
Abstract
Invasive fungal infections in humans are common in people with compromised immune systems and are difficult to treat, resulting in high mortality. Amphotericin B (AmB) is one of the main antifungal drugs available to treat these infections. AmB binds with plasma membrane ergosterol, causing leakage of cellular ions and promoting cell death. The increasing use of available antifungal drugs to combat pathogenic fungal infections has led to the development of drug resistance. AmB resistance is not very common and is usually caused by changes in the amount or type of ergosterol or changes in the cell wall. Intrinsic AmB resistance occurs in the absence of AmB exposure, whereas acquired AmB resistance can develop during treatment. However, clinical resistance arises due to treatment failure with AmB and depends on multiple factors such as the pharmacokinetics of AmB, infectious fungal species, and host immune status. Candida albicans is a common opportunistic pathogen that can cause superficial infections of the skin and mucosal surfaces, thrush, to life-threatening systemic or invasive infections. In addition, immunocompromised individuals are more susceptible to systemic infections caused by Candida, Aspergillus, and Cryptococcus. Several antifungal drugs with different modes of action are used to treat systemic to invasive fungal infections and are approved for clinical use in the treatment of fungal diseases. However, C. albicans can develop a variety of defenses against antifungal medications. In fungi, plasma membrane sphingolipid molecules could interact with ergosterol, which can lead to the alteration of drug susceptibilities such as AmB. In this review, we mainly summarize the role of sphingolipid molecules and their regulators in AmB resistance.
Introduction
C
Due to an increase in the number of immunogenic diseases, use of excessive immunosuppressive drugs, and prolonged use of medical devices such as respirators and catheters and aging, the prevalence of C. albicans and non-albicans species infections such as Candida tropicalis, Nakaseomyces glabrata (previously known as Candida glabrata), Candida parapsilosis, and Pichia kudriavzevii (previously known as Candida krusei) increased as major isolates, particularly in the last two decades, and is getting harder to control.6–8 Candida species produce superficial and deep mycoses, which are prevalent worldwide. 9
However, clinical resistance in Candida became a major problem, which seized the opportunity to develop new therapeutics. Amphotericin B (AmB), nystatin, natamycin, and filipin are a member of the polyene class of antifungal drugs. 10 Natamycin is given to humans for the topical treatment of fungal infections of the eyes, mouth, skin, and vagina caused by various fungi such as Candida spp., Aspergillus spp., Cephalosporium spp., and Fusarium spp.11–13 Although nystatin aids in the treatment or prevention of minor fungal infections like oropharyngeal or vaginal infections, 14 AmB is used for the treatment of serious life-threatening fungal infections.15,16
AmB was originally extracted from a culture of filamentous bacterium Streptomyces nodosus, in 1955 at the Squibb Institute for Medical Research from the soil collected in the Orinoco River region of Venezuela.17,18 Its name arises due to the amphoteric properties of that extract. Two forms of amphotericin are known, amphotericin A and AmB, but only the B form is used clinically since it was shown to be significantly more active in vivo.19,20 AmB has a long history in the arsenal of antifungal medications and is frequently administered intravenously for systemic fungal infections. AmB has been a popular treatment option for invasive infections for more than 60 years due to its wide range of activity and low levels of acquired or clinical resistance.
AmB Structural Features
AmB consists of a planar elongated macrolide ring with two distinct regions a hydrophilic region “polyol subunit” that holds several hydroxyl groups and a hydrophobic region “polyene subunit” and an amino-sugar mycosamine residue connected through a glycosidic bond. 21 The first, hydrophobic, region consists of a rigid heptaene fragment with a series of seven conjugated double bonds. 22
The second, hydrophilic, region consists of a polyol carbon chain with eight hydroxyl groups, which can form intramolecular hydrogen bonds, and a so-called “polar head” containing a hydroxyl group at the C-35, carboxyl acid residue at the C-41, and a mycosamine sugar moiety at the C-19 (Fig. 1), and due to the presence of these functional groups, AmB is a zwitterion under normal physiological conditions. Also, because of these groups, the AmB molecule exhibits amphiphilic properties. Both functional group hydroxyl at C-35 and mycosamine groups at C-19 are important for the activity, and chemical modifications of either of these groups led to new derivatives of AmB. 23 The ability of these molecules to interact and create membrane channels in a lipid environment and various supramolecular complexes in water, or at the membrane surface, as well as on the air-water interface, is due to the amphiphilic and amphoteric properties of AmB.24,25

AmB Mode of Action
The most abundant sterol in fungal cells, plasma membrane ergosterol, is the AmB's primary target for interaction and binding, which results in the formation of pores and the leakage of intracellular substances like water, solutes, and ions.24,26 Even after extensive research, AmB's action mechanism is still unknown. The widely recognized and accepted mechanism of action for AmB involves ergosterol interaction and oxidative stress induction.
Ergosterol Interaction
The various physiological processes of fungal cells, such as endocytosis, vacuolar fusion, cell signaling, membrane functional segmentation, and functioning of membrane proteins, are significantly influenced by ergosterol.27,28 AmB can bind to ergosterol and its binding with ergosterol stops it from taking part in these cellular processes, which causes fungal cell death or it can simply promote ergosterol sequestration, causing membrane stability disturbance.
Several experiments demonstrated that the primary mechanism by which AmB kills fungal cells is by binding to the membrane ergosterol known as the surface adsorption model29,30 (Fig. 2a), whereas the second mechanism is an increase in permeability through channel formation, increases the potency of the drug and accelerates the killing of yeast. Recent structural, molecular, and biophysical research demonstrate that the presence of ergosterol in the lipid membrane is critical for AmB fungicidal action. 22 AmB is an amphipathic molecule, which means that one side of the structure is hydrophobic and the other is hydrophilic.
