchial alveolar lavage; CSF, cerebrospinal fluid; CU, catheter urine; CUT, catheter urine tip; ETA, endotracheal aspirate; FA, fluid aspirate; IVCT, intravenous catheter tip; MDR, multidrug-resistant; PusA, Pus aspirate; PusS, Pus swab; TA, treated aspirate; VPS, ventriculoperitoneal shunt; RecSw, rectal swab.
The epidemiological data of each GNB isolate are observed in Supplementary Table S1 and summarized in Supplementary Table S3 (Fig. 2). These data show that majority of the isolates were collected from males (158, 51.63%) and middle-aged people (135, 44.11%), where 100 of the 135 isolates collected from this age group were K. pneumoniae (Fig. 2b). Neonates, infants <1 year, constituted 12.46% (38) of total isolates; 73.68% (28) of these neonatal isolates were obtained from infants <1 month. Neonates are highly susceptible to bacterial infections due to their immature immune system; thus, neonatal infections lead to a rapid progression of disease.30,31 It is, therefore, concerning to observe MDR GNB in this age group.
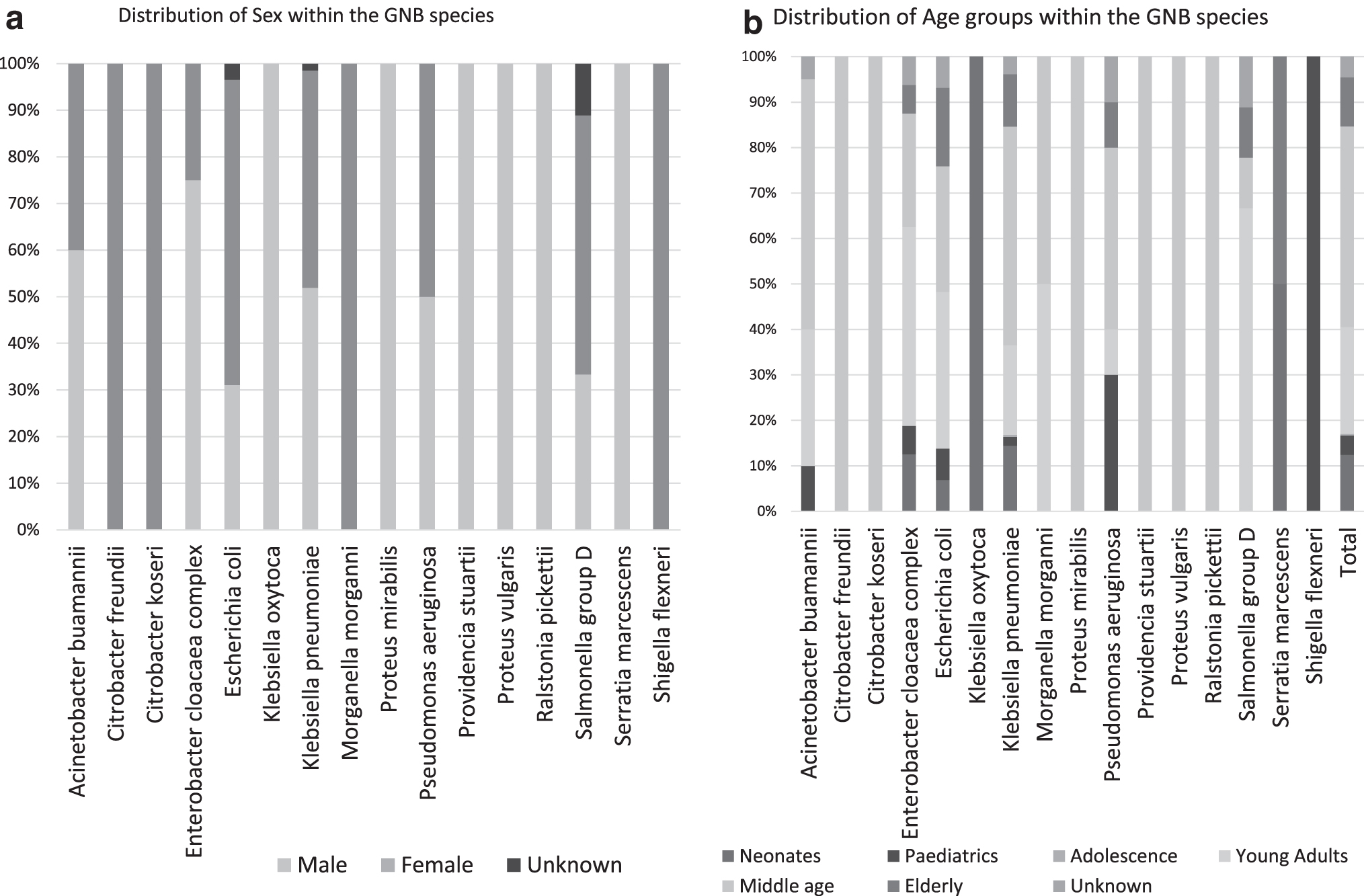
The demographics of
Antimicrobial susceptibility profile results
The antimicrobial susceptibility profile seen in Supplementary Table S1 reveals that more than two-thirds of the isolates were highly resistant to β-lactam antibiotics (ampicillin 77.8%, ceftriaxone/cefotaxime 74.5%, cefuroxime 71.2%, cefepime 69.3%, ceftazidime 69.3%, and cefoxitin 68.5%). Further, 217 (70.9%) of isolates were resistant to at least one carbapenem, and resistance to each included 57.5% (179) ertapenem, 44.8% (139) imipenem, and 41.5% (127) meropenem. The frequency of carbapenem-resistant isolates was calculated to be 64.7% with K. pneumoniae isolates being the most predominant. There were, however, low levels of colistin-resistant isolates (80, 26.1%) seen among the 306 isolates, further, among those found to be ertapenem resistant, only 10% (19), were colistin resistant.
