
Abstract
Methicillin-resistant Staphylococcus aureus (MRSA) is a major infectious disease pathogen, and its molecular epidemiological profile has been changing. In this study, a total of 279 MRSA isolates were collected from patients with bloodstream infection (BSI) in Hokkaido, northern main island of Japan, for a 2-year period from August 2019 to July 2021. CC5 (ST5/ST764)-MRSA-IIa (SCCmec-IIa) (47%, n = 132) and CC1 (ST1/ST2725/ST2764)-MRSA-IVa (42%, n = 116) were found to be major lineages, with CC8-MRSA-IVa being lower prevalence (5%, n = 13). CC1-MRSA-IVa showed a relatively increased proportion compared with our previous study (22%, 2017–2019). Seven isolates with SCCmec IVa (2.5%) were positive for Panton–Valentine leukocidin genes on ΦSa2usa and belonged to ST8/spa-t008/agr-I/coa-IIIa, showing genetic features of the USA300 clone. Among these isolates, six isolates harbored arginine catabolic mobile element (ACME) type I typical to the USA300 clone, while it was not detected in an isolate (strain R3-8). Whole genomic analysis of strain R3-8 revealed that its chromosome was highly similar to the USA300 strain TCH1516, but lacked ACME, carrying a plasmid genetically close to that of USA300 strains. The present study revealed increasing trend of CC1-MRSA-IV and occurrence of a novel variant of the USA300 clone among MRSA from BSI in northern Japan.
Introduction
S
A major resistance determinant of MRSA is mecA, which encodes PBP2a with low affinity to β-lactam antimicrobials. mecA gene is carried on a cassette chromosome SCCmec, which is variable in size (∼20–68 kb) and genetically differentiated into at least 15 types (I through XV).5,6 Among them, SCCmec types IV and V are more commonly associated with infections in community, while types I through IV are involved in health care-associated infections.7,8 Although numerous MRSA clones have been described to date, major clones are spread globally through international transmission. Health care-associated MRSA (HA-MRSA) includes ST5-SCCmec II (ST5-MRSA-II; New Yolk/Japan clone), ST239-MRSA-III (Hungarian/Brazilian clone), ST22-MRSA-IV (EMRSA-15), and others, while more variable clones have been identified for community-acquired MRSA (CA-MRSA) such as ST1-MRSA-IV (USA400), ST8-MRSA-IV (USA300), ST30-MRSA-IV (Southwest Pacific clone), ST59-MRSA-V (Taiwan clone), ST80-MRSA-IV (European clone), ST772-MRSA-IV (Bengal Bay clone), etc.7–10
It is notable that CA-MRSA is considered more virulent and transmissible than HA-MRSA, 11 by which current epidemiological status of MRSA has been changing worldwide. In recent epidemiological studies of S. aureus from BSI, the increasing trend of specific MRSA clones over the past few decades is observed in Asia,12–16 Australia, 17 Europe,18,19 and the United States, 20 although types of the clones are different depending on country.
Among the CA-MRSA clone, USA300 is a representative international/pandemic clone that has been predominating the United States, 21 and shows global spread 22 with an upward trend among BSI isolates in Taiwan and Spain.14,15,18 USA300 belongs to ST8/spa-t008 with SCCmec IVa, and characteristically harbors in its chromosome a bacteriophage (ΦSA2usa) genome that contains Panton–Valentine leukocidin (PVL)-encoding genes (lukS-PV-lukF-PV) and arginine catabolic mobile element (ACME). 23 PVL is a bicomponent pore-forming toxin targeting neutrophil, and is primarily linked with skin and soft tissue disease and also severe invasive disease, including bacteremia.24,25 ACME is a genome cassette located adjacent to SCCmec and contains three genetic components in a complete form (type I) (arc cluster, opp cluster, speG) that are related to increased ability to colonize human skin. 23
In Japan, spread of clonal complex (CC) eight PVL-positive MRSA from skin and pus over the last decade was documented by nationwide surveillance, 26 and slight increased prevalence of USA300 clone was described among general clinical isolates in Tokyo. 27 Regarding MRSA from BSI, the predominance of ST8-SCCmec IV MRSA was reported in a cross-sectional study, 28 and relative increase in SCCmec IV MRSA replacing previously dominant SCCmec II MRSA was observed in a single medical institution. 29 A nationwide surveillance of MRSA from BSI showed the dominance of ST8-MRSA-IV and ST1-MRSA-IV, while describing regional difference among major types of MRSA. 30 However, the trend of MRSA clones from BSI in Japan has not yet been clearly understood due to dearth of molecular epidemiological study for a long period of time. We previously reported the molecular epidemiological features of HA-MRSA and CA-MRSA in northern Japan, and described the dominance of CC5-MRSA-II strains and low prevalence of ST8-MRSA-IV.31,32
Furthermore, in the study of MRSA from BSI during the period 2017–2019, the highest rate (5.1%) by that time of USA300 clone among MRSA was described, with detection of novel isolates (ACME-positive ST764-MRSA-II and ACME-negative ST1-MRSA-IV). 33 The present study was conducted to monitor clonal diversity and their prevalence of MRSA from BSI, for 2-year period following the previous study under the same study design in northern Japan. Comparison with our previous findings revealed evident change in the prevalence of MRSA clones and emergence of new variant presumably derived from the USA300 clone.