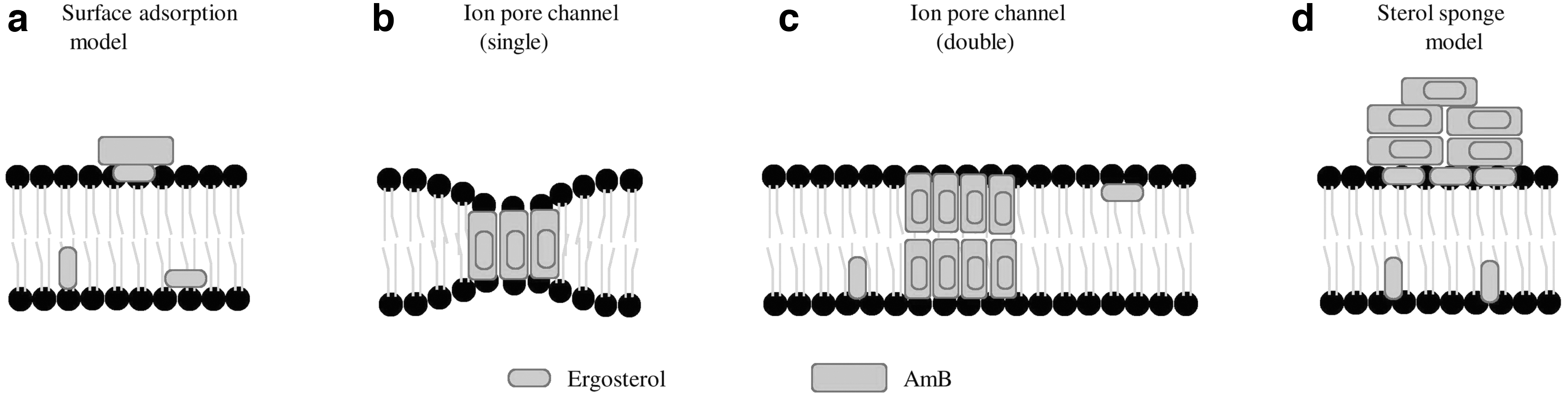
Four mechanistic models for how AmB and ergosterol interact in or near the plasma membrane:
Binding to the membrane ergosterol generates channels (pores) using hydrophobic interactions between the lipophilic portion of the polyene antibiotics and the ergosterol.10,31 AmB can form either hexameric and octameric single or double ion pores channels within the fungal cell membrane 32 (Fig. 2b, c). The pores disrupt the function of the membrane, which causes loss of membrane integrity and osmotic instability, metabolic dysfunction, and finally the death of fungal cells. It is now a widely accepted mode of action that AmB kills fungal cells by interacting with plasma membrane ergosterol and forming pores in the membrane, which causes altered plasma membrane leakage and cellular death. 33 Recently, it was shown that AmB mainly exists in the form of large, extramembranous aggregates, which kills the Saccharomyces cerevisiae and C. albicans cell by extracting ergosterol from the membrane, also known as a “sterol sponge model” 30 (Fig. 2d).
Recently, a functional group deficient AmB probe was synthesized and the interaction between the probe and ergosterol through isothermal titration calorimetry was studied, which demonstrated that the mycosamine sugar residue at the C-19 position directly interacts with membrane-embedded ergosterol and exerts in vitro activity against C. albicans and S. cerevisiae. 29
This derivative of AmB that retains the mycosamine residue at the C-19 lacks a hydroxyl group at the C-35, and retains the anionic carboxylate group at the C-41, known as C35deOAmB, binds with ergosterol, and is not able to permeabilize the membrane, but shows substantial antifungal activity. 34 However, another derivative of AmB in which mycosamine residue at the C-19 position was deleted yields a derivative known as Amphoteronolide B (AmdeB), which is unable to bind with ergosterol, is not able to form pores, and has no antifungal activity 29 (Fig. 3).

Two different functional derivatives of AmB that affect the binding of ergosterol to AmB. C35deOAmB lacks a hydroxy group at the C-35, and AmdeB lacks a functional mycosamine at the C-19 position. Mycosamine residue at C-19 is important for its antifungal activity, which is independent of its pore-forming action. AmdeB, Amphoteronolide B.
In conclusion, these studies demonstrated that AmB-mediated formation of the pore is not required to kill the yeast. Lipid membranes contain very high levels of sterols and are extremely vulnerable to permeability by AmB because of their affinity for ergosterol. 35 Due to the combined affinity of AmB for ergosterol and cholesterol, AmB indirectly affects sterols in the host membrane and disrupts its properties. The majority of AmB toxicity potential is due to its capacity to bind to cholesterol found in mammalian cell membranes, 36 despite this, AmB continues to be the major choice for the treatment of systemic fungal infections that are life-threatening, even with the advent of new antifungal medications over the past 60 years. 37
Oxidative Stress Induction
The first study that established oxidative damage as an action mechanism of AmB reported that exposure of C. albicans protoplasts to AmB under hypoxic conditions prevented protoplast lysis by more than 80% compared to the incubations in the air, and exogenous catalase and/or superoxide dismutase inhibit AmB-induced lysis of protoplast. 38 AmB is also known to cause the induction of oxidative stress in pathogenic fungi and induce an oxidative burst in the cells. 39 Several studies reported that the induction of reactive oxygen species (ROS) is a universal action mechanism of AmB. 40 Apoptosis induced by AmB treatment leads to the induction of ROS, which was found to be an additional mode of action for AmB in fungal pathogens such as C. albicans, Cryptococcus spp., and Aspergillus fumigatus. 41 AmB can function as a direct pro-oxidant and causes the buildup of ROS.
