
Abstract
The aim of this study was to clarify the biological and clinical significance of a tandem duplicate of blaVIM-24 in Pseudomonas aeruginosa ST1816 isolates. Thirteen ST1816 isolates carrying a plasmid harboring blaVIMs were obtained from two medical settings in Japan between 2016 and 2019. Complete sequencing revealed that, of the 13 plasmids, four had a tandem duplicate of blaVIM-24. These four plasmids harbored a replicon, a relaxase gene, and T4SS genes belonging to IncP-9, MOBF, and MPFT, respectively. All four plasmids transferred to PAO1 by filter mating. Cefepime marginally affected the growth of PAO1, carrying a pUCP19 harboring the tandem duplicate. Western blotting analysis showed that the relative intensity of VIM-24 metallo-β-lactamase produced by a PAO1 transformant containing a tandem duplicate was 2.6-fold higher than that produced by a PAO1 transformant containing a single copy. These results suggest that the tandem duplicate of blaVIM-24 in plasmids may confer resistance against cefepime, enabling P. aeruginosa ST1816 strains to proliferate in hospitals in Japan.
Introduction
Metallo-β-
A strain of P. aeruginosa carrying a plasmid harboring blaVIM-2 was first identified in France in 1996. 2 Since then, plasmids harboring blaVIMs genes have been detected in Enterobacterale as well as in P. aeruginosa strains in several countries. 2 In Japan, a strain of Citrobacter freundii harboring a plasmid containing blaVIM-2 was isolated in 2018. 4 In addition, P. aeruginosa strains isolated in Japan were found to carry plasmids harboring blaIMPs , but not blaVIMs. 5
Multilocus sequence typing (MLST) of P. aeruginosa strains is a useful molecular method to globally understand and identify international clones of P. aeruginosa, such as multidrug-resistant P. aeruginosa sequence type (ST) 235. 6 Compared with ST235, ST1816 has been a minor clone. 7 P. aeruginosa ST1816 isolates were first obtained in 2011 from blood samples of three children in Mexico, one each with Burkitt lymphoma, non-Hodgkin lymphoma, and Wilms tumor. 8 These isolates were susceptible to meropenem and did not harbor any gene encoding a carbapenemase. P. aeruginosa ST1816 was isolated from a sputum sample in 2014 in Japan (PubMLST). A clinical isolate of P. aeruginosa ST1816, not harboring genes encoding any MBL, was reported in 2018 in Thailand. 9
We recently reported the emergence and spread of VIM-type MBL-producing P. aeruginosa clinical isolates in Japan. 7 Findings from this study showed that carbapenem-resistant P. aeruginosa ST235 and ST1816 strains harboring blaVIMs were spreading clonally in hospitals in Japan. ST235 isolates were found to harbor blaVIMs on their chromosomes, whereas ST1816 isolates harbored blaVIMs on plasmids.
This study focused on plasmids harboring a tandem duplicate of blaVIM-24 in four P. aeruginosa ST1816 clinical isolates. The bacteriological properties of these tandem duplicates were evaluated.
Materials and Methods
Bacterial strains
Thirteen clinical isolates of P. aeruginosa ST1816, each carrying a 65 kbp plasmid harboring blaVIM genes, corresponding to blaVIM-24, blaVIM-60, or blaVIM-66, were obtained from two hospitals in Japan between 2016 and 2019. 7 Of these isolates, 12 were from hospital A in Ehime prefecture, and one was from hospital B in Hiroshima prefecture. The characteristics of these 13 isolates are shown in Supplementary Table S1, an update of Table 1 in our previous study. 7 Antimicrobial susceptibility profiles were determined using the microdilution method, according to the criteria of the Clinical Laboratory Standards Institute (M100-S30). 10 Genes associated with resistance to β-lactams, aminoglycosides, and quinolones were detected using ResFinder 3.0. Aminoglycoside resistance genes were detected using a future database 11 and in ResFinder 3.0.