Materials and Methods
Bacterial isolates
During a 2-year period between August 2019 and July 2021, S. aureus was isolated from blood specimens that were submitted to Sapporo Clinical Laboratory, Inc. from hospitals and clinics in Hokkaido prefecture, Japan, for routine bacterial examination. The blood culture bottles used were BD BACTEC™ 23F (aerobic. medium) or 22F (anaerobic medium). Isolates grown in culture bottles were inoculated onto blood agar plates and incubated at 37°C for 24 hr aerobically, and species identification was performed by matrix-assisted laser desorption ionization (MALDI)–time of flight mass spectrometry using MALDI Biotyper (BRUKER). Initially, MRSA was identified phenotypically by measuring minimal inhibitory concentrations (MICs) of oxacillin (OXA) and cefoxitin (FOX). Individual isolates were stored in Microbank (Pro-Lab Diagnostics) at −80°C, and were recovered when they were analyzed. The presence of mecA in these isolates was confirmed by multiplex PCR as described below. Only one isolate per patient was included in this study.
Antimicrobial susceptibility testing
Antimicrobial susceptibility was measured by broth microdilution test using Dry Plate Eiken DP42 (Eiken Chemical, Tokyo, Japan). MICs within limited ranges were measured for 21 antimicrobials; OXA, FOX, penicillin (PEN), ampicillin, ampicillin–sulbactam, cefazolin, cefepime, imipenem, gentamicin (GEN), arbekacin (ABK), erythromycin (ERY), clindamycin (CLI), vancomycin (VAN), teicoplanin (TEC), daptomycin (DAP), linezolid (LZD), minocycline (MIN), fosfomycin (FOF), rifampicin, levofloxacin (LVX), and trimethoprim/sulfamethoxazole. Resistance was judged according to Clinical Laboratory Standards Institute (CLSI) breakpoints for broth microdilution. 34 For antimicrobials whose breakpoints are not defined by CLSI guidelines, we employed the European Committee on Antimicrobial Susceptibility Testing breakpoint for FOF (32 μg/mL, Staphylococcus spp.), 35 and a unique breakpoint for ABK (4 μg/mL, which is higher than the 2 μg/mL defined by the Japanese Society of Chemotherapy for respiratory infection). 36
Genotyping and genetic characterization
The presence of staphylococcal 16S rRNA gene, nuc, mecA, lukS-PV-lukF-PV, and ACME-arcA, was examined for all the isolates by multiplex PCR assay according to the previously described protocol 37 with some modifications. In this multiplex PCR, primers for detection of mecA was replaced with MecA2 (5′-AGTTCTGCAGTACCGGATTTGC-3′) and MecA147-F (5′-GTGAAGATATACCAAGTGATT-3′) that amplify 343-bp product. In the PCR reaction mixture, a concentration of 0.22 pmol/μL was used for 16S rRNA primers (Staph756F and Staph756R), whereas 0.44 pmol/μL for all other primers. In addition, PCR targeting the nonribosomal peptide synthetase (NRPS) gene with the primers nrps-F and nrps-R was performed as described previously for all the isolates to discriminate non-S. aureus SAC species (e.g., Staphylococcus argenteus) from S. aureus. 38 SCCmec type and subtype of SCCmec IV and SCCmec II were determined by multiplex PCR,39,40 and PVL phage was typed by PCR assay as described previously. 41 For all the ACME arcA-positive strains, ACME types I, II, and II′ were classified by long-range-PCR as described previously. 31
Staphylocoagulase genotype (coa type), spa type, and agr type were determined as described in previous studies.33,42,43 Sequence type (ST) was determined by multilocus sequence typing (MLST) scheme 44 and assigned to CC, according to the PubMLST website. The presence of genes encoding the following virulence factors were analyzed by multiplex or uniplex PCR as described previously32,45,46: staphylococcal enterotoxin (-like) proteins (sea-see, seg-selu, selx, sey, selw, selz, sel26, and sel27), TSST-1 (tst-1), exfoliative toxins (eta, etb, and etd), leukocidins (lukDE and lukM), hemolysins (hla, hlb, hld, and hlg), adhesins (eno, cna, sdrC, sdrD, sdrE, fib, clfA, clfB, fnbA, fnbB, icaA, icaD, ednA, ednB, bap, and vWbp), and modulators of host defense (sak, chp, and scn).