However, it is also possible that AmB must bind to ergosterol for this intracellular action to occur. AmB appears to also affect the mitochondrial activity and hence contribute to the oxidative burst because ROS are natural by-products of the respiratory chain. 42 The AmB-mediated buildup of free radicals (H2O2, OH−, and O2•) and reactive nitrogen species has several negative consequences on the cellular components such as membranes lipids, proteins, nucleic acids, and mitochondria43,44 (Fig. 4). The elevated endogenous ROS is also known to contribute to the sensitivity of the AmB-resistant isolate of Aspergillus flavus. 45 In Cryptococcus neoformans, AmB exposure induces a high oxidative burst and contributes to AmB-induced cell death besides membrane ergosterol interactions and pore formation. 46
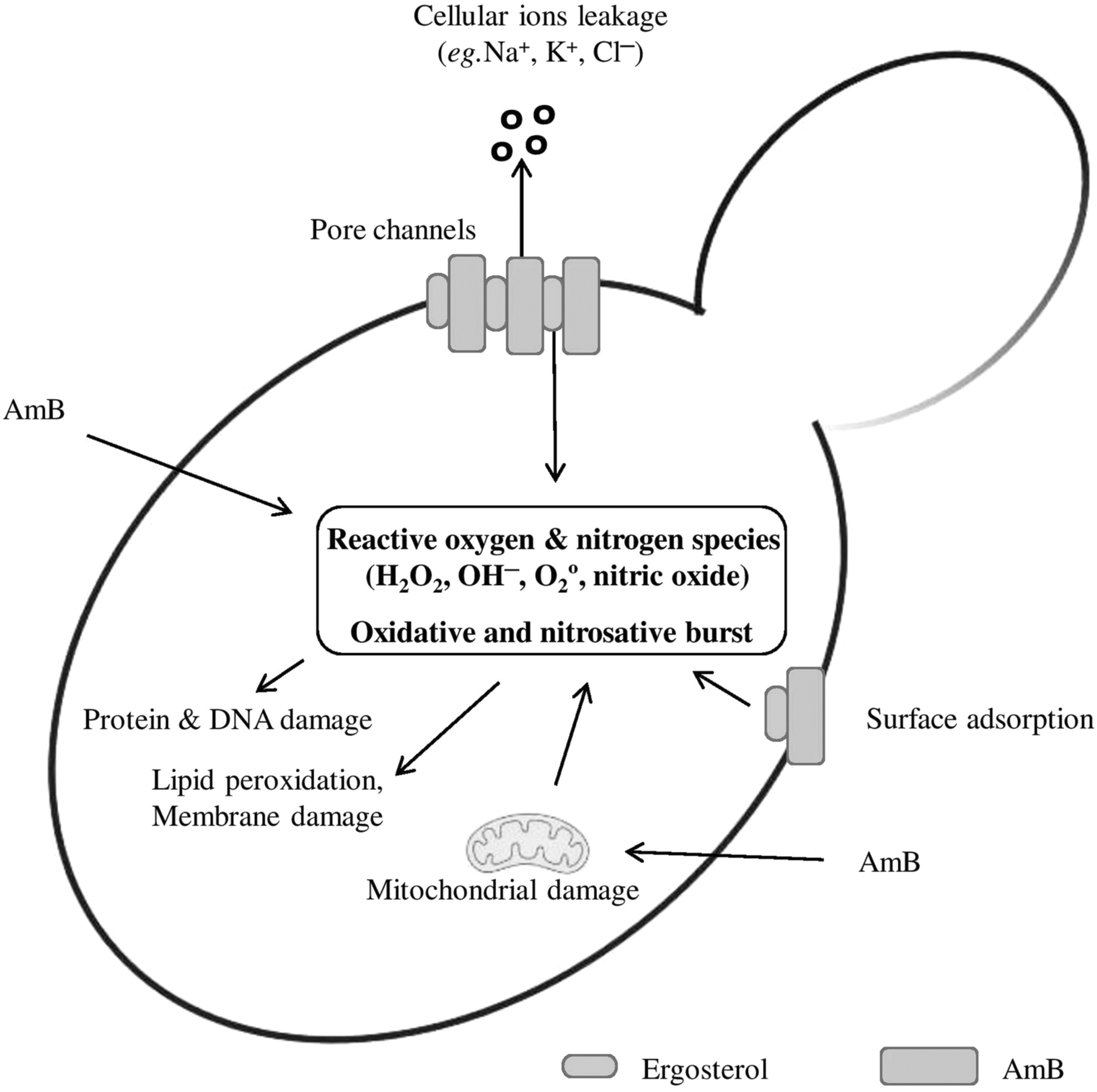
The function of the oxidative stress pathway in AmB's antifungal activity. Through direct binding with ergosterol or as a result of mitochondrial damage, AmB treatment causes the production of ROS directly. The proteins, nucleic acids, and lipid molecules found inside or in the plasma membrane of yeast cells can all be directly oxidized and harmed by ROS molecules. ROS, reactive oxygen species.
Toxicity and Lipid Formulations of AmB
A major limitation of the use of AmB in the clinic is its inherent host toxicity. Although AmB has a more than 10-fold higher affinity for fungal ergosterol than for mammalian cholesterol, nonselective disruption of mammalian cell membranes occurs. Nephrotoxicity and acute infusion-related side effects (e.g., fever and nausea) are most commonly associated with intravenous AmB, whereas liver injury can occur, but is less common. 47 The first lipid formulation of AmB, AmB deoxycholate (D-AmB, Fungizone), was developed by the U.S. Food and Drug Administration and approved for use in humans in 1959, and exhibited strong nephrotoxicity due to either direct tubular toxicity or decreased glomerular blood flow. The dose-dependent toxicity limits the maximum tolerated dose of D-AmB to 0.7–1.0 mg/kg/day, which may not be the optimal dose to achieve clinical success.
