
Abstract
Purpose:
Chryseobacterium indologenes is a clinically relevant microorganism that has been on the rise, with multidrug-resistant (MDR) strains being reported. C. indologenes carrying tet(X2) has been demonstrated to be resistant to the antibiotic tigecycline, yet, sensitive to all other members of the tetracycline family. This inconsistency in resistance prompts an inquiry into the contribution of tet(X2) to tigecycline resistance in C. indologenes.
Materials and Methods:
In this study, we report on a comprehensive analysis of the genomic mechanisms underlying tigecycline resistance in a MDR C. indologenes strain (CI3125) that was resistant to tigecycline but sensitive to tetracycline, doxycycline, and minocycline. We used whole-genome sequencing, quantitative reverse transcription PCR, Western blot, antibiotic-degrading tests, and efflux pump inhibiting tests to reveal the mechanism of tigecycline resistance in C. indologenes and elucidate the inconsistency in the antibiotic resistance mechanism for the tetracycline family.
Results:
Our findings demonstrate that CI3125 carries 60 antibiotic resistance genes distributed on 6 different genetic islands (GIs), with the potential for horizontal transfer. Notably, the tet(X2) gene is located on GI06 of CI3125. Genetic environment analysis of tet(X2) showed that all tet(X2) genes in Flavobacterium and Bacteroides share a conservative and functional ribosome-binding site upstream. Contrary to expectation, our RT-qPCR showed that tet(X2) was not transcribed in CI3125, and Western blot suggested the absence of tet(X2) protein in CI3125. Rather, we demonstrate that minimum inhibitory concentration values for tigecycline decreased two- to eight
Conclusions:
Overall, our results highlight the importance of genomic approaches in understanding the underlying mechanisms of antibiotic resistance in clinically relevant microorganisms. While tet(X2) in CI3125 is silent, our findings suggest that it may be horizontally spread through GIs. Hence, our findings have significant implications for the management of C. indologenes infections in clinical settings.
Introduction
Tigecycline, the third-generation tetracycline, was approved by U.S. Food and Drug Administration in 2005. 1 Tigecycline specifically binds to bacterial ribosomes, interfering with aminoacyl-tRNA and halting bacterial protein synthesis. Tigecycline has a unique structure, in which a 9-tert-butyl-glycylamido side chain modification structure overcomes the efflux pump mechanism [tet(A–E), tet(K)], ribosome-protection mechanism [tet(M)], and other mechanisms of resistance seen with previous tetracyclines (Linkevicius ea al. 2016). 2 Tigecycline is particularly efficacious against multidrug-resistant (MDR) gram-negative bacteria, and is thought as the last-resort for bacterial gram-negative infections. 3
Tigecycline resistance in bacteria primarily arises from four mechanisms: altered cell membrane pore channels, ribosome protection, efflux pumps, and drug-degrading enzymes. Within gram-negative bacteria, overexpression of efflux pumps of the major facilitator superfamily and resistance-nodulation-cell (RND) families, including TET(A), AcrAB-TolC, OqxAB, and AdeABC, contributes significantly to resistance. 4 Among these pumps, TET(A) and AcrAB-TolC have been most extensively studied. 5 The FAD-dependent monooxygenase tet(X) can degrade nearly all tetracycline antibiotics in the presence of FAD, NADPH, Mg2+, and O2. 6
Although the tet(X) gene was first isolated from the anaerobic Bacteroidetes, the current study points the origin of the tet(X) to R. anatipestifer, the tet(X) and its variants share the same ancestry with the monooxygenase gene carried in the chromosomes of Flavobacteriaceae bacteria.7,8 The discovery of tet(X) and its variant tet(X2) in Bacteroides highlighted the potential for tigecycline degradation, although resistance was relatively low; however, these effects persisted when the gene was transferred to Escherichia coli. 9 Distinct variants of the tet(X) gene provide varying levels of tigecycline resistance, with tet(X3) and tet(X4) genes mediating high-level resistance with minimum inhibitory concentration (MIC) values of 32–64 mg/L. 10 TET(X4) is commonly found in mobile plasmids and, occasionally, in chromosomes. 11
Chryseobacterium indologenes, a member of the genus Chryseobacterium belonging to the Flavobacteriaceae, is widely present in soil, water, and nosocomial environments and is an emerging nosocomial opportunistic pathogen in critically ill patients. 12 In addition, cases of healthy infants and community-acquired infections caused by C. indologenes have been recently reported. 13 Although it lacks typical virulence factors, C. indologenes is often resistant to most clinical antimicrobial agents, particularly tigecycline.14,15 C. indologenes strains are often reported to carry tet(X2), which is generally thought to be responsible for tigecycline resistance.7,16 Zhang et al., isolated 40 tigecycline-resistant C. indologenes strains carrying tet(X2) throughout China. 7 Interestingly, all of these tigecycline-resistant isolates were susceptible to minocycline. 16 The inconsistent resistance to the tetracycline family raises the question of whether tet(X2) mediates tigecycline resistance.