Transferability of Plasmid Harboring Two Copies of bla VIM-24 to Pseudomonas aeruginosa PAO1
Complete plasmid genome sequencing
The whole genome sequences of 13 isolates were determined using Miseq (Illumina, San Diego, CA) and MinION (Oxford Nanopore Technologies, Oxford, United Kingdom), according to the manufacturers' instructions. Genomic DNAs were extracted from these isolates using DNeasy Blood and Tissue kits (Qiagen, Tokyo, Japan) for Miseq, and Genomic-tip 100/G and Genomic DNA Buffer Set (Qiagen) for MinION. Summaries of Miseq and MinION were generated using CLC Genomic Workbench v.11.0.2 and NanoStat, 12 respectively. Phred quality scores (Q scores), defined as a property logarithmically related to the base calling error probabilities were determined as described. 13 The raw reads generated by MiSeq and by MinION were assembled using Unicycler v.0.4.7. 14
Replicon types, mobility (MOB) types (classification of relaxase amino acid sequences), and mating pair formation (MPF) types [classification of type IV secretion system (T4SS) involved in MPF] of plasmids15,16 were determined using MOB-suite v.3.0.3 with a customized library that included replicon genes belonging to known incompatibility (Inc.) groups of Pseudomonas plasmids and customized parameters (—min_rep_ident 80—min_mob_ident 80—min_rep_cov 60—min_mob_cov 60). 17 The complete DNA sequences of plasmids from all 13 isolates were compared using Easyfig. These sequences were analyzed using the Basic Local Alignment Search Tool (BLAST) (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and BLAST Ring Image Generator. The blaVIM genes were detected by PCR using two sets of primers, F1/R1 and F2/R2 (Supplementary Table S2). These PCR products were sequenced using a DNA sequencer (3500xl Genetic Analyzer; Thermo Fisher Scientific).
Conjugation experiments
Plasmid transferability was tested using a rifampicin-resistant mutant of P. aeruginosa PAO1 (PAO1rifR) as a recipient strain and P. aeruginosa isolates carrying a plasmid harboring a tandem duplicate of blaVIM-24 as donor strains. Conjugation was performed in broth 18 and on membrane filters. 19 The recipient and donor strains were used at the exponential phase of growth in the presence of rifampicin (50 mg/L) and meropenem (20 mg/L), respectively. In the conjugation experiments using broth, cells were harvested by centrifugation and washed with Mueller Hinton Broth (MHB) without antibiotics, and recipient and donor cells were mixed at a 4:1 ratio and incubated at 37°C for 2 hr. In the conjugation experiments using membrane filters, 2.0 mL of recipient cultures and 0.5 mL of donor cultures were mixed and filtered through membrane filters (pore size, 0.45 μm). The membrane was placed on a Mueller Hinton agar (MHA) plate and incubated at 37°C for 2 hr. The cells on the membrane were collected into 20 mL of MHB by washing the membrane.
Transconjugants were selected on MHA plates containing both meropenem (20 mg/L) and rifampicin (50 mg/L), and recipients were selected on plates containing rifampicin (50 mg/L) alone. The conjugation frequency was defined as the ratio of colony-forming units (CFUs) of the transconjugant to those of the recipient. Transconjugant DNA was subjected to PCR using primers F1/R1 (Supplementary Table S2) to detect blaVIM-24; these DNA samples were subjected to PCR using primers F3/R3 to detect acsA, a housekeeping gene of P. aeruginosa. The PCR products were sequenced.
Transformation experiments
The open reading frames of blaVIM-24 in P. aeruginosa NCGM3596 and armA, an aminoglycoside-resistant gene, in Escherichia coli MyJU780, along with their promoter regions, were PCR amplified using specific primer sets (Supplementary Table S2). The PCR products of blaVIM and armA were digested with ShpI/BamHI and KpnI/SacI, respectively, and ligated into the plasmid shuttle vector pUCP19. 20 Plasmids harboring these PCR products were used to transform E. coli DH5α (TaKaRa Bio, Shiga, Japan). Transformants were selected on LB agar plates containing 50 mg/L ampicillin. These plasmids in E. coli DH5α were extracted and used to transform P. aeruginosa PAO1 by electroporation using an Eppenndorf Eporator (Eppendorf, Hamburg, Germany), according to the manufacturer's instructions. Transformants were selected on MHA plates containing amikacin (20 mg/L) and meropenem (20 mg/L). blaVIM in the transformants was detected by PCR using the primers F1/R1 (Supplementary Table S2).