In view of the above, toxicities associated with the fungizone drug led to the development of new lipid formulations to improve the therapeutic index and reduce nephrotoxicity compared to conventional D-AmB, which has the same activity profile as the parent compound. 48 These lipid formulations included AmB lipid complex (ABLC-Abelcet), AmB liposome (L-AmB-AmBisome), and AmB colloidal dispersion (ABCD-Amphotec).49,50 The lipid composition of these formulations differed significantly and resulted in markedly different pharmacokinetic parameters, 49 for instance, ABLC, which is bigger and contains a 35% molar ratio of AmB complexed with two phospholipids dimyristoyl phosphatidyl and dimyristoyl phosphatidyl-choline in a ratio of 7:3. The usual dose of 5 mg/kg/day has been given without substantial nephrotoxicity.
However, L-AmB is a true liposomal lipid formulation of AmB and includes hydrogenated phosphatidylcholine and distearoyl phosphatidylglycerol stabilized by cholesterol, which is combined with AmB, because L-AmB is small and negatively charged, resulting in significantly greater plasma levels compared to standard AmB deoxycholate and usually administered at a therapeutic dose of 1–5 mg/kg/day. Patients receiving L-AmB typically have tissue concentrations that are greatest in the liver and spleen and significantly reduced in the kidneys and lungs. The ABCD, a stable complex of AmB and cholesteryl sulfate in a 1:1 molar ratio, displays dose-limiting, infusion-related toxicities.
Many factors, including the sort of illness such as cryptococcosis, candidosis, and histoplasmosis and the patients' backgrounds, influence the study's conclusions regarding the impacts of these lipid formulations on therapeutic effectiveness and patient mortality (e.g., AIDS and neutropenia). 47 Overall, L-AmB shows fewer infusion-related responses and exhibits less nephrotoxicity than all other formulations of AmB. 51 This reduced toxicity has allowed the delivery of increased amounts of the drug as liposomal-AmB, which is important for the treatment of fungal infections that are relatively resistant to AmB.52,53
Mechanisms of Resistance to AmB
Despite more than 60 years of clinical use, minimal resistance has developed to AmB, and the drug continues to be important in the treatment of a variety of life-threatening systemic fungal infections caused by C. albicans and A. fumigatus. 54 Some species of Aspergillus show various spectrum of drug resistance such as Aspergillus terreus and to some extent, A. flavus have a polyene-resistant phenotype; however, others spp. can show pan-resistance to available antifungal drugs. 55
According to a recent study, A. fumigatus with a transcription factor deficiency that encodes the negative cofactor 2 complex exhibits multiple drug resistance phenotypes, including resistance to azoles and AmB. 56 Although AmB-resistant strains of C. albicans (defined as a minimum inhibitory concentration ≥2 μg/mL) have been reported, AmB resistance has become more evident, because of an increase in non-albicans Candida species such as C. tropicalis, N. glabrata, P. kudriavzevii, Clavispora lusitaniae (previously known as Candida lusitaniae), and C. parapsilosis and emerging invasive mold infections that have intrinsic or acquired resistance to polyenes.57,58
Acquired AmB resistance tends to be species dependent and emerges slowly in patients undergoing AmB treatment. This has been reported in some strains of C. albicans, non-albicans Candida spp., Aspergillus, Cryptococcus, Scedosporium, Trichosporon, and Fusarium species.59–61 A surveillance study of Candida bloodstream isolates in the United States reported a 2–3% increase in the rate of AmB resistance in C. parapsilosis and P. kudriavzevii. 62 C. lusitaniae and Meyerozyma guilliermondii (previously known as Candida guilliermondii) both are reported for the rapid and inherent acquisition of AmB resistance.63–65 Acquired resistance to AmB has been also reported in C. albicans, often in concurrence with resistance to the azole antifungal agents. 66
The resistance to the AmB antifungal can occur in a variety of ways and the loss of ergosterol content is one of them. Many investigations revealed changes in the ergosterol biosynthesis pathway and sterol intermediate buildup. 63 The potential to develop AmB resistance also varies depending on the Candida species, but is higher for N. glabrata and C. parapsilosis. 67 Moreover, mutations in the ergosterol biosynthesis pathway gene (ERG) led to severe growth fitness deficits and have been associated with the AmB resistance mechanism in N. glabrata 68 (Fig. 5).