Whole-genome sequencing can provide more information about antibiotic resistance genes and may explain the mechanism of antibiotic resistance. However, few studies have provided detailed analysis on the whole-genome sequence and antibiotic resistance mechanism of C. indologenes. This study reports the complete genome sequence of a clinically isolated C. indologenes strain, 3125 (CI3125), which is resistant to tigecycline but sensitive to minocycline and doxycycline. We also performed experiments to reveal the mechanism of its multidrug resistance. Our study may deepen the understanding of the clinical characteristics of C. indologenes.
Materials and Methods
Bacterial culture and preliminary identification
CI3125 was isolated from a sputum sample of a patient suffering from nephrotic syndrome with severe pneumonia. CI3125 was cultured on blood agar plates at 30°C or 37°C for 24 hours, respectively. The VITEK 2 Compact automatic microbial analysis system was used to identify the bacterium genus and detect the antimicrobial susceptibility of CI3125. Motility assay was performed as described by Rashid et al. 17
Genome sequencing and analysis
Bacterial genomic DNA was extracted (Takara MiniBEST Bacterial Genomic DNA Extraction Kit Ver. 3.0) and was subjected to whole genome sequencing through a combination of the Illumina HiSeq 4000 platform and the PacBio RS II platform by Majorbio Corporation (Shanghai, China). Pbdagcon was used for self-correction, and the genomic draft was assembled using the Celera Assembler. A single-base correction was available through the GATK and SOAP tool packages (SOAP2, SOAPsnp, SOAPindel). The genome size was estimated by K-mer analysis before assembly.
MEGA6, CSIPhylogeny, and iTOL were used to build phylogenetic trees. Rapid Annotation using Subsystem Technology (RAST) online bioserver was used to annotate the genome. tRNA and rRNA genes were verified using tRNAscould-SE and Barrnap, respectively. PHAge Search Tool (PHAST) was used to predict the phage region.
The insertion sequences were predicted by ISEScan. Genetic island was verified using Islandview 4. The ICE elements were predicted using ICEberg. MAUVE was used to perform genome colinear alignment. Protein function was predicted using Cluster of Orthologous Groups of proteins (COGs). Sequence alignment was performed using BLAST+ (https://ftp.ncbi.nlm.nih.gov/blast/executables/blast+/LATEST/), BRIG, and Easyfig was used for visualization.
CARD database was used to verify the resistome of CI3125. Antibiotic resistance genes were predicted based on CARD databases. PromPredict was used to predict the promoter. The genome sequence of CI3125 was uploaded onto NCBI (National Center for Biotechnology Information) under the accession number NZ_CP067044 (Assembly: GCF_016694815.1).
Antimicrobial susceptibility testing
Drug susceptibility system analyzer (bioMerieux, Marcy l'Etoile, France) was used to detect the antimicrobial susceptibility of CI3125. Broth microdilution assay was used to determine the MIC value for tigecycline. 18 Results were interpreted according to the breakpoints according to the Clinical and Laboratory Standards Institute (CLSI). 19 Susceptibility/resistance breakpoints of tigecycline were interpreted according to European Committee on Antimicrobial Susceptibility Testing (EUCAST) criteria (susceptible: ≤1 μg/mL, resistant: ≥4 μg/mL). 19
Gene cloning
Constitutive expression vectors of tet(X2),and tet(X4) were constructed by replacing the open reading frame (ORF) of the tetR in pBR322 using the infusion cloning method (Takara In-Fusion HD Cloning Kit). 20 tet(X4) gene was directly synthesized (Beijing Genomics Institute BGI Co., Ltd.). tet(X2) and tet(X4) gene were PCR amplified, and was cloned into pBR322 to produce plasmid pBR322-tet(X2) and pBR322-tet(X4). After being digested with DpnI, plasmids were transferred into E. coli DH5 using the method of chemical transformation.
Quantitative reverse transcription PCR for transcription of tet(X2)
The total RNA of CI3125 was extracted using RNAfast200 Kit (FASTAGEN Co., Ltd.). Reverse transcription and qPCR assay were performed using TaKaRa PrimeScript TM RT Reagent Kit and TB Green Premix Ex Taq II Kit (Takara Co., Ltd.).
Preparation of tet(X2) monoclonal antibody and Western blot assays
CI3125 genome contained 327 bp length hypothetic ORF predicted as a tet(X)-like gene, which shares about 70% identity with tet(X2). When preparing for constructing the tet(X2) expression vector, the first 327 bp of tet(X2) ORF was skipped to avoid cross-reaction in Western blot assays. The truncated tet(X2) was cloned between the NcoI and XhoI sites in pET32a. Protein was purified as described by Cheng et al. 21 (Supplementary Fig. 3). Routine rabbit immunization with purified truncated tet(X2) and purification of the polyclonal antibody against tet(X2) was performed as described by Xu et al. 22 Western blot 23 was performed to identify the polyclonal antibody.
Analysis of ribosome-binding site
tet(X2) ORF and its upstream 18 bp sequence were PCR amplified using CI3125 genome as template. The PCR product was cloned into pBR322, and placed immediately after the termination codon of ampR using an In-Fusion Cloning Kit (Takara In-Fusion HD Cloning Kit). The constructed vector (pBR322-tet(X2-RBS)) was transferred into E. coli DH5α using the chemical transformation method. DH5 containing pBR322-tet(X2-RBS) was cultured on an liquid broth agar (LB) plate containing 2 μg/mL tigecycline to identify the expression of tet(X2).