Genetic determinants conferring resistance to PEN (blaZ), macrolides [erm(A), erm(B), erm(C), and msrA], aminoglycoside [aac(6′)-Im, aac(6′)-Ie-aph(2″)-Ia, ant(3″)-Ia, ant(4′)-Ia, ant(6)-Ia, ant(9)-Ia, ant(9)-Ib, aph(2″)-Ib, aph(2″)-Ic, aph (2″)-Id, and aph(3′)-IIIa], oxazolidinone and phenicol resistance gene (optrA, fexA, and cfr), and those related to methicillin resistance (femA/femB) were detected by uniplex or multiplex PCR using the primers previously reported.32,45–47
Detection of speG was performed for all the isolates by PCR as described previously.31,33
Whole-genome sequencing analysis of an ACME-negative USA300-related strain R3-8
For the whole-genome analysis of an ACME-negative, ST8-MRSA-IVa (strain R3-8), genomic DNA was extracted using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany), and purified with AMPure XP (Beckman Coulter) followed by removal of low-molecular-weight fragments with the Short Read Eliminator XS Kit (Circulomics, MD). The sequencing library was prepared with SMRTBell Express Template Prep Kit 2.0 (PacBio, CA) and long-read sequencing was performed by using the PacBio Sequel IIe system (PacBio). The assembled contigs of chromosome and plasmid were annotated by the National Center for Biotechnology Information (NCBI) Prokaryotic Genome Annotation Pipeline (https://www.ncbi.nlm.nih.gov/genome/annotation_prok). ResFinder, PlasmidFinder, VirulenceFinder, and SCCmec Finder databases from the Center for Genomic Epidemiology (https://bitbucket.org/genomicepidemiology) were used for the identification of resistance genes, replicons, virulence determinants, and the SCCmec type, respectively.
A threshold of 98% was used for identity and 60% for the minimum length for ResFinder and PlasmidFinder.
Data availability
The chromosome and plasmid sequences of strain R3-8 are deposited in GenBank of the NCBI and available as the accession numbers CP103860 and CP103861, respectively. Information of the sequences was made publicly available through the Sequence Read Archive in NCBI (Bioproject PRJNA874487).
Statistical analysis
Antimicrobial resistance (AMR) rates between different genetic groups of MRSA, and proportion of coa-VII (CC1) or coa-II (CC5) compared with those in the previous study 33 were statistically analyzed by Fisher's exact test using the js-STAR XR ver.1.1.9 software. A p-value <0.01 was considered statistically significant.
Results
Genotypes of MRSA isolates, prevalence of PVL and ACME
During the study period, a total of 279 MRSA isolates were obtained from blood culture. Table 1 summarizes frequencies of SCCmec types, coa type, and ST, and incidence of lukS/F-PV and ACME-arcA. Both SCCmec type IIa and IVa were dominant and showed almost the same frequency (47% and 46%; 132 and 129 isolates, respectively), while the remaining were other SCCmec IV subtypes (n = 14) and SCCmec V (n = 4). All the isolates were differentiated into 6 coa types and 12 STs containing 3 major CCs. CC5 (ST5/ST764)-coa IIa was the most common genotype (47%, n = 132) and found only in SCCmec IIa, with ST5 and ST764 showing similar frequency (n = 64 and 68, respectively).