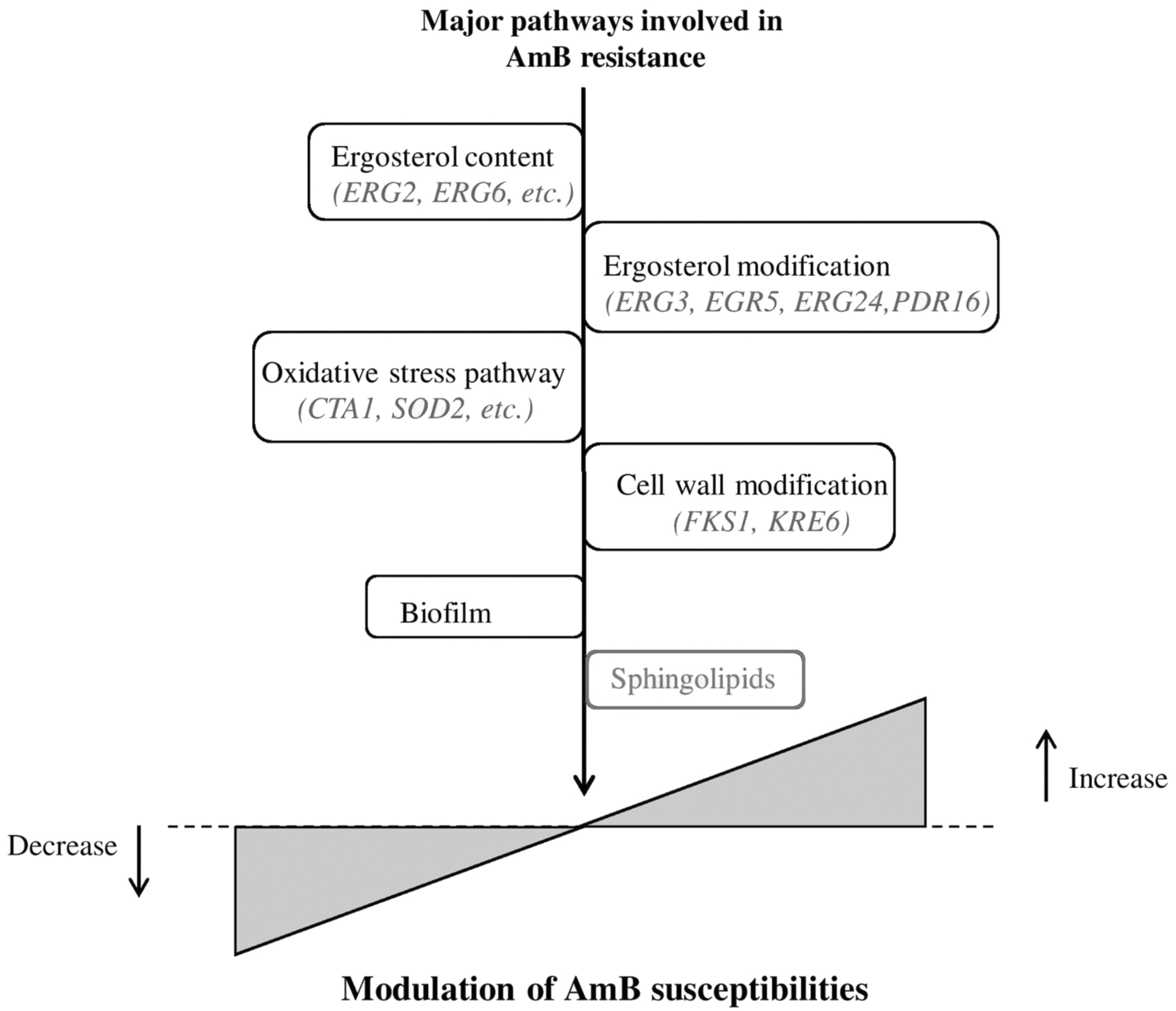
An overview of the main mechanisms reported in different fungi that contribute to AmB susceptibility. The types of ergosterol and their variations have a big impact on how AmB susceptibility changes. This figure draws attention to the newly discovered role of sphingolipids in AmB resistance.
The substitution mutation in ERG11 (lanosterol 14α-demethylase), loss in function of ERG5 (C-22 sterol desaturase), and mutational and regulatory suppression of ERG3 genes (C-5 sterol desaturase) of C. albicans lead to the synthesis of alternate sterols such as lanosterol, 4,14-dimethyl-zymosterol, and eburicol in the membrane. An earlier study demonstrated that fluconazole-resistant C. albicans isolates from the late stage of AIDS patients can show cross-resistance to AmB due to defective sterol Δ5,6-desaturation. 69 This suggests that mutational and regulatory suppression of the ERG3 gene responsible for Δ5,6 desaturation may lead to multidrug resistance, such mutations have been also reported in Candida dubliniensis. 70 Similarly, a loss-of-function mutation in ERG2 also confers resistance to azoles and AmB in C. albicans. 71 Moreover, some clinical isolates may show multidrug resistance due to simultaneous mutations in multiple genes such as ERG3/ERG11 in C. tropicalis and ERG5/ERG11 in C. albicans. 72
Moreover, in other Candida spp., such as C. lusitaniae and N. glabrata inactivation of ERG6 (C-24 sterol methyl-transferase) and ERG2 (C-8 sterol isomerase), involved in AmB resistance,63,68 in addition, a nonsynonymous mutation in ERG6 (V126F or C198F), and ERG2 (G119S or G122S) resulted in the accumulation of zymosterol and fecosterol, due to a lack of enzymatic activity by ERG6 and ERG2, respectively. Zymosterol and fecosterol can support fungal cell growth, and depletion of ergosterol in Candida cell membranes confers reduced susceptibility against AmB. 68 A loss of function mutation in the ERG2 gene is also associated with AmB resistance in C. neoformans. 73
Nevertheless, C. neoformans is also reported to be AmB resistant without substantial alteration in ergosterol biosynthesis, suggesting the ergosterol independent mechanism of AmB resistance. 74 According to its mechanism of action, the AmB performance can be affected by a drop in binding sites caused by a change or decrease in ergosterol levels in the cell membrane. Like the CaERG genes, which are necessary for ergosterol biosynthesis, any mutation in the ergosterol production pathway could impair the kinetics of this interaction. When compared to wild-type strains, C. albicans resistant to AmB and fluconazole demonstrated the involvement of CaERG2, CaERG5, CaERG6, and CaERG25 genes. The alterations resulted in an increase in sterol mediators and a reduction in AmB affinity.