Growth curve and time-kill assay
The transformants, PAO1 carrying a plasmid harboring armA and blaVIM-24 (PAO1::blaVIM-24) and PAO1 carrying a plasmid harboring armA and a tandem duplicate of blaVIM-24 (PAO1::blaVIM-24blaVIM-24), were cultured overnight. Aliquots containing 106 CFU/mL each were inoculated into MHB containing meropenem (32 mg/L: 1/2 MIC) or cefepime (128 mg/L: 1/2 MIC) and further incubated at 37°C with shaking at 25 rpm in a TN-2612 incubator (Advantec, Osaka, Japan). A 100-μL aliquot of each culture was harvested at 0, 1, 3, 4, 6, and 24 hr, diluted in saline, and plated onto LB agar plates. After incubation for 24 hr at 37°C, the numbers of CFU were counted. Three independent time-kill assays were performed.
Western blotting
Aliquots containing 106 CFU/mL PAO1::blaVIM-24 and PAO1::blaVIM-24blaVIM-24 were inoculated into MHB and incubated for 3 hr to the exponential phase. Each strain was further incubated for 30 min in the presence or absence of cefepime (128 mg/L: 1/2 MIC). The bacterial cells were collected, solubilized in SDS-PAGE sample buffer, electrophoresed on SDS-PAGE gels (∼ 2 × 107 CFU per 20 μL), and transferred to Immobilon-P membranes (Merck Millipore Ltd., Cork, Ireland). The Immobilon-P membranes were incubated with a mouse monoclonal anti-VIM-2 MBL antibody (RayBiotech, Peachtree Corners) and rabbit polyclonal anti-RecA antibodies (BioAcademia, Osaka, Japan). 21
The membranes were subsequently incubated with the secondary antibodies, horseradish peroxidase (HRP)-linked goat anti-mouse IgG (Abcam, London, United Kingdom) and goat anti-rabbit IgG (Abcam), respectively, with signals detected by chemiluminescence (Western BLoT chemiluminescence HRP substrate; TaKaRa Bio, Shiga, Japan). The ratios of intensities of VIM-type MBL to RecA were calculated using NIH ImageJ software (https://imagej.nih.gov/ij/). Three independent Western blotting assays were performed.
GenBank accession numbers
The whole genome sequences of P. aeruginosa ST1816 using Miseq and Minion, and the complete plasmid sequences, were deposited in GenBank under accession numbers PRJDB8977 and PRJDB10522. Novel integrons were deposited in INTEGRALL under the numbers In2000, In2001, In2002, In2003, and In2004.
Results
DNA sequence accuracy
Supplementary Table S3 shows the accuracy of whole genome sequences of 13 P. aeruginosa ST1816 isolates harboring blaVIMs. The MiSeq data for the 13 isolates consisted of 912,210 to 1,461,424 reads, of mean lengths ranging from 199.4 to 242.3 bases. The base call accuracy was high with a mean Q-score above 31, with 74.8% to 88.2% of reads having Q-scores ≥30. These Miseq data did not produce any whole DNA sequence of plasmids harboring blaVIMs.
In contrast, the MinION data consisted of 19,424 to 56,907 reads, of mean lengths ranging from 9,484.6 to 14,276.9 bases. The base call accuracy was lower compared with Miseq data, with a mean Q-score of approximately 10, with 33.8% to 84.8% of reads having Q-scores ≥10. The MinION data alone yielded incomplete plasmid DNA sequences, despite their products being long reads of sequences (Supplementary Table S3). Assembly of both the Miseq and MinION data yielded complete DNA sequences of all 13 plasmids harboring blaVIMs.