Prevalence of carbapenemase genes
All the 306 MDR GNB were subjected to carbapenemase screening regardless of their antibiotic susceptibility profile. The five carbapenemase genes included in the singleplex and multiplex PCR were Klebsiella pneumoniae carbapenemase (KPC) (blaKPC), metallo-β-lactamases (MBLs) (blaIMP, blaVIM, blaNDM), and OXA-48 (blaOXA-48). Positive controls were included in PCRs to ensure PCR validity, and amplicons are seen in Fig. 3b.

Gel electrophoresis of PCR amplicons obtained from the isolates. Two percent TBE agarose gel of PCR controls used in the study, ran against a 100 bp ladder
Overall, a total of 201 (65.7%) of isolates were found to harbor one or more carbapenemase genes, and a total of 216 carbapenemase genes were identified. The results, shown in Fig. 4, included 171 (55.6%) OXA-48 producers, where 14 of these isolates also coharbored an additional carbapenemase gene (Fig. 4). MBL producers were 41 (13.4%), and were made up of nine NDM-OXA-48 coproducers, 20 NDM producers, 2 IMP-VIM-OXA coproducers, 2 VIM-OXA 48 producers, 1 IMP producer, 1 VIM producer, and 1 VIM NDM producer (Fig. 4). Finally, eight KPC producers were identified, where one isolate coharbored KPC with OXA-48. The carbapenemase genes were largely identified in K. pneumoniae isolates (92%).

Distribution of carbapenemases identified in the isolates. Two hundred one isolates were identified to be harboring carbapenemase genes; however, 16 isolates were harboring ≥2 carbapenemases. One hundred fifty-seven isolates were harboring blaOXA-48 genes only, thereafter two were coharbored with blaVIM, two with blaVIM and blaIMP, nine with blaNDM, and one was coharbored with blaKPC. One isolate was harboring blaVIM only, two was coharbored with blaIMP, and one was harboring blaIMP only. Twenty isolates were harboring NDM only, and two co-haboured blaIMP. Finally, seven isolates were blaKPC producing. NDM, New Delhi Metallo-β-Lactamase.