Recent studies have reported that C. tropicalis strains with reduced susceptibility to AmB completely lack ergosterol in their membranes 75 ; therefore, the absence of ergosterol in the cell membrane challenges growth and survival. In these strains, an accumulation of alternate sterol molecules complements the ergosterol deficiency. Genome-wide expression analysis has also been used to investigate C. albicans resistance to AmB. 76 This study discovered that resistance to AmB and fluconazole was linked to an increase in the expression of ERG genes and the downregulation of mitochondrial enzymes like acetyl-CoA synthetase and cytochrome c oxidase, as well as the stimulation of oxidative stress genes like catalase, implying that resistance to AmB may be related to reduced mitochondrial activity and ROS. 76 Furthermore, a strong relationship between resistance to AmB and oxidant stress in C. albicans was observed; hence, strain resistant to AmB had reduced susceptibility to ROS. 77
In this work, resistance to AmB and oxidative stress is highly correlated with enhanced catalase activity. 38 However, the production of an oxidative stress level among the Candida species varies differently, such as in P. kudriavzevii, AmB induced a less proportion of ROS and displayed lower susceptibility among the more prevalent Candida species. This might be connected to differences in their cellular membrane sterol and cell wall compositions that reduce the binding of AmB to the cell membrane. 39 Alterations in mitochondrial activity and catalase between AmB-resistant and AmB-susceptible isolates have been also reported. 78
The filamentous fungus A. terreus, which is believed to be inherently resistant to AmB, provides another illustration of the importance of oxidative damage. 78 Ergosterol levels in this fungus are equivalent to those in sensitive species like A. fumigatus. On the other hand, A. terreus did not experience lipid peroxidation in response to AmB, demonstrating that the fungus had a higher capability for antioxidant activities. It was consistent that A. terreus had significantly higher catalase activity than A. fumigatus. 78
The catalase activity might break down the ROS generated upon AmB treatment and protect the cell. The mitochondria's respiratory chain is crucial in producing free radicals since they are by-products of respiration. It is tempting to connect the AmB effect and the mitochondrial activity as a result. Although little is known about this connection, it has been established that C. albicans developed increased resistance to AmB, when its respiratory system was disturbed. Changes in mitochondrial activity can have a variety of effects on AmB antifungal effectiveness because the mitochondria are necessary for the generation of ergosterol, as well as the accumulation of free radicals. 79
Biofilms are microbial communities that will grow on the attached surface and have shown reduced susceptibilities to AmB, and have been associated with reduced ergosterol content and modification in the cell wall.80–82 In addition to altered ergosterol composition and oxidative stress pathway, there is a strong correlation between AmB resistance and fungal cell wall. Previous studies demonstrated acquired AmB resistance in A. flavus isolate, which was able to grow in very high concentrations of AmB, because of an increase in the 1,3-α-glucan fraction in the fungal cell wall composition contributing to AmB resistance. 80
More recently, an enlarged cell wall with enhanced levels of 1,3-β-glucan in AmB-resistant C. tropicalis isolate was also reported. 83 Another study reported altered cell wall chitin content responsible for AmB resistance in Kluyveromyces strains, 84 although the enhanced glucan biosynthesis could be the result of altered regulatory pathways upon AmB exposure, which can inhibit the penetration of AmB through the cell wall.
Recently a comparative transcriptomics study in Candida auris revealed that the AmB-resistant isolates display high resistance to cell wall-perturbing agents, including calcofluor white, congo red, and caffeine. 85 C. auris, a new multidrug-resistant yeast pathogen isolated from a female patient in Tokyo, Japan, in 2009, causes invasive infections with very high mortalities and morbidities.86,87 Most clinical isolates of C. auris are resistant to azoles (∼70%), echinocandins (∼8%), and even to AmB (∼35%).88,89 To date, only ergosterol biosynthesis genes (i.e., ERG1, ERG2, ERG6, and ERG13) due to increased expression 90 or ERG6 amino acid substitution YY98V (MIC >32-fold increase) lead to the synthesis of cholesta-type sterol instead of ergosterol have been reported to provide AmB resistance in C. auris.88,91
Notably, single-nucleotide polymorphism, a nonsynonymous mutation in utg5_821828 (C/T), resulted in an amino acid change from serine to asparagine in a gene predicted to encode a transcription factor similar to FLO8 in C. albicans, and a mutation in utg4_968953 (T/C) results in a change from the hydrophobic amino acid isoleucine to the polar amino acid threonine in a gene predicted to encode a membrane transporter, which has been reported to induce AmB resistance in C. auris. 92 In addition, C. auris is also able to form biofilms that have been suggested to provide resistance to common antifungal therapies. 93 Figure 5 provides a summary of the various modes of the AmB resistance mechanism.
Sphingolipid Biosynthesis Pathway and AmB Resistance
In S. cerevisiae, the endoplasmic reticulum (ER) and Golgi apparatus are involved in the sphingolipid biosynthesis pathway. 94 The first reaction in sphingolipid biosynthesis is the serine palmitoyl transferase (SPT)-catalyzed condensation of L-serine and palmitoyl-CoA in the ER, which is the rate-limiting step 95 (Fig. 6). The SPT is a heterodimer of two subunits Lcb1 and Lcb2, and associated with Tsc3 (a minor subunit), which is required for maximum activity. 96 3-ketodihydroshingosine (3-KDS), the first by-product of this process, is further converted by 3-keto reductase (Tsc10) into dihydrosphingosine (DHS), utilizing NADPH as a cofactor. DHS is further metabolized into phytosphingosine (PHS; a chief long-chain base in yeast and affects cell growth upon accumulation) by Sur2 using NADPH as a coenzyme. 97

Diagrammatic representation of the Saccharomyces cerevisiae sphingolipid biosynthesis pathway and its regulator, which are involved in AmB resistance. Complex sphingolipids are synthesized in the Golgi apparatus, whereas long-chain bases are produced in the ER. Each step's relevant genes are highlighted in italics. ER, endoplasmic reticulum.