Frequency of SCCmec type, Coagulase Genotype (coa), and Sequence Type/Clonal Complex Among Methicillin-Resistant Staphylococcus aureus Blood Isolates (n = 279) (August 2019 to July 2021) and Incidence of Panton–Valentine Leukocidin Genes and Arginine Catabolic Mobile Element
speG-positive isolates are included; all the six PVL+/ACME+ isolates (SCCmec IVa/coa-IIIa/ST8), two isolates with ACME II′ (SCCmec-IIa/coa-IIa/ST5, SCCmec-IIa/coa-IIa/ST764).
Strain R3-8 having USA300 clone-like genotypes (ST8/t008, SCCmec-IVa/coa-IIIa).
ACME, arginine catabolic mobile element; CC, clonal complex; PVL, Panton–Valentine leukocidin; ST, sequence type.
Second most common type was CC1-coa VIIa with SCCmec IVa (42%, n = 116). CC1 of this group comprised a predominant ST1 (n = 104), and minor types ST2725 (n = 7) and ST2764 (n = 5). Third common type was CC8-coa IIIa (9%, n = 24) with CC8, including ST8 (n = 23) and ST5990 (n = 1), and having SCCmec IV of three different subtypes (IVa, IVh, and IVl). SCCmec V MRSA was genotyped as ST30-coa IVa or ST45-coa VIIb. Remaining isolates had SCCmec IV and belonged to CC1 (ST81, ST188) or CC8 (ST72).
PVL genes were detected in seven isolates (2.5%, 7/279), which had SCCmec IVa and belonged to ST8, coa IIIa, and spa-t008, and their PVL phage was identified as ΦSa2usa (Tables 1 and 2). Six isolates among these ST8 MRSA harbored type I ACME, although one isolate was negative for ACME. Type II′ ACME was identified in PVL-negative isolates of ST5 (n = 1) and ST764 (n = 8) isolates. speG was detected in all the isolates with ACME-I and two isolates with ACME-II′ (Supplementary Tables S1 and S2).
Antimicrobial Resistance Profile and Resistance Determinants of Panton–Valentine Leukocidin and/or Arginine Catabolic Mobile Element-Positive Isolates (n = 16) and Selected Panton–Valentine Leukocidin/Arginine Catabolic Mobile Element-Negative Isolates (n = 78)
The following genes were undetectable in any strain: erm(T), erm(Y), aac(6′)-Im, ant(9)-Ib, ant(3″)-Ia, aph(2″)-Ib, aph(2″)-Ic, aph(2″)-Id, optrA, cfr, and fexA. femA/femB (factor essential for expression of methicillin resistance) and blaZ were detected in all the strains.
Novel ST identified in this study.
CLI-c, constitutive resistance to clindamycin; CLI-i, inducible resistance to clindamycin; ERY, erythromycin; FOF, fosfomycin; GEN, gentamicin; LVX, levofloxacin; MIN, minocycline.
Antimicrobial susceptibility
Most of CC5 MRSA isolates were resistant to ERY and CLI-c (constitutive) (>97%), and showed higher resistance rates to GEN, MIN, ERY, CLI-c, FOF than CC1, and CC8 isolates, as well as isolates of other STs (Table 3). In contrast, significantly higher rates of inducible resistance to CLI (CLI-i) were observed in CC1 and CC8 isolates than CC5 isolates. More than 96% of isolates were resistant to LVX, while all the isolates were susceptible to ABK, DAP, LZD, TEC, and VAN.
Antimicrobial Resistance Profile in Different Clonal Complex/Sequence Type of Methicillin-Resistant Staphylococcus aureus Isolates from Bloodstream Infection
None of the isolates showed resistance to ABK, DAP, LZD, SXT, TEC, and VAN.
Significant difference compared with CC5 (ap < 0.01; bp < 0.05), or CC1 and CC5 (cp < 0.01).
ABK, arbekacin; DAP, daptomycin; LZD, linezolid; SXT, trimethoprim/sulfamethoxazole; TEC, teicoplanin; VAN, vancomycin.