Prevalence of mcr genes
A total of 80 isolates were identified to be colistin resistant using the Vitek-2 automated system. The MICs of these isolates to colistin were re-evaluated using BMD, which found 76 of those isolates to be colistin resistant. The PCR screening, however, only identified one isolate, A. baumannii BB2, to harbor an mcr-1 gene (Supplementary Table S1). None of the remaining 79 isolates were found to be harboring mcr-1, 2,3,4,5 genes. Positive controls were included in PCRs to ensure PCR validity, and amplicons are seen in Fig. 3a.
Discussion
Using PCR, we screened clinical samples stored at a clinical laboratory, NHLS, in Pretoria, South Africa. This laboratory serves other regions outside of Pretoria. We show from this molecular screening that carbapenemases are pervasive in Pretoria.
Multiple studies have also identified K. pneumoniae as the most common organism harboring carbapenemase genes.32,33 In this study, the K. pneumoniae isolates harbored all the identified carbapenemases: blaKPC, blaIMP, blaNDM, blaOXA, and blaVIM. This is concerning because carbapenem-resistant K. pneumoniae (CRKP) has been implicated in outbreaks in South Africa,34,35 and has previous reports of being endemic in certain hospitals and regions. 7
Xu et al 36 further showed in their meta-analysis that CRKP health care-associated infections have a mortality rate of 42%. It is therefore important for health care workers and South African health institutes to continually monitor this public health threat, ensuring that infection prevention and control measures are implemented to prevent the spread of CRKP.
The isolates were collected from 22 sample sources, with urine (36; 27.45%), blood (68; 22.22%), and RecSws (31; 10.13%) being the most prevalent. From blood cultures, A. baumannii, E. cloacae, E. coli, K. pneumoniae, P. aeruginosa, Salmonella Group D, P. mirabilis, and R. pickettii isolates were isolated (Fig. 1, Supplementary Table S1). This is concerning because most of these isolates are part of the ESKAPE group (i.e., Enterococcus faecium, Staphylococcus aureus, K. pneumoniae, A. baumannii, P. aeruginosa, and Enterobacter spp.), which are commonly MDR, highly virulent, and threaten public health.17,37 Furthermore, 13 of these isolates, isolated from blood cultures, were obtained from neonates.
The isolates identified from urine samples were mostly those of Enterobacteriaceae, which are common etiological agents of urinary tract infections (UTIs).38,39 UTIs are commonly identified in women, pregnant women, and immunocompromised patients, 40 and in this study, 49 of the 84 (58.33%) urine samples were obtained from women. However, due to incomplete clinical data, the clinical condition of patients is unknown.
The PCR screening showed that there is an overall higher prevalence of carbapenemase genes than mcr genes in these isolates. The Vitek 2 automated system (BioMérieux) classified 81 isolates as resistant to colistin, which was confirmed using BMD on 76 of those isolates, representing an colistin resistance sensitivity of 93.83%. Of the 80 isolates, only one (BB2) harbored an mcr-1 gene. Isolate BB2 was isolated from a fluid aspirate specimen collected from a 53-year-old female. The Vitek 2 (BioMérieux) identified BB2 to be A. baumannii complex. BB2 was also identified to be coharboring an OXA-48 gene.
In South Africa, mcr-1 genes are commonly identified in E. coli and K. pneumoniae, and this is the first report of an mcr-1–producing A. baumannii isolate in South Africa. Colistin resistance is acquired mainly through modification of the Lipid A of the LPS membrane layer through an addition of a 4-amino-4-deoxyl-1-arabinose of phosphoethanolamine.11,41,42 This is achieved through chromosomal mutations in genes encoding the two-component systems: PhoPQ and PmrAB, or mutations such as insertions or deletions that result in complete loss of the mgrB locus, a regulator of PhoPQ.