Complete DNA sequences of plasmids harboring blaVIMs
Figure 1 shows a comparative analysis of plasmid sequences harboring blaVIMs from the 13 isolates. The 12 isolates from hospital A had plasmids harboring blaVIMs, ranging in size from 64,644 to 65,553 bp with sequence similarities >98.47% (Fig. 1A). These plasmids harbored a replicon, a relaxase gene, and T4SS genes belonging to IncP-9, MOBF, and MPFT, respectively. The isolate JUPA4295 from hospital B had a plasmid 59,422 bp in size harboring blaVIM-60 and consisted of two parts. One part, consisting of nucleotides 9,514 to 28,871, had 95.46% sequence similarity to nucleotides 20,791 to 39,299 of pJUPA4001, and the second part, consisting of nucleotides −28,872 to 9,513, had 40.78% sequence similarity to nucleotides −39,300 to 20,790 bp of pJUPA4001 (Fig. 1A). This plasmid harbored a replicon gene belonging to IncP-11, whereas no relaxase or T4SS gene could be detected.

Comparative analysis of plasmid sequences harboring blaVIMs. Genome rearrangement maps of plasmids in 13 Pseudomonas aeruginosa ST1816 isolates
Of the 13 plasmids harboring blaVIM, four, pNCGM3596, pNCGM3741, pNCGM3822, and pNCGM3814_2, had two copies of blaVIM in tandem (Fig. 1A). PCR and DNA sequencing of the PCR products confirmed that pNCGM3596 carried two copies of blaVIM-24 in tandem, whereas pNCGN3661 carried a single copy of blaVIM-60 (Supplementary Fig. S1).
Analysis of genetic environments surrounding the blaVIM genes showed that all the blaVIM genes were located in one of five class 1 integrons, In2000, In2001, In2002, In2003, and In2004 (Fig. 1B). The blaVIM-24 genes were located in In2001 and In2002, followed by aadA1a (aadA1) and aacA31 (aac(6′)-31) or aadA1a. The blaVIM-60 genes were located in In2000 and In2004, followed by aadA1a and aacA31 or aadA1a, aacA31, and aadA1a. The blaVIM-66 gene was located in In2003 followed by aadA1a and aacA31 (Fig. 1B). Of the five integrons, three (In2000, In2003, and In2004, which had a copy of blaVIMs) had a promoter variant PcS with strong promoter activity. 22 The remaining two integrons (In2001 and In2002, both of which had a tandem duplicate of blaVIM-24) had PcH1 with moderate promoter activity (Fig. 1B). 22
Comparative analysis of plasmids harboring blaVIMs
The structures of plasmids of NCGM3596 obtained from Hospital A (pNCGM3596) (accession number LC586264) were similar to those of the plasmid in P. aeruginosa strain UNC_PaerCF16 (accession number CP080283), isolated in the United States in 2021 (query cover, 83%; identity, 99%); the plasmid pCCBH28525_KPC in P. aeruginosa strain CCBH28525 (accession number CP086065), isolated in Brazil in 2021 (query cover, 70%; identity, 99%); the plasmid in Pseudomonas mosselii strain DSM17497 (accession number CP081967), isolated in China in 2021 (query cover, 79%; identity, 99%); and the plasmid in Pseudomonas putida strain MX-2 (accession number CP046873), isolated in China in 2020 (query cover, 77%; identity, 99%) (Fig. 2A).

Comparative analysis of plasmids, pNCGM3596 and pJUPA4225, in P. aeruginosa NCGM3596
The plasmid from the isolate JUPA4295 from Hospital B (pJUPA4295) (accession number LC586269) had a structure similar to the plasmid pPA7790 in P. aeruginosa strain PA7790 (accession number CP015000), isolated in Brazil in 2016 (query cover, 64%; identity, 98%); and the plasmid pMS14403 in P. aeruginosa strain MS14403 (accession number CP049160), isolated in Australia in 2016 (query cover, 64%; identity, 98%) (Fig. 2B).
Transferability of plasmids harboring two copies of blaVIM-24
The four plasmids harboring a tandem duplicate of blaVIM-24, pNGCM3596, pNCGM3741, pNCGM3822, and pNCGM3814-2 (Fig. 1A), were transferred to P. aeruginosa PAO1-rifR by filter mating (Table 1). The plasmids pNCGM3741 and pNCGM3822 were also transferred by broth mating, but the other two were not (Table 1). Compared with PAO1, the transconjugants were resistant to cefepime, ceftazidime, imipenem, and meropenem, but remained susceptible to amikacin, aztreonam, ciprofloxacin, and colistin (Table 2).