AMR genes and virulence factors
Further genetic analysis (spa typing, agr typing, AMR genes, virulence factors) was performed for a total of 94 representative isolates comprising all the PVL- or ACME-positive isolates (n = 16) and 78 PVL/ACME-negative isolates (Table 2 and Supplementary Tables S1 and S2). Seventy-eight PVL/ACME-negative isolates were selected from different CC/ST, SCCmec types, and from those isolated in different dates (year/month). All the seven PVL-positive ST8 were assigned to spa type t008, agr group I, and mostly harbored sek, seq, and immune evasion cluster (IEC) type B (sak, chp, scn). CC5 (ST5, ST764) isolates were classified into agr-II and various spa types, with t002 being the most common, and mostly harbored resistance genes to macrolides and aminoglycosides. The enterotoxin gene cluster (seg-sei-sem-sen-seo) was more commonly detected in ST764. CC1 (ST1, ST2725, ST2764) with SCCmec IVa (MRSA-IVa) isolates belonged to spa-t1784/agr-III, and had erm(A) at high rate, with low prevalence of aminoglycoside-modifying enzyme gene.
Among these CC1 isolates, sea, sek, and seq were detected in 32% (11/34), and collagen-binding protein gene (cna) was highly prevalent, although it was not detected in CC5 and CC8 isolates. TSST-1 gene (tst-1) was detected in a few CC5 isolates and all the four ST8-SCCmec IVl isolates harboring spj specific to this SCCmec IV subtype. Although IEC type B was generally found in CC5 and CC8 isolates, IEC type E (sak, scn) was most frequent in CC1 MRSA and also found in some isolates of CC5 and CC8.
Change in incidence of main MRSA clonal group over time
To presume the trend of major MRSA clones from BSI in northern Japan, proportions of CC1-MRSA-IV, CC5-MRSA-II, and CC8-MRSA-IV in the present study were compared with those in our previous study (Table 4). 33 Proportion of CC1-MRSA-IV in 2019–2021 (41.6%; 116/279) was significantly higher than that in 2017–2019 (22.0%; 61/277), while proportion of CC5-MRSA-IIa in the present study was lower than that in the previous study. Rates of CC8-MRSA-IV and the USA300 clone (ST8-MRSA-IVa, PVL-positive) among all the MRSA in 2019–2021 (8.6% and 2.5%, respectively) were slightly lower than those in 2017–2019.
Proportion of CC5, CC1, CC8, and USA300 Clone Among Methicillin-Resistant Staphylococcus aureus Isolates from Bloodstream Infection in Different Periods in Northern Japan
Significant difference (p < 0.01) compared with those in August 2017 to July 2019.
MRSA, methicillin-resistant Staphylococcus aureus.
Whole genomic analysis of ACME-negative ST8-MRSA-IVa
In the present study, we identified seven ST8-MRSA-IVa isolates belonging to spa-t008/agr-I/coa-IIIa and harboring lukS-PV-lukF-PV in ΦSa2usa phage, which are typical traits of the USA300 clone. 23 However, only an isolate designated strain R3-8 lacked ACME, whereas other isolates had type I ACME as known for the USA300 clone. To understand the genomic structure of R3-8 and its genetic relationship to USA300 clone, whole genomic sequence was determined.