These mechanisms are commonly identified in Enterobacteriaceae species, and mgrB is commonly seen in K. pneumoniae.41,42 In A. baumannii, colistin resistance is acquired through complete loss of the Lipid A enzyme, through mutation within its biosynthesis pathway genes, lpxACD. 43 Isolates such as S. marcescens and P. mirabilis are intrinsically resistant to colistin. 42 In this study, the PCR was set up to screen for mcr-1 to mcr-5 genes, and thus mcr-6 to 10 genes would have been missed.
A total of 217 (70.9%) isolates were found to have reduced susceptibility to at least one carbapenem; that is, ertapenem, meropenem, and imipenem. Carbapenem resistance can be acquired through a combination of different resistance mechanisms that include the loss of major porin proteins, increased activity of efflux pumps, and the production of carbapenemases alone, or the hyperproduction of ESBLs and AmpCs alongside porin loss or efflux hyperactivity.44–46
In this study, the prevalence of carbapenemase genes was evaluated, and a total of 201 (65.7%) isolates were identified to harbor a carbapenemase gene. Therefore, of the 217 isolates identified to be resistant to at least one carbapenem, 201 harbored a carbapenemase gene. Thus, 16 isolates encode other resistance mechanisms that confer carbapenem resistance.
A total of 171 (55.6%) isolates harbored blaOXA-48 genes, 31 (10.1%) harbored blaNDM, 5 (1.6%) had blaVIM, 5 (2%) had blaIMP, and 4 (1%) harbored blaKPC (seen in Fig. 4). It is also seen that 80% of the blaVIM, 29% of the blaNDM, 12.5% of the blaKPC, and 40% of the blaIMP were identified to also coharbor blaOXA-48 genes (Fig. 4). These data show that there is a high prevalence and wide dissemination of oxacillinase carbapenemase in Pretoria, which correlates with the findings of Perovic et al, 15 who found that 52% of CRE isolates collected in four provinces of South Africa (Gauteng, Western Cape, KwaZulu-Natal, and Free State) were blaOXA-48 producing.
It is important to understand how common carbapenem- and colistin-resistant GNB are identified in hospitals, as these isolates include three of the five ESKAPE nosocomial pathogens. Snyman et al 47 investigated the gastrointestinal carriage of colistin-resistant microorganisms and mcr genes in healthy individuals in Cape Town's communities. 47 The study found no evidence of mcr genes, and concluded that mcr genes are not widely disseminated in the community setting. 47 The low prevalence of mcr genes elucidated in this study indicates that in this clinical setting, mcr genes are not widely distributed.
The data showed a high presence of carbapenemase genes and a low presence of mcr genes in MDR clinical GNB isolates in Pretoria, South Africa. Notably, the co-occurrence of two carbapenemases per isolate is a disturbing finding and suggests the high circulation of carbapenemases among GNB in Pretoria. Evidently, K. pneumoniae remains a cardinal MDR pathogen that requires urgent attention.
Future studies need to correlate clinical outcomes with isolated resistant organisms to better understand the true implications of the high prevalence of carbapenemases seen in these MDR organisms. Furthermore, the spread of resistant organisms in health care settings can be curtailed by effectively practicing appropriate infection prevention and control measures, and continued surveillance helps with the identification of clinical areas that require extra attention.
Footnotes
Acknowledgment
This work is based on the research supported wholly/in part by the National Research Foundation of South Africa under grant no. 131013.
Authors' Contributions
M.M. undertook laboratory work and article drafting; T.M.S.L. and L.B.S. assisted with collection of isolates and reviewing of the article; N.M.M. was a cosupervisor of the study and assisted with funding; J.O.S. designed and supervised the study and reviewed the article, as well as assisted with analysis of the data.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was funded by a grant from the NHLS given to J.O.S. under grant no. GRANT004 94809 (reference no. PR2010486).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.