Tigecycline degradation assays
CI3125 and DH5ɑ-tet(X4) were cultured on the LB agar plates at 30°C and 37°C for 18 hours, respectively. Bacterial colonies were harvested and suspended in LB medium and diluted to a concentration of 3 × 108 CFU/mL. Tigecycline was added into bacterial suspension to a final concentration of 8 μg/mL. The appropriate bacterial rest was sampled and centrifuged at 12,000 g for 5 min, and the supernatant was filtered through a 0.22-μm filter and was subjected to scan at a wavelength range of 300–700 nm in 8 hours.
Efflux pump inhibition assays
MICs using the broth microdilution method were assayed for tigecycline in the presence of the following efflux pump inhibitors: carbonyl cyanide 3-chlorophenylhydrazone (CCCP), phenyl-arginine-β-naphthylamide (PAβN), 1-(1-naphthyl- methyl)-piperazine (NMP), reserpine, and verapamil (Sigma). These inhibitors were added to the broth at final concentrations of 5, 70, 25, 50, and 100 μg/mL, respectively. A significant inhibition effect was defined as a four-fold or greater decrease in the MIC values. 24
Results
The phenotypic properties of CI3125
CI3125 bacterial colony on the blood agar plate cultured at 30°C for 24 hours was pigmented yellow and without hemolysis. CI3125 showed swarming and swimming motility (Supplementary Fig. S1). CI3125 is resistant to most clinical antimicrobials (Table 1). Interestingly, it is resistant to tigecycline (MIC ≥8 μg/mL) but susceptible to minocycline (MIC ≤1 μg/mL) and doxycycline (MIC = 4 μg/mL).
Antibiotic Susceptibility Test in the CI3125
AMK, amikacin; ATM, aztreonam; CAZ, ceftazidime; CIP, ciprofloxacin; COL, colistin; DO, doxycycline; FEP, cefepime; IPM, imipenem; LEV, levofloxacin; MEM, meropenem; MNO, minocycline; SFP, cefoperazone/sulbactam; SXT, trimethoprim/sulfamethoxazole; TCC, ticarcillin/clavulanic acid; TGC, tigecycline; TOB, tobramycin; TZP, piperacillin/tazobactam.
General features of CI3125 genome
The phylogenetic tree based on 16 s rRNA shows that CI3125 belongs to C. indologenens (Fig. 1A). We chose 12 complete genome sequences of C. indologenes in the NCBI database to align with that of CI3125. The phylogenetic tree based on genomic single-nucleotide polymorphisms shows that C. indologenes strains FDAARGOS 510 and C. indologenes strain H5559 are the closest strains to CI3125 (Fig. 1B).
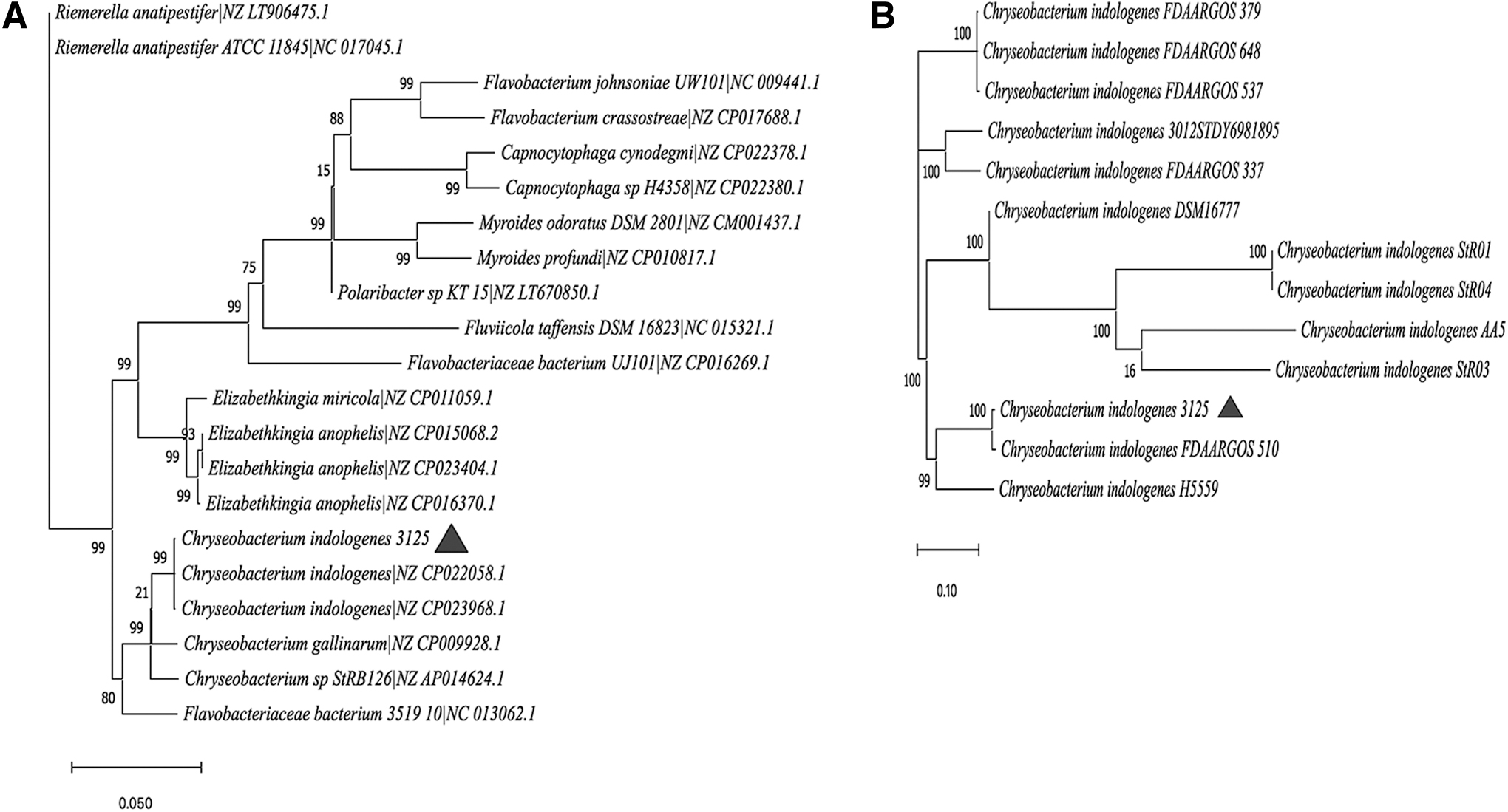
Phylogenetic inferences were obtained using the neighbor-joining method. The branches of the tree are indicated by the genus and species name of the type strains followed by the NCBI gene accession numbers.
