
Abstract
Efflux by resistance nodulation cell division transporters, such as AcrAB-TolC in Escherichia coli, substantially contributes to the development of Gram-negative multidrug resistance. Therefore, the finding of compounds that counteract efflux is an urgent goal in the fight against infectious diseases. Previously, an efflux inhibitory activity of the antimalarials mefloquine and artesunate was reported. In this study, we have investigated further antimalarials regarding efflux by AcrB, the pumping part of AcrAB-TolC, and their drug-enhancing potency in E. coli. We show that 10 of the 24 drugs tested are substrates of the multidrug efflux pump AcrB. Among them, tafenoquine and proguanil, when used at subinhibitory concentrations, caused an at least 4- and up to 24-fold enhancement in susceptibility to 6 and 14 antimicrobial agents, respectively. Both antimalarials are able to increase the intracellular accumulation of Hoechst 33342, with proguanil showing similar effectiveness as the efflux inhibitor 1-(1-naphthylmethyl)piperazine. In the case of proguanil, AcrB-dependent efflux inhibition could also be demonstrated in a real-time efflux assay. In addition to presenting new AcrB substrates, our study reveals two previously unknown efflux inhibitors among antimalarials. Particularly proguanil appears as a promising candidate and its chemical scaffold might be further optimized for repurposing as antimicrobial drug enhancer.
Introduction
Gram-negative pathogens have become a public health threat due to the emergence of difficult-to-treat multidrug resistance (MDR). 1 The interaction of a strong influx barrier constituted by the outer membrane and highly effective multidrug efflux pumps contributes to MDR. Particularly resistance nodulation cell division (RND)-type efflux transporters, such as the pumping part AcrB of the AcrAB-TolC complex in Escherichia coli, are highly efficient in cleaning up the bacterial cell from drugs and other toxic compounds. 2
Therefore, one of the main goals of efflux research has been the development of efflux pump inhibitors (EPIs). Besides permeabilizing agents and β-lactamase inhibitors, they are promising as drug activity modulators.3,4 However, no EPI has been in clinical development so far, mostly because of compound toxicity. Thus, repurposing due to the identification of EPIs among drugs having already successfully passed clinical trials could accelerate the approval for combination therapies.
An earlier study reported a derivative of the quinolines, to which many antimalarials belong, 5 as RND-type efflux inhibitor in Enterobacter and Klebsiella isolates. 6 A later report showed another quinoline, the antimalarials mefloquine, to increase the accumulation of the fluxable dye resorufin in Pseudomonas aeruginosa and in E. coli. 7 Furthermore, the susceptibility of the latter to β-lactam antibiotics, novobiocin, and fluoroquinolones (FQs) appeared also enhanced by the antimalarial artesunate.8,9 These findings and the reported inhibition of human MDR transporters by atovaquone and quinine 10 gave reason for the present screening of antimalarials.
Materials and Methods
Escherichia coli 3-AG100 (gyrA acrR mutant) and its acrB deletion mutant ΔacrB were described previously. 11 Chemicals were purchased from Merck (Darmstadt, Germany), with the following exceptions: 1-(1-naphthylmethyl)-piperazine (NMP) was from Chess (Mannheim, Germany) and cycloguanil, dihydroartemisinine, piperaquine, and pyronaridine were from Hycultec (Beutelsbach, Germany). Standard MIC microdilution with and without adjuvants (EPI or antimalarial), Hoechst 33342 accumulation, and Nile red real-time efflux assays were carried out as described earlier12,13 with fluorescence recorded by the fluorescence plate reader Infinite M200PRO (Tecan, Männedorf, Switzerland).
The logP values were from PubChem, National Center for Biotechnology Information. 14 Chemical structures were drawn using the PubChem Sketcher. 14 Statistical analyses were performed by unpaired two-tailed t-tests with the GraphPad Prism 9.3.1 software.
Results and Discussion
AcrB-dependent efflux of antimalarials
In this study, we included 24 antimalarials from various compound classes5,15 (Table 1). First, we examined the resistance of an acrB overexpressing E. coli and its acrB deletion derivative to the antimalarials themselves. The susceptibility of the AcrB-deficient mutant to artemisinine and its derivatives dihydroartemisinine, arteether, artemether, and artesunate, to doxycycline, proguanil, pyrimethamine, tafenoquine, and to mefloquine was increased more than or equal to fourfold compared with the wild-type AcrB strain, suggesting that these compounds are efflux substrates (Table 1).
Susceptibilities of the acrB Overexpressing Escherichia coli 3-AG100 and Its acrB Deletion Mutant to Antimalarials
Precipitation at concentrations >64 μg/mL in broth.
MIC, minimal inhibitory concentration.
This has already been shown for doxycycline, 16 like for other tetracyclines, 17 and such indications have also been given for artesunate. 8 The other artemisinine derivatives as well as proguanil, pyrimethamine, tafenoquine, and mefloquine, to our best knowledge, have to be newly added to the AcrB substrate library. It should be noted that if the MICs of the antimalarials were ≥256 μg/mL for both strains (test concentrations limited due to precipitation in broth), efflux by AcrB appeared unlikely but could not be definitely excluded (e.g., if the MIC with 3-AG100 was >1,024 and that with ΔacrB 512 μg/mL).