PHS and DHS then further condensed with very long-chain fatty acyl-CoA produced by fatty acid elongase (Fen1 and Sur4) to form phytoceramide and dihydroceramide, respectively, in the presence of ceramide synthase (Lag1, Lac1, and Lip1). 98 Sur2 is also involved in the conversion of dihydroceramide to phytoceramide. Moreover, Lcb4 and Lcb5 phosphorylate long chain bases such as DHS, and PHS are responsible for the formation of DHS-1 phosphate and PHS-1-phosphate, respectively, whose accumulations are toxic to the cells. 99 In addition, Dpl1 converts both these substrates to ethanolamine. 100
Phytoceramide is synthesized in ER and transported to the golgi apparatus for further synthesis of complex sphingolipids. 101 The COP-II vesicle-mediated transport of ceramides to golgi needs oxysterol-binding proteins. 102 Phytoceramide is converted by Aur1 encoding IPC synthase into inositol phosphoryl ceramide (IPC). Afterward, MIPC synthase encoded by Sur1 adds mannosyl residues to IPC to form MIPC. Csh1 and Csg2 (Ca2+-sensitive mutants) are other subunits of MIPC synthase. CSH1 has functional as well as sequence similarity to Sur1, whereas Csg2 has no sequence similarity to the MIPC synthase reactive site; however, it encodes the regulatory subunit of MIPC synthase.
Further modification of MIPC takes place in the golgi apparatus by insitolphosphotransferase 1 (IPT1), which results in the synthesis of M(IP)2C. The complex sphingolipids are transported to the plasma membrane by the COP-II vesicular transport mechanism and perform various functions. 103 They physically associate/interact with sterols and glycerophospholipids and form the important constituents of lipid raft. 104
A high AmB resistance without significant alterations or modification in ergosterol has been reported in the experimentally evolved C. neoformans, suggesting the ergosterol-independent AmB resistance mechanism. 74 Recently, a study performed on the sphingolipidomics of drug-resistant isolates of C. auris suggested the existence of altered sphingolipid profiles can also influence AmB susceptibilities in C. auris. 105 Moreover, it has been discovered that the fatty acid elongase (FEN1 and SUR4) gene deletant of S. cerevisiae, and its homolog in C. albicans (CaFEN1 and CaFEN12), are defective in the biosynthesis of very long-chain fatty acyl-CoA, increases AmB drug sensitivity and supplementation of PHS a sphingolipid biosynthesis intermediate restores the AmB susceptibilities of myriocin treated cells, which suggests a direct role of sphingolipid molecules in AmB resistance. 106 This study was the first to provide a direct link between sphingolipids and AmB resistance.
Sphingolipid Biosynthetic Regulators in AmB Resistance
The biosynthesis and degradation of sphingolipid molecules should be maintained for normal cellular homeostasis. 107 Sphingolipid biosynthesis regulators mainly exist as a complex with SPT, known as SPOTS complex, comprises of Orm1, Orm2, and Sac1, in addition to Lcb1, Lcb2, and Tsc3 in S. cerevisiae. 108 This complex is well recognized for controlling the initial and rate-limiting step in the biosynthesis of sphingolipids. Each component of this complex exhibits significant physical interactions, which are supported by affinity purifications and mass spectrometry in S. cerevisiae. 96 Orm1/2 and Sac1 both interact with SPT and are responsible for its negative regulation. 96
However, both bind to the SPT enzyme independently and their mechanisms of regulation are also distinct. Orm1 and Orm2 inhibit the sphingolipid biosynthesis pathway by interacting with SPT through its Lcb1 subunit. 109 Sac1 also interacts with SPT through Tsc3, resulting in the enzyme's negative regulation. Together, this complex regulates sphingolipid biosynthesis, and mutations in any one of these genes resulted in impaired sphingolipid metabolism. 110
In S. cerevisiae, PDR16 (phosphatidylinositol transfer protein) is involved in regulating complex sphingolipid biosynthesis by interfering with the action of AureobasidinA (AbA), the inhibitor of AUR1, which catalyzes the synthesis of IPC from ceramide. 111 In S. cerevisiae, overexpression of PDR16 confers resistance to AbA by suppressing the growth defect and alleviates the reduction in complex sphingolipids in AUR1-repressed cells by AbA treatment. 112 However, the resistance conferred by overexpression of PDR16 was not observed in cells in which a tetracycline-regulatable promotor was used for the repression of AUR1 and even worsen the growth defect in these cells, unlike AbA-treated cells on PDR16 overexpression.