CI3125 has a genome length of 4,928,298 bp with a GC content of 37.2% and carries no plasmid (Fig. 2A). The size of gene region of CI3125 is 4,329,969 bp, containing 4,306 CDS, 4,453 genes coding proteins, and 101 RNAs (83 tRNAs, 15 rRNAs, and 3 ncRNAs), and 43 pseudogenes (0.97%). The gene region accounts for 87.86% of the total genome length. The GC content in the gene region is 38.28%, while that of the intergenetic area is 29.36%.
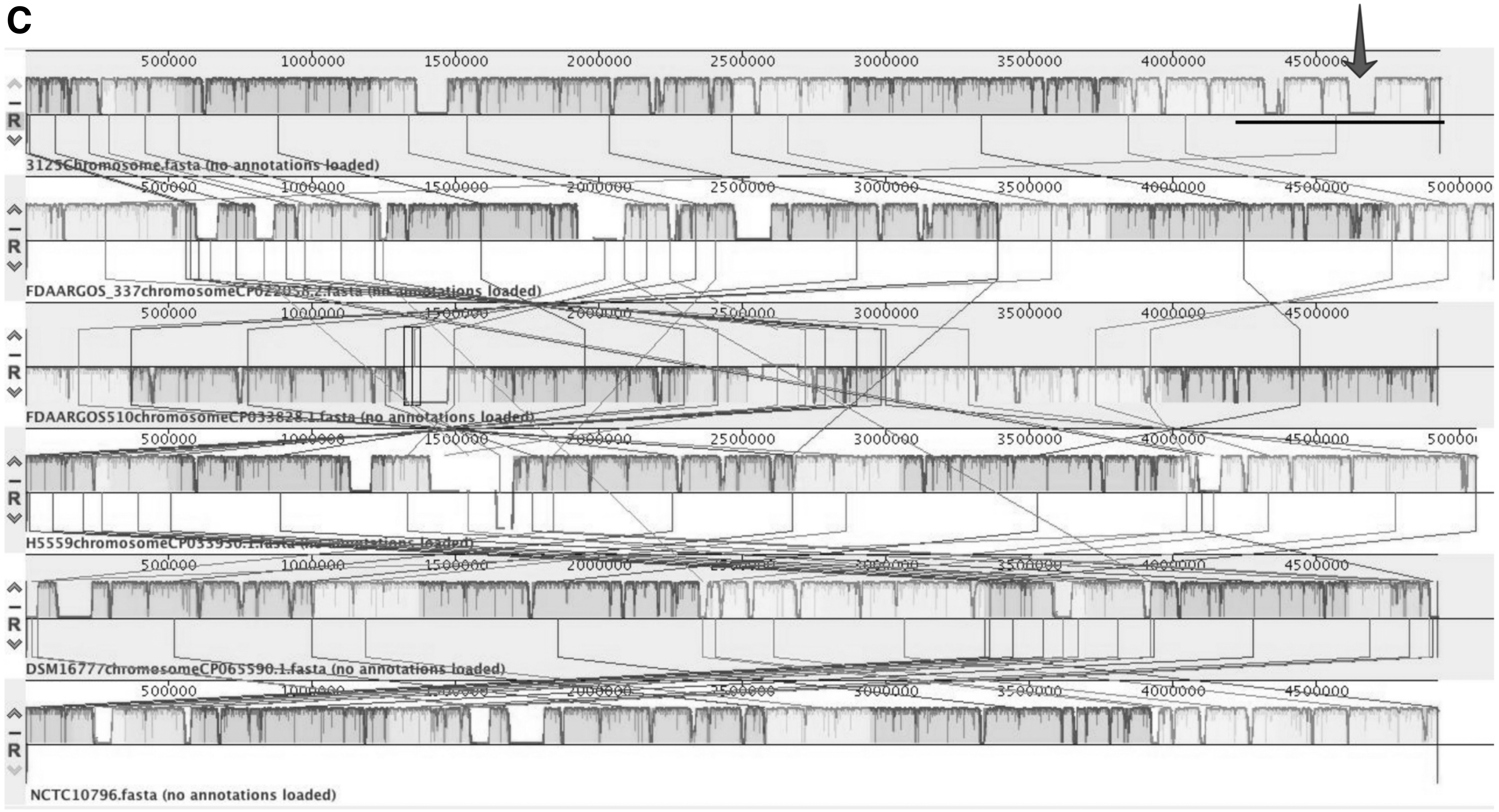
Genomic map of Chryseobacterium indologenes and its comparison with other bacteria
As for mobile elements, there are three incomplete phage regions in the genome of CI3125. In addition, we found three insertion sequences, two of which share the same sequence belonging to the ISL3 family. Moreover, there are six genetic islands (GIs) in CI3125 genome. The complete information on mobile elements is shown in Table 2. For better analysis of the homology of CI3125, we chose five closest C. indologenes strains (strain FDAARGOS 337, FDAARGOS 510, H5559, DSM16777, and NCTC10796) for whole-genome comparison with CI3125 based on the two types of phylogenetic trees described above (Fig. 2B). We found that a unique region only exists in CI3125 genome (Fig. 2C). The GC content of this region is relatively lower than that of the whole genome of CI3125, and we further found that this region contains GI06.