The identification of chemically diverse AcrB substrates provides information for drug designer aiming to develop noneffluxable drugs. For example, it might be of interest that the hydrogenation of a single oxo group in artesiminine resulting in dihydroartemisinine can change the AcrB efflux ratio (quotient of the MIC of the AcrB overexpressing strain and that of its knockout mutant ΔAcrB [Table 1]).
Screening for drug-enhancing activity in E. coli
Next, we screened subinhibitory concentrations (1/4 MIC or 100 μg/mL, if MIC >256) of the antimalarials in combination with a range of antibacterial compounds in standard twofold microdilution MIC assays (Table 2). Representatives from nine antibiotic classes were used, namely of the oxazolidinones (linezolid), lincosamides (clindamycin), phenicols (chloramphenicol), β-lactams (oxacillin, cefuroxime), aminocoumarins (novobiocin), tetracyclines (tetracycline, minocycline), rifamycins (rifaximine), macrolides (clarithromycin, erythromycin, azithromycin), and FQs (levofloxacin, moxifloxacin).
Drug Sensitizing in the acrB Overexpressing Escherichia coli 3-AG100 by Antimalarials
Concentrations in micromolar shown in parentheses. If the MIC of the antimalarial in E. coli 3-AG100 was >256 μg/mL (Table 1), 100 μg/mL was used for combinational MIC testing. In all other cases, the test concentrations correspond to ¼ MIC.
Ratio of MICs in the absence and presence of an antimalarial or EPI (mean values, n ≥ 3, if ratios >2); decreases more than or equal to fourfold given bold faced; MICs of the antimicrobials alone given parenthesized in microgram per milliliter below the drug names.
AZM, azithromycin; CHL, chloramphenicol; CLI, clindamycin; CLR, clarithromycin; CXM, cefuroxime; ERY, erythromycin; LZD, linezolid; MIN, minocycline; ND, not determined; NMP, 1-(1-naphthylmethyl)-piperazine; NOV, novobiocin; OXA, oxacillin; PAβN, phenylalanine-arginine β-naphthylamide; RIX, rifaximine; TET, tetracycline.
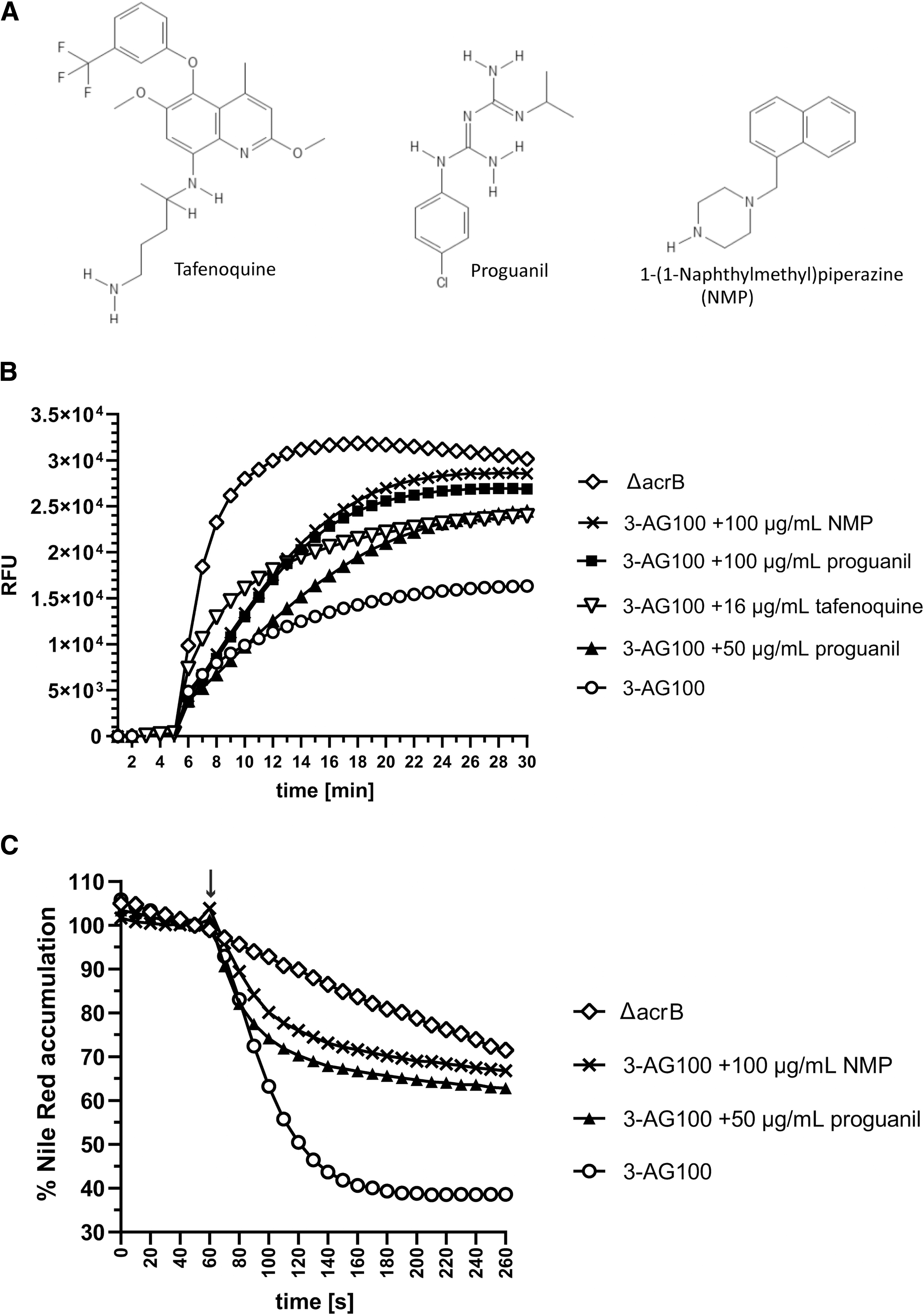
With the latter (not shown in Table 2), only proguanil achieved greater than or equal to fourfold in vitro sensitizing (Table 3). The best candidates regarding broad activity were tafenoquine and proguanil (Fig. 1A), revealing ≥4-fold MIC decreases with 6 and 11 of the antibacterial test drugs (except FQs), respectively (Table 2). These compounds including amodiaquine (activity with two drugs) were additionally tested with the highly lipophilic FQ nadifloxacin.