Antimicrobial Susceptibilities of ST1816 Isolates Harboring a Tandem Duplication of blaVIM-24, Transconjugants and Transformants
AMK, amikacin; AZT, aztreonam; FEP, cefepime; CAZ, ceftazidime; CIP, ciprofloxacin; CST, colistin; IMP, imipenem; MEM, meropenem; RIF, rifampicin.
Kinetics of PAO1 carrying a plasmid harboring a tandem duplicate of blaVIM-24
PAO1 carrying a plasmid harboring armA and blaVIM-24 (PAO1::blaVIM-24), and PAO1 carrying a plasmid harboring armA and a tandem duplicate of blaVIM-24 (PAO1::blaVIM-24blaVIM-24) showed significantly higher MICs toward cefepime, ceftazidime, imipenem, and meropenem than PAO1 carrying a plasmid harboring armA, a vector control (Table 2). Both PAO1::blaVIM-24 and PAO1::blaVIM-24blaVIM-24 showed essentially the same MIC values toward these drugs (Table 2).
To clarify the biological significance of the tandem duplicate of blaVIM-24, growth kinetics assays were performed using these transformants (Fig. 3). PAO1::blaVIM-24 and PAO1::blaVIM-24blaVIM-24 showed similar growth curves in the absence of drug, with both starting to grow exponentially 3 hr after inoculation (Fig. 3A, B). In the presence of 1/2 MIC of meropenem (32 mg/L), both PAO1::blaVIM-24 and PAO1::blaVIM-24blaVIM-24 did not start to grow until 9 hr after inoculation, followed by exponential growth (Fig. 3A).
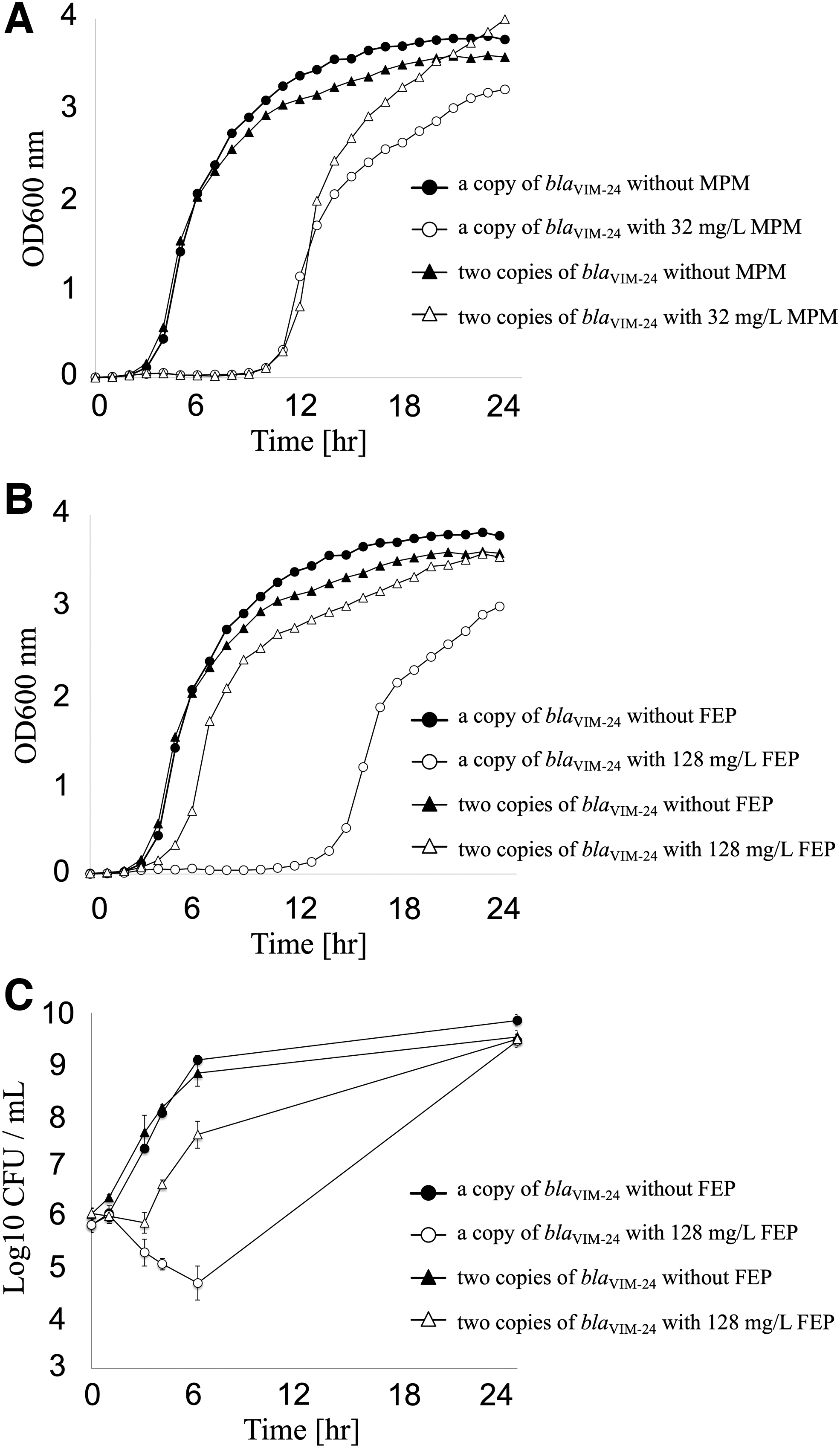
Kinetics of PAO1::blaVIM-24 and PAO1::blaVIM-24blaVIM-24 growth in the presence or absence of drugs. PAO1::blaVIM-24 (circle) and PAO1::blaVIM-24blaVIM-24 (triangle) were cultured in the presence (open symbols) or absence (closed symbols) of 1/2 MIC of meropenem (MPM; 32 mg/L)