Mobile Elements Analysis of CI3125
GI, genetic island; IS, insertion sequence; PR, phast region.
Analysis of the GI 06
GI06 is a 99178-bp region that harbors 102 ORFs. The region can be characterized into three functional components: a 33,067-bp section comprising genes for the transport of heavy metals, a 38,947-bp sector consisting of conjugative transfer genes, and a 27,167-bp section containing genes for drug resistance. Located within the heavy metal transfer component are 30 ORFs that confer resistance to many heavy metals (including mercury, arsenic, zinc, iron, and copper), primarily occurring in Chryseobacterium and Sphingobacterium spp. Notably, two copies of RteC regulating the excision of conjugative transposon CTnDOT in Bacteroides spp. are found in this region (Fig. 3). The conjugative transfer component that harbors 45 ORFs predominantly comprised conjugative transfer genes.
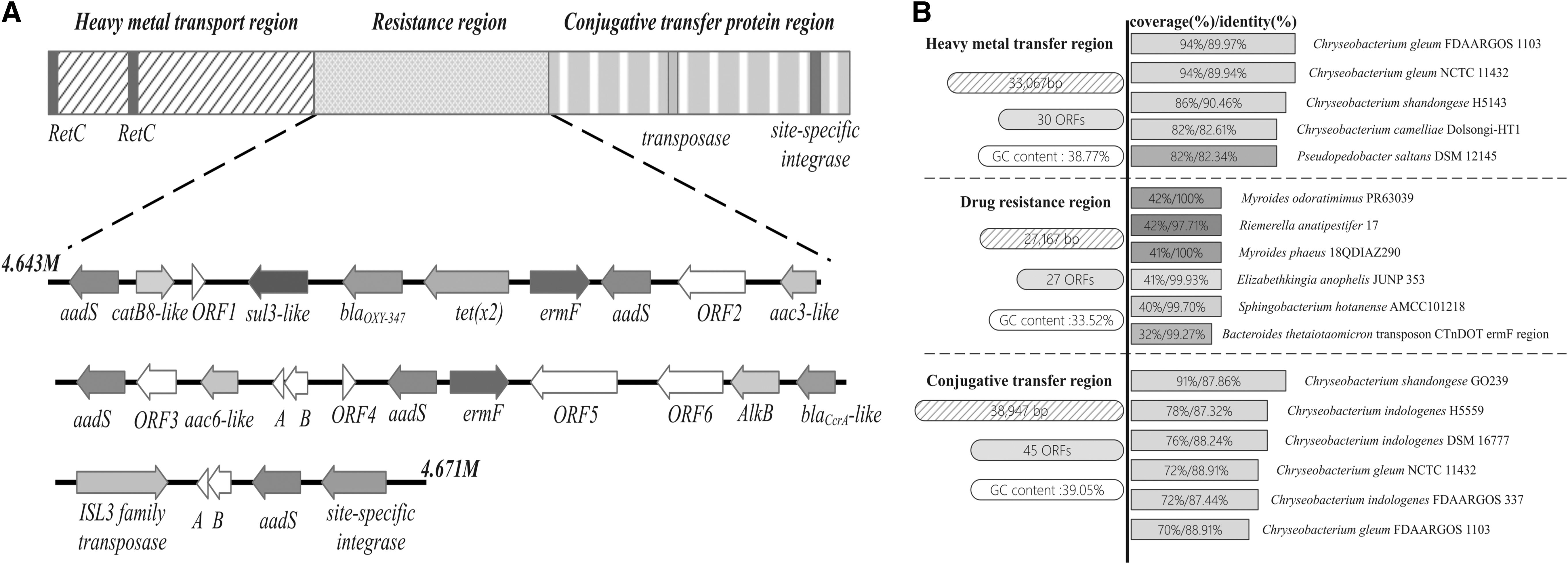
General features of GI06 in CI3125
Of significant note, the component corresponding to drug resistance consisting of 33.52% GC content (lower than the 37.2% GC content of the CI3125 genome) encompasses five copies of aadS, two copies of ermF, two β-lactase genes (blaOXA-347 and blaCcrA-like), catB6-like, tet(X2), sul3-like, and nine hypothetical ORFs. blaOXA-347 and ermF flank tet(X2), while two ORFs encoding one ISL3 family transposase and one site-specific integrase, are situated downstream of tet(X2) at 16,376 bp and 19,598 bp, respectively. Interestingly, complete mobile elements, such as transposons, integrons, and integrative and conjugative elements, adjacent to tet(X2), were not detected using BLASTN. Alignment of this region by BLASTN highlighted that all matches were below 50% in coverage. The best match (identity of 100% and coverage of 42%) was Myroides odoratimimus strain PR63039.