R3-8 had a circular chromosome of 2,918,241 bp, and a plasmid of 27,068 bp. The pairwise dDDH value with R3-8 genome was analyzed by using Type Strain Genome Server. R3-8 chromosome had the highest dDDH value (d0) of 99.5% with USA300 strain FPR3757 (CP000255) and USA300 strain TCH1516 (NC010079.1), whereas lower values to MRSA strains MW2 (NC003923.1) (97.2%) and N315 (BA000018). Subsequently, genomic organization of strain R3-8 was compared with USA300 strains FPR3757 and TCH1516 (Fig. 1). Although strains FPR3757 and TCH1516 show 100% dDDH value, there is a difference in genomic organization, that is, position of genetic element ICE6013 (13.4 kb).48,49
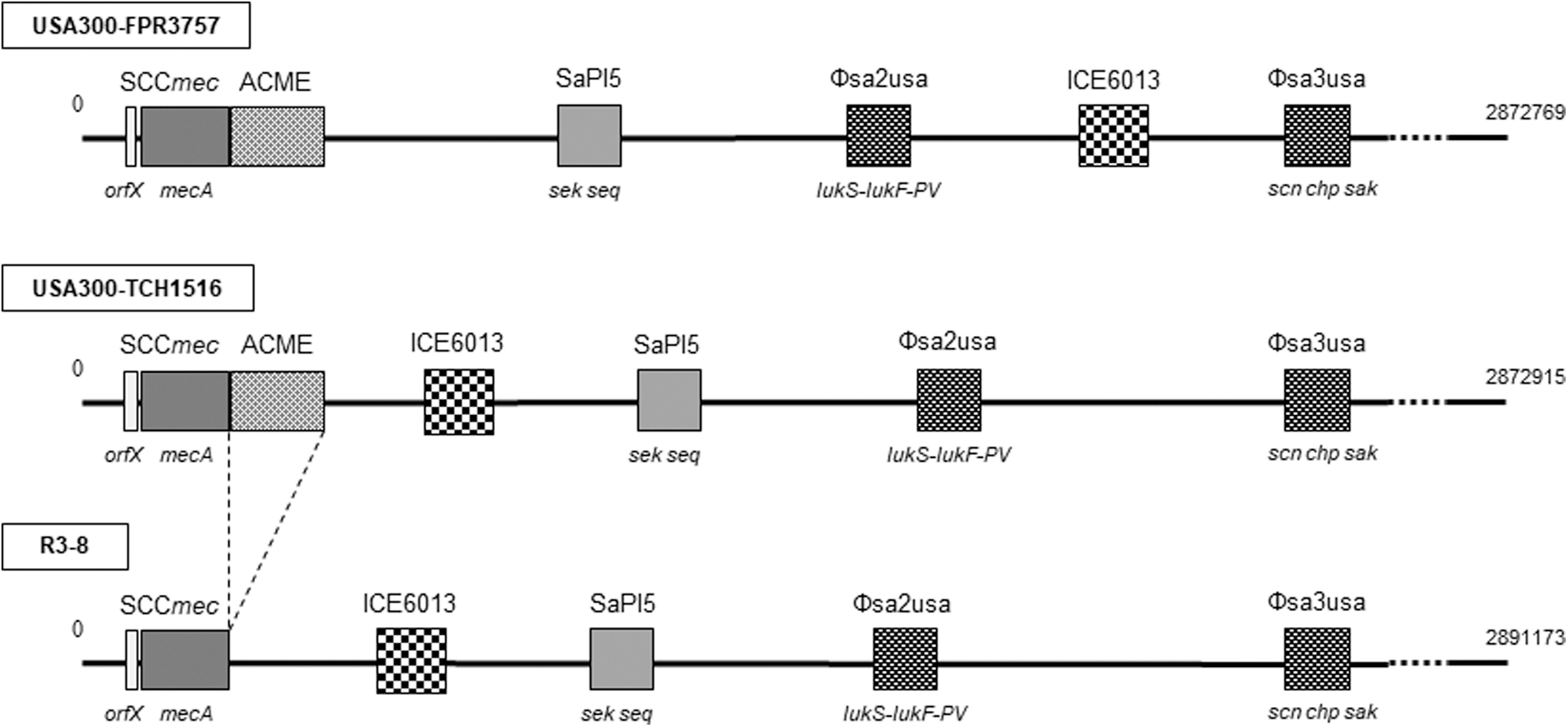
Schematic representation of genomic structures of the USA300 strains FPR3757, TCH1516, and the USA300-like strain R3-8 (GenBank Accession Nos. CP000255, NC_010079, CP103860, respectively). Boxes show representative genomic components (cassette chromosome, pathogenicity island, phage, and ICE6013), and their sizes in the figure are not proportionally expressed as compared with that of whole genome. ICE, integrative and conjugative element; SaPI, Staphylococcus aureus pathogenicity island.
Strain R3-8 chromosome showed the identical genomic organization to that of strain TCH1516, having the same positions and orientation of ICE6013, SaPI5, ΦSa2usa, and ΦSa3usa. Only a distinct finding was that strain R3-8 lacked ACME, which is located adjacent to SCCmec of strains FPR3757 and TCH1516. In addition to virulence factors detected by PCR (Supplementary Table S2), analysis by VirulenceFinder revealed the presence of some genes encoding exoenzymes (aureolysin, serine proteases) in the strain R3-8 (Supplementary Table S3). A plasmid associated with strain R3-8 was revealed to have high identity (99–100%) to many plasmids of S. aureus strains reported in the United States, including strain ER02069.3 (GenBank Accession No. CP030667), USA300-SUR19 (CP014431.1), and also TCH1516 (CP000731.1), by BLAST search. This plasmid contained AMR genes blaZ, cadD, msr(A), and aph(3′)-IIIa.