In the presence of 1/2 MIC of cefepime (128 mg/L), PAO1::blaVIM-24 did not start to grow until 12 hr after inoculation, followed by an exponential growth, whereas PAO1::blaVIM-24blaVIM-24 started to grow exponentially 4 hr after inoculation, with the start slightly delayed compared with the same strain in the absence of drugs (Fig. 3B). Similar delay to start growth of PAO1::blaVIM-2 was observed in the presence of 1/2 MIC of ceftazidime (64 mg/L) compared with that of PAO1::blaVIM-24blaVIM-24 in the presence of the same concentration of ceftazidime (1/4 MIC), although the MICs of ceftazidime were different between these two isolates (128 and 256 mg/L, respectively) (Supplementary Fig. S2).
Counting of viable cells showed essentially the same growth curves of PAO1::blaVIM-24 and PAO1::blaVIM-24blaVIM-24 in the absence of drug (Fig. 3C). In the presence of 1/2 MIC of cefepime (128 mg/L), the number of CFUs of PAO1::blaVIM-24 declined until 6 hr after inoculation, subsequently increasing to the same number of CFUs observed in the absence of cefepime at 24 hr after inoculation (Fig. 3C). In contrast, the number of CFUs of PAO1::blaVIM-24blaVIM-24 did not change until 4 hr after inoculation, subsequently increasing to the same number of CFUs observed in the absence of cefepime (Fig. 3C).
Production of VIM-24
Western blotting analysis showed that, in the absence of cefepime, PAO1::blaVIM-24blaVIM-24 produced a larger amount of VIM-24 than PAO1::blaVIM-24, whereas the two strains produced similar amounts of RecA (Fig. 4A). Cefepime had little effect on the production of both VIM-24 and RecA by these strains (Fig. 4A). Densitometric analysis showed PAO1::blaVIM-24blaVIM-24 produced a 2.6-fold higher amount of VIM-24 MBL than PAO1::blaVIM-24 (p < 0.01), with the amount of VIM-24 MBL not enhanced in the presence of cefepime (Fig. 4B).