Upon restricting the coverage parameter to 30–100% in BLAST, 10 of the 20 matches (excluding uncultured bacteria) were isolated from Riemerella anatipestifer strains. Moreover, we observed that this region demonstrated high homology (coverage of 32% and identity of 99.27%) with the conjugative transposon CTnDOT ermF region.
Resistome of CI3125
The genome of CI3125 contains a plethora of antimicrobial resistant genes, including 7 perfect-matched, 6 strict-matched, and 220 loose-matched genes (Supplementary Fig. S2). Of note, CI3125 genome harbors various types of β-lactamase genes, including Ambler class A genes (blaCIA-4, blaIDC-like, blaACC-like, blaADC-like, and blaCMY-like), class B Metallo-β-lactamase (MBL) genes (blaIND2 and two blaCcrA-like), and class D gene (blaOXY-347). Intriguingly, the GC content of blaOXY-347 is 25.99%, significantly lower than that of other β-lactamase genes in CI3125, and its best hit organism is Bacteroides stercoris. Tetracycline resistance genes, such as tet(A)-like, tet(41)-like, tet(44)-like, rpsJ-like, tet(B)-like, tet(X)-like, and tet(X2), were also identified in CI3125 genome.
In addition, CI3125 genome contains genes conferring resistance to aminoglycosides (aadS, AAC(6)-like, and AAC(3)-like), chloramphenicol (catB6-like, catB8-like, and catB11-like), macrolides-lincosamides-streptogramins (ermF, macB-like, and rana-like), glycopeptide (van-like, bacA-like, and bcrA-like), fluoroquinolone (ermA-like and sul3-like), and tuberculous drugs (katG-like and tuf-like).
Despite a lack of information on the efflux pump of C. indologenes, we identified 12 genes that shared higher coverage and identity with known efflux pump genes (coverage between 99.5% and 50% and identity between 41.7% ≥ 35%) for analysis. Among the predicted efflux pumps, five belong to the RND family, three to the ABC family, and the remaining to the SMR family. The details of these resistance genes are catalogued in the Supplementary Table S1.
tet(X2) was not expressed in CI3125
The tet(X2)-expressing plasmid, pBR322-tet(X2), was constructed based on pBR322. The ampR ORF was precisely replaced by tet(X2), which was under the control of the ampR promoter, ensuring a constitutive and robust expression of tet(X2) (Supplementary Fig. S4). E. coli DH5ɑ, initially sensitive to tigecycline (MIC = 0.25 μg/mL), displayed a 16-fold increase in tigecycline resistance (MIC = 4 μg/mL) upon acquisition of pBR322-tet(X2), providing clear experimental validation of the functional capacity of tet(X2) ORF. The expression level of tet(X2) was assayed in CI3125 and DH5ɑ-tet(X2) after culturing with 2 μg/mL tigecycline. The results of quantitative reverse transcription PCR indicated that the expression level of tet(X2) in DH5ɑ-tet(X2) was ∼3,764-fold higher than that in CI3125 (Fig. 4).

The transcription level of tet(X2) in CI3125 and DH5ɑ-tet(X2).
Moreover, the deployment of a polyclonal antibody for tet(X2), prepared in this study, provided further corroboration, revealing expression of tet(X2) in E. coli DH5ɑ-tet(X2), but not in CI3125 (Fig. 5). An analysis of the promoter and ribosome-binding site (RBS) sequences of tet(X2) was carried out to elucidate the reason for its nonexpression in CI3125. The results showed that the sequence of tet(X2) ORF, plus 50 bp upstream of the initiation codon of 24 gene fragments, was highly conserved with only two types (T and G) of variations at the 12 bp site (Supplementary Fig. S3). We replaced the RBS of the ampR gene in pBR322-tet(X2) with the two types of RBS to examine if they were both complete and functional.

Western blot analysis for Tet(X2) detection. The molecular weight of Tet(X2) is 43 kDa. The sample of the lane “The truncked Tet(X2)” is the purified truncated Tet(X2) (33 kDa) by removing the initial 109 amino acid residues, and the sample volume is 10 μL (0.5 μg/mL); the sample volume of the lane “DH5ɑ-tet(X2)” is 10 μL; the sample volume of the lane “CI3125 S1” is 10 μL; the sample volume of the lane “CI3125 S2” is 20 μL; the sample volume of the lane “CI3125 S3” is 30 μL.