These findings suggest that PDR16 overexpression alleviates the effect of AbA and enhances cell survival. 112 In a genome-wide approach for exploring drug resistance mechanisms, it has been reported that overexpression of PDR16 in S. cerevisiae and C. albicans also provides resistance to AmB. 76 Resistance conferred by PDR16 overexpression might be due to changes in membrane permeability and membrane lipid composition because PDR16 also alters membrane ergosterol composition. 113 Similarly, another study reported plasma membrane proteolipid 3 (PMP3), a nonessential gene in S. cerevisiae that is highly expressed under high salt concentration, as well as low-temperature conditions, modulates AmB resistance.114,115
These studies further demonstrated that the PMP3 modulates sphingolipid biosynthetic pathways, which lead to AmB sensitivity in PMP3 deletants of S. cerevisiae, N. glabrata, and C. albicans. However, the mechanism by which PMP3 regulates sphingolipid metabolism is still unclear.115,116 PMP3-mediated increased resistance to AmB is decreased (2-fold) in S. cerevisiae cells lacking Fen1 and Sur4, the sphingolipid biosynthetic genes, or Ypk1 and Sac1, the sphingolipid regulatory gene, in comparison to wild-type cells. This provides the preliminary evidence that PMP3-mediated increase in AmB resistance requires active long-chain sphingolipid biosynthetic machinery. 106
In addition, PMP3 knockout cells showed increased sensitivity to myriocin, an SPT inhibitor in S. cerevisiae, whereas sensitivity to these drugs is decreased in PMP3-overexpressing cells. In addition to this, growth defect in PMP3-deleted cells in the presence of myriocin or AmB is reversed in PHS-supplemented cells, which is the sphingolipid biosynthetic pathway intermediate. 115 These results provided evidence that PMP3 by modulating the sphingolipid metabolic pathway increases resistance to AmB. Figure 7 provides a summary of the role of sphingolipids and their regulators in the modification of AmB susceptibilities.

A diagram showing the emerging role of regulatory genes (PDR16 and PMP3) and specific sphingolipid biosynthesis genes (FEN1, SUR4, and IPT1) that are involved in changing AmB susceptibilities in Saccharomyces cerevisiae and Candida albicans. PMP3, plasma membrane proteolipid 3.
Conclusion and Future Prospectus
The role of sphingolipid and its regulators in fungi, along with all the known pathways that modulate AmB drug resistance, was summarized in our review. However, there are still many unrecognized details about the function of sphingolipid and its regulators in AmB drug resistance, which demands further study. Over the past few decades, AmB has been widely used to treat a variety of fungal infections, but its exact mechanism of action is still unknown, although the studies demonstrated that AmB binds to sterols and forms pores at the plasma membrane, and induces oxidative damage in the cells. The induction of antioxidant enzymes or a decreased ergosterol content are two pathways that have been linked to AmB drug resistance; in some cases, both these pathways can affect an individual's susceptibility to the drug. To clearly define the importance of each of these mechanisms, more research is necessary.
In addition, the fact that AmB is a medication with stronger fungicidal activity is consistent with the induction of numerous pathways to kill the fungi. It is unclear how AmB interacts with the pathogen or host during infection, given the immunomodulatory effects of antifungal medications. Due to the immunosuppressed nature of patients with systemic fungal infections, this problem is very intriguing. As a result, the AmB treatment may have unanticipated effects on patients, depending on their immunological conditions. In the event of a deadly systemic fungal infection, despite its well-established track record, its well-known adverse effects and toxicity may necessitate treatment withdrawal. An ergosterol molecule is well known to be involved in the AmB resistance mechanism.
However, recent studies highlight the crucial role of sphingolipid molecules. Sphingolipids might be the major player in drug resistance of AmB since the two classical genome-wide studies on AmB drug resistance identified the role of sphingolipid biosynthesis pathway genes PMP3 and PDR16 in drug resistance.76,117 Mutants defective in the sphingolipid metabolic pathway affect growth inhibition by AmB drugs, but the exact molecular mechanism is poorly understood. 118 Particularly, there is a strong interconnection between sphingolipids and sterols in yeast. 119 Classical genetic and high-throughput studies in the yeast S. cerevisiae also gave preliminary evidence of possible genetic interactions between genes involved in the sterol transport pathway, as well as the sphingolipid metabolic pathway.119–122 Characterizing the nature of these interactions will further help in finding out clear molecular mechanisms.
Sphingolipid biosynthesis serves as a potentially useful target because of its involvement in various pathways such as fungal virulence, pathogenicity, cell growth, and replication. There is an urgent requirement for the discovery of novel antifungal therapeutics that would help to control the emerging cases of fungal AmB resistance and have a nontoxic attribute to the mammalian cells in immunocompromised patients. Sphingolipid biosynthesis and its regulators are now well known to modulate antifungal drug resistance such as AmB in both S. cerevisiae and C. albicans (Table 1).
List of Sphingolipid Biosynthetic and Regulatory Genes Involved in Amphotericin B Susceptibility
AmB, amphotericin B; PMP3, plasma membrane proteolipid 3.
We suggest that this AmB resistance might involve Orm proteins, which are master regulators of the sphingolipid metabolic pathway. 123 They regulate SPT by interaction with the N-terminal domain of Lcb1, but which domain of Orm protein is essential for regulatory purposes and other interacting partners of Orm protein is yet to be investigated. Possible investigation of the detailed mechanism of sphingolipid-mediated AmB resistance will highlight a potential and novel role of sphingolipid biosynthetic and regulatory genes in AmB resistance.
Footnotes
Ethical Approval
This article does not contain any study with human participants or animals by any of the authors.
Consent for Publication
Written informed consent for publication was obtained from all the authors.
Authors' Contributions
K.M. prepared initial draft and V.K.B. conceptualized, finalized the content and wrote the article.
Disclosure Statement
The authors declare no competing financial or nonfinancial interests.
Funding Information
The Science and Engineering Research Board (SERB), Department of Science and Technology, Government of India, provided financial support for this work (Award No. SRG/2020/000171/LS). K.M. is appreciative of SERB's, India, financial assistance as a fellowship for a summer internship under SSR initiatives. We also acknowledge the research funding provided by the Central University of Punjab, Bathinda's RSM grant, UGC's Start-Up grant letter number F. 30-583/2021(BSR), and ICMR-Adhoc grant number 2021-8947 awarded to V.K.B. The decision to submit the article for publication or the preparation of the article was both made independent of the funding organizations.