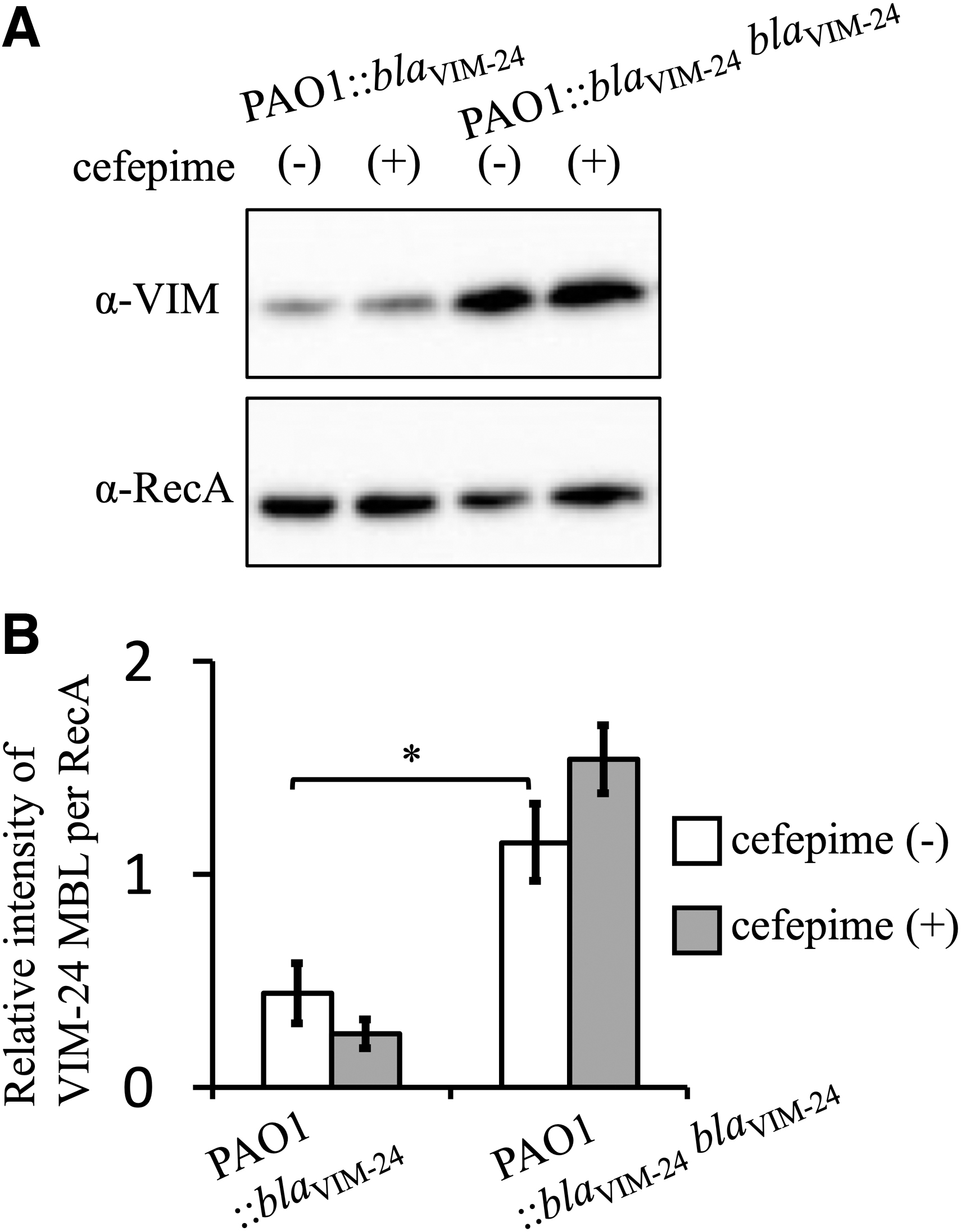
VIM-24 metallo-β-lactamase production by transformants. PAO1::blaVIM-24 and PAO1::blaVIM-24 were incubated for 3 hr and then further incubated for 30 min in the presence or absence of cefepime at 1/2 MIC. The production of VIM-24 MBL and RecA was detected by Western blotting using antibodies specific to each protein
Discussion
Acquisition of multiple copies of genes encoding carbapenemases is a strategy adopted by drug-resistant P. aeruginosa to survive and expand clonally in medical settings. This study clearly indicated that a tandem duplicate of blaVIM-24 conferred more resistance against 1/2 MIC of cefepime than a single copy of blaVIM-24, although the tandem duplicate did not change the MIC values. The tandem duplicate also conferred more resistance against sub-MICs of ceftazidime than the single copy. The results may be explained by the twofold higher production of VIM-24 in PAO1 harboring a tandem duplicate of blaVIM-24 than a single copy. A carbapenem-resistant P. aeruginosa isolate carrying a plasmid with a class 1 integron containing three copies of blaVIM-1 was reported in Spain in 2015. 23 A PAO1 transformant carrying this plasmid showed twofold higher MICs toward carbenicillin, ceftazidime, and meropenem than a PAO1 transformant carrying a plasmid harboring a single copy of blaVIM-1, although these plasmids did not have isogenic structures. 23 A carbapenem-resistant P. aeruginosa isolate carrying a plasmid harboring four copies of blaGES-5 was reported in China in 2018. 24 These four blaGES-5 copies were located within a class 1 integron, and a PA1280 transformant carrying the plasmid showed a 2.07-fold higher expression of blaGES-5 and twofold higher MICs toward piperacillin-tazobactam, cefepime, meropenem, and imipenem than a PA1280 transformant with the same plasmid harboring a single copy of blaGES-5. 24
The presence of a tandem duplicate of blaVIM-24 in four plasmids, pNGCM3596, pNCGM3741, pNCGM3822, and pNCGM3814-2, may be due to the enzymatic properties of VIM-24. In this study, the growth of PAO1::blaVIM-24blaVIM-24 was marginally affected by cefepime, but not meropenem, at 1/2 MIC. VIM-24 was first detected in a clinical isolate of Klebsiella pneumoniae, which was highly resistant to ceftazidime and cefepime, but moderately resistant to imipenem. 