Remarkably, E. coli DH5ɑ, carrying the modified plasmids, showed the same level of tigecycline resistance as those carrying the original pBR322-tet(X2), indicating that the two types of RBS were functional. Although we identified a putative promoter located 16,381 bp upstream of tet(X2), the qRT-PCR assay indicated that it could not facilitate efficient transcription of tet(X2), for reasons unknown.
Efflux pumps play a significant role in the tigecycline resistance of CI3125.
We conducted a thorough investigation of the activity of tet(X2) at the cellular level in CI3125 by examining the degradation of tigecycline. To this end, CI3125 and DH5ɑ-tet(X4) were each incubated with tigecycline (8 μg/mL) for 8 hours. Strikingly, the concentration of tigecycline decreased by ∼50% when coincubated with DH5ɑ-tet(X4), while no significant concentration change was observed when incubated with CI3125, as reflected in our findings (Fig. 6). To further explore the role of efflux pumps in the resistance of tigecycline, we assessed the effects of five different efflux pump inhibitors on the growth of CI3125. Among these inhibitors, only CCCP demonstrated apparent toxicity toward the growth of CI3125, whereas the others had no discernible effects.
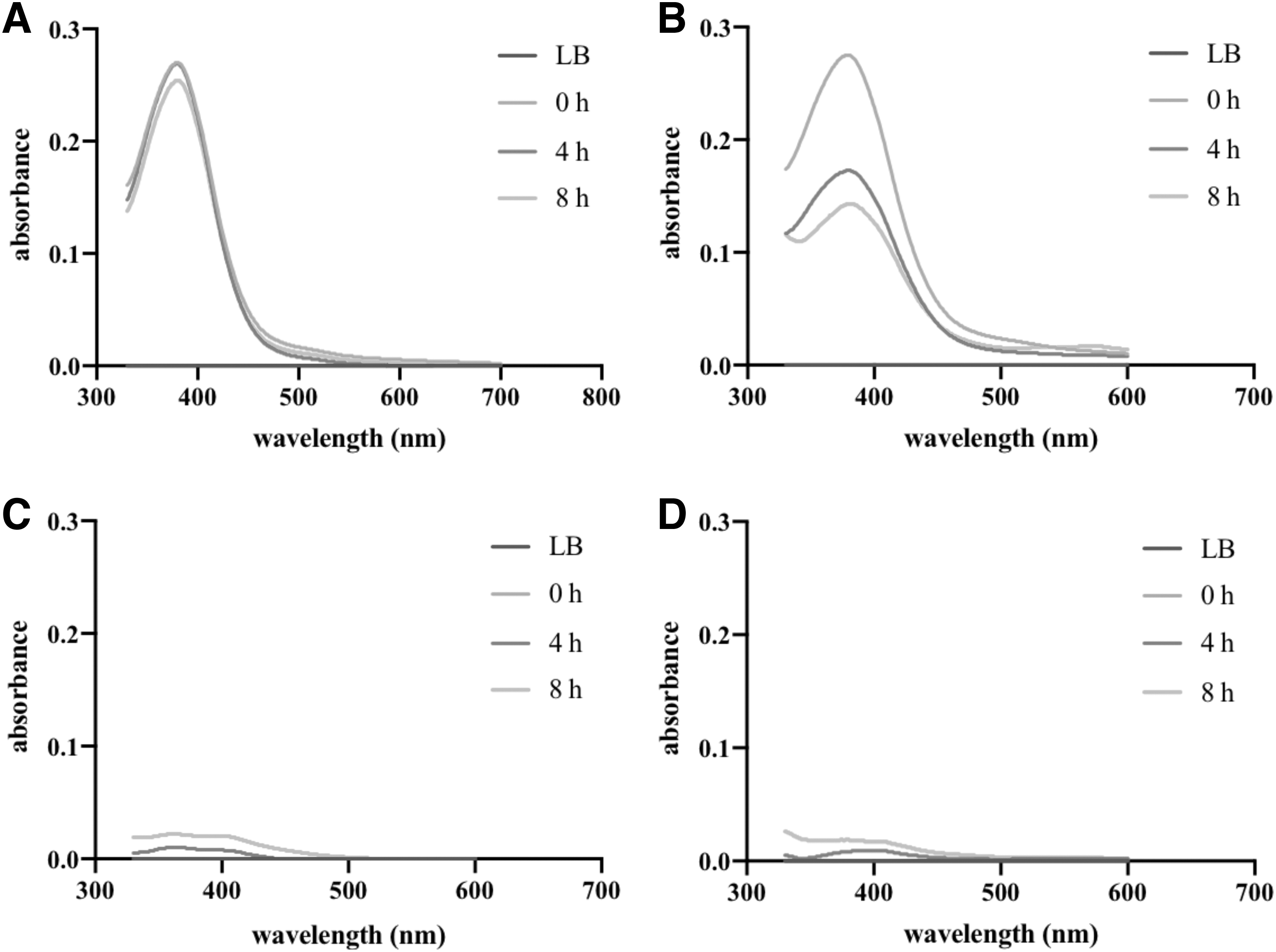
Based on the bactericidal effects of these inhibitors, we selected concentrations that had no apparent effect on the growth of CI3125 (25 μg/mL for NMP, 70 μg/mL for PAβN, 100 μg/mL for verapamil, and 50 μg/mL for reserpine) to test their MIC for CI3125. Remarkably, we noted two-fold, two-fold, four-fold, and eight-fold reductions of tigecycline MICs in CI3125 in the presence of NMP, reserpine, PAβN, and verapamil, respectively (Fig. 3 and Table 3). These results underscored the critical role of efflux pumps in conferring resistance to tigecycline in CI3215. Remarkably, of all the inhibitory agents, verapamil demonstrated the most robust inhibitory effect, leading us to speculate that the ABC transporter plays a pivotal role in conferring resistance to tigecycline.