Impact on Fluoroquinolone Activity and on an AcrB Nonsubstrate
Ratios of MICs in the absence and presence of an antimalarial or EPI (n ≥ 3, if ratios >2); decreases greater or equal to fourfold given bold faced.
MICs of the FQs alone given parenthesized in microgram per milliliter below the FQ names.
logP values from PubChem (see “Experimental details”).
FQ, fluoroquinolone; GEN, gentamicin (aminoglycoside drug); LVX, levofloxacin (FQ); MXF, moxifloxacin (FQ); NAD, nadifloxacin (FQ); ND, not determined.
The exclusive FQ sensitizing by proguanil remarkably increased with the FQ lipophilicity, a similar finding as for the EPI phenylarginine alanine β-naphthylamide (PAβN), which, however, works at lower concentrations 18 (Table 3). It should be noted that neither proguanil nor tafenoquine shows any effect on the sensitivity to the AcrB nonsubstrate gentamicin (Table 3).
Proguanil showed broader activity, even if applied at 50 μg/mL, and acts more efficiently (sensitizing up to 16-fold) than tafenoquine (Tables 2 and 3). The efficiency of proguanil appears comparable with the well-described EPI NMP 19 with somewhat differing selectivity, but worse as found with PAβN (Tables 2 and 3). However, with the latter, an additional permeabilizing activity at least partially explains increased sensitizing to high-molecular antimicrobials such as rifamycins.18,20 Even though some permeabilization contributing to drug sensitizing cannot be totally excluded for proguanil and tafenoquine, their relatively low impact on rifamycin resistances suggests rather negligible effects on membrane integrity.
Evaluating the efflux inhibitory activity by intracellular dye accumulation
To elucidate the impact on AcrB efflux further, we monitored the kinetics of intracellular dye accumulation in E. coli cells. As shown in Fig. 1B, Hoechst 33342 accumulates in the presence of 100 μg/mL proguanil to similar extent as with 100 μg/mL NMP, meaning 1.7–1.8-fold relative to the acrB overexpressing E. coli without adjuvants, whereas it was 1.5-fold with 16 μg/mL tafenoquine and 1.9-fold due to acrB deletion (p-values ≤0.0001). In Nile red real-time efflux assays, 50 μg/mL proguanil was sufficient to achieve similar efflux inhibition as found with 100 μg/mL NMP (Fig. 1C). The impact of tafenoquine on Nile red efflux could not be examined, since its presence caused immense fluorescence decreases presumably due to quenching effects.
Taken together, among the screened antimalarials, proguanil showed broadest and most effective drug potentiating in E. coli and clearly the ability to inhibit AcrB-mediated efflux. Tafenoquine revealed less broad activity but at lower concentration. Notably, the concentrations of both compounds required are well above those reached in plasma after administration of standard doses in humans.21,22 Being substrates of AcrB themselves, which is similar to PAβN, both antimalarials do not perfectly meet the requirements of an ideal EPI. Their AcrB substrate nature suggests a competitive rather than an allosteric mode of inhibition.
Conclusions
Despite many failures, the development of RND-type efflux inhibitors still appears as a promising strategy in fighting Gram-negative pathogens. 23 The discovery of new EPIs, even if they only have experimental applicability, as well as of AcrB substrates from chemical classes other than already reported, can advance research in RND-type transport mechanisms and inhibition options. With regard to the latter, modified derivatives of tafenoquine and in particular of proguanil might be good candidates in the search for drug enhancers in Gram-negative pathogens.
Footnotes
Authors' Contributions
W.V.K. contributed to conceptualization and article reviewing; S.S. was involved in study design, experimental procedures, data analysis, article writing, and article reviewing; and M.V. contributed to experimental procedures.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.