25 VIM-24 had higher hydrolytic activities toward cefepime and ceftazidime than toward imipenem. 26 The four plasmids harboring a tandem duplicate of blaVIM-24 may enhance the survival rates of P. aeruginosa clinical isolates in the presence of sub-MIC levels of cephalosporins, but not carbapenems.
Although all four plasmids could be transferred to PAO1 during filter mating, two of these plasmids, pNGCM3596 and pNCGM3814-2, did not transfer to PAO1 in broth. These differences may be due to MPF types. The four plasmids encoded MPFT, which forms short and rigid pili. 27 The transferability of plasmids with MPFT is more efficient on solid surfaces than in liquid. 28 The conjugation frequency of a plasmid belonging to MPFT, pWW0, was reported to be 18-fold higher during filter than broth maiting. 29
In conclusion, this study found that a tandem duplicate of blaVIM-24 on plasmids confers resistance against sub-MICs of cefepime and ceftazidime, but not meropenem, on P. aeruginosa ST1816. This tandem duplicate resulted in the spread of this strain in a hospital in Japan.
Ethical Approval
This study was approved by Biosafety Committee, 226 Juntendo University (approval number: BSL2/29-1).
Authors' Contributions
The article was seen and approved by all the authors and is not under consideration elsewhere. All the authors contributed to the work in this study. T.H. performed all experiments and wrote the initial draft of the article; T.T. designed this study and edited the article; M.T. performed genomic analyses; M.Shin. performed conjugation experiments; M.Su. conducted in silico plasmid typing; M.Shim. collected clinical isolates; and T.K. supervised this study. All authors read and approved the final article.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from Japan Society for the Promotion of Science (grant numbers: 18K07120, 20K16252, and 22K15460) and Japan Agency for Medical Research and Development (grant numbers: JP21fk0108604, JP22gm1610003, JP22fk0108139, JP22fk0108133, JP22fk0108642, JP22wm0225008, JP22wm0225004, JP22wm0225022, JP22wm0325003, JP22wm0325022, and JP22wm0325037).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.