The Minimum Inhibitory Concentration Value of Tigecycline in Efflux Pump Inhibition Assay
MIC, minimum inhibitory concentration; NMP, 1-(1-naphthyl-methyl)-piperazine; PAβN, phenyl-arginine-β-naphthylamide.
Discussion
Insufficient research has been conducted on the intricate mechanisms underlying the multidrug resistance exhibited by C. indologenes. In this study, we aimed to unravel the mechanisms of antimicrobial resistance in this species by using whole-genome sequencing and molecular biology experiments. Our results indicate that the drug-resistant region, housing tet(X2) within GI GI06, possessed a considerably lower GC content than that of the entire genome. Therefore, it appears plausible that this region represents an exogenous DNA fragment that was captured by CI3125. Remarkably, R. anatipestifer also harbors tet(X2) in a drug-resistant GI, thus revealing a noteworthy parallel with CI3125. 25 Besides, we identified other DNA fragments that were not originally assigned to C. indologenes. Although genomic sequences of C. indologenes, including CI3125, indicate the absence of plasmids, mobile elements such as GIs and ICE elements might facilitate gene exchange in this species. 26
Clinical isolates of Chryseobacterium spp. carrying tet(X2) present a relatively high rate of tigecycline resistance, which was directly associated the presence of tet(X) gene. 7 If tet(X2) is adequately expressed in CI3125, it can impart resistance to all members of the tetracycline family antibiotics. Nevertheless, despite previously reported C. indologenes strains carrying tet(X2) being susceptible to minocycline, our WB and qPCR experiments disclosed that tet(X2) was not expressed in CI3125. Thus, the inconsistent resistance to tigecycline and minocycline in CI3125 can be explained. We speculate that the weak expression of tet(X2) in CI3125 is attributable to the absence of a sufficiently effective promoter for its transcription. Moreover, we caution against relying on the detection of tet(X2) in Flavobacteriaceae as a means of verifying tigecycline resistance.
Efflux pump systems have been implicated in potentially mediating tigecycline resistance in bacteria.4,5 In evaluating the involvement of efflux pumps in drug resistance, efflux pump inhibitors can be utilized. For instance, verapamil has been demonstrated to inhibit several bacterial ABC efflux pumps in Mycobacterium tuberculosis isolates. 27 Our study showed that verapamil restored tigecycline susceptibility in CI3125, thus pointing toward the indispensable role of ABC efflux pumps in conferring tigecycline resistance in this species. Notably, RND efflux pumps have been identified to significantly impact tigecycline resistance in Klebsiella pneumoniae and Acinetobacter baumannii.24,25
PAβN, an inhibitor of RND family efflux pumps, is capable of significantly reducing tigecycline MIC values in CI3125, indicating the involvement of ABC efflux pumps in the bacterium's tigecycline resistance. However, the impact of NMP, another RND family efflux pump inhibitor, 28 on CI3125's tigecycline resistance was found to be insignificant. Similarly, Reserpine's potential to decrease MICs of nalidixic acid, ciprofloxacin, and norfloxacin in A. baumannii isolates has been reported previously, 29 but its impact on CI3125's tigecycline resistance was comparatively weak, akin to NMP. Thus, these results lead us to speculate that multiple efflux pumps are possibly involved in mediating tigecycline resistance in CI3125.
Conclusions
The carriage of a dormant tet(X2) gene renders CI3125 susceptible to the entirety of the tetracycline analog family except tigecycline, while concurrently exhibiting functional efflux pumps that endow the bacterium with resistance to the potent antimicrobial agent tigecycline. The apparent discordance in susceptibility toward tigecycline observed among various tetracycline family members may be attributed to a ubiquitous mechanism. Although the tet(X2) gene in CI3125 remains silent, its horizontal transfer via genetic elements such as genomic islands cannot be precluded.
Footnotes
Authors' Contributions
Y.L., M.C., Y.J., and W.W. contributed to methodology, software, collecting data, and to writing the original draft. H.W. and L.D. were involved in investigation, methodology, data curation, and validation, Z.Z. contributed to data curation and editing.
Data Availability
Ethics Approval
Ethics approval for this study was obtained from Shenzhen Children's Hospital Ethics Committee (201601304). Because the data are anonymous, the requirement for patient informed consent was waived.
Consent to Participate
All authors directly participated in the planning, execution, or analysis of the study.
Consent for Publication
The contents of this article have not been copyrighted or published previously and are not under consideration for publication elsewhere. All authors have read and approved the final version submitted.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Discipline Construction Project of Guangdong Medical University (Grant No. 2043K2022005) and Health and Family Planning Commission of Shenzhen Municipality (Grant No. SZGSP012).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.