Discussion
In the present study on BSI isolates of MRSA in 2019–2021, it was notable that the proportion of CC1-MRSA-IV (mostly ST1-spa t1784) (42%) was comparable to that of CC5-MRSA-II (47%), being significantly higher than that observed in 2017–2019 (22%). In Japan, similar trend of MRSA clones has been reported in recent epidemiological studies. In a nationwide surveillance of BSI in Japan in 2019, ST1-MRSA-IV was found as a dominant type (29.6%) as well as ST8-MRSA-IV (30.7%), although difference in prevalence of MRSA types was observed depending on region. 30 In a study for blood and soft tissue isolates in Tokyo and its surrounding area in 2018–2019, SCCmec IV was the dominant type among CA-MRSA (73%), of which almost half belonged to CC1 (major type ST1 and minor types ST573, ST2725, ST2764, and ST4143) with major spa type t1784. 50 Similarly, in our present and previous study for BSI isolates, 33 among CC1-MRSA-IVa, the most common ST1 and infrequent ST2725 and ST2764, and single dominant spa t1784 were identified. Nationwide study on BSI isolates also reported the dominance of spa t1784 among ST1-MRSA-IV. 30
Therefore, CC1-MRSA-IV, represented by ST1-spa t1784, has been spreading in Japan, among BSI and also other infection types in the past decade. This ST1-MRSA-IV/spa t1784 lacks PVL gene, unlike the early CA-MRSA clone USA400 (prototype strain MW2). 8 Such PVL-negative ST1-MRSA-IV was previously described in Australia as WA-MRSA-151 and also in some European countries.8,52 Although genomic characterization was reported, any distinct virulence factor was not specified. 50 Thus, further study is necessary to define virulence determinants and biological characteristics to confer putative advantage to grow and spread for this MRSA clone.
Although the reason for the decline of CC5-MRSA-II, in exchange for the rise of CC1-MRSA-IV in Japan is not evident, it may also be associated with a gradually decreasing trend of MRSA since 2000. According to the national AMR surveillance (JANIS), the rate of MRSA among S. aureus from inpatient shown as 69% in 2000 decreased to 51.1% in 2013, and 46% in 2021. 53 This trend is considered to be accelerated by the National Action Plan on AMR formulated in 2016, in response to the Global Action Plan released by WHO in 2015. 53 Actually, antimicrobial use, especially oral antimicrobials as well as β-lactams, has been decreasing after 2016, with remarkable drop in 2020. 53 Health care-associated ST5-MRSA-II colloquially referred to as New York/Japan clone,7,10 has been predominant in Japan and shows multidrug resistance as found in the present study as well as previous studies.30–33 Thus, the declining of MRSA rate in association with control of antimicrobial usage might lead to reduction of multidrug-resistant clone such as ST5-MRSA-II.
Alternatively, less-resistant MRSA clone represented by CC1-MRSA-IV might have relatively become obvious and increased among S. aureus causing infections in humans. This change is suggested to occur in parallel with global spread of CA-MRSA in the community, which frequently carries SCCmec IV.8–10 Similarly, chronological decrease in ST5 MRSA was observed in other countries, for example, South Korea 12 and Spain. 18 Nevertheless, it was revealed in recent studies in Czech Republic 54 and Slovakia 55 that CC5-MRSA-II was dominant and belonged to multiple spa types, including a presumptive emerging one. Although the trend of CC5 MRSA in these countries is not evident, the findings suggest that this MRSA clonal group has been still persisting locally associated with genetic evolution, posing a need for further attention.
Global spread of PVL-positive ST8-MRSA-IV (USA300 clone) has been concerned in the past decade. 22 In Japan, first presumptive autochthonous case of infection with USA300 MRSA clone occurred in 2008. 56 Thereafter, this clone has been sporadically reported, and slightly increasing trend of prevalence is observed among various clinical isolates.26,27 In the nationwide study on BSI isolates in 2019, 30 ST8-MRSA-IV accounted for 30.7%, among which 6% of isolates had PVL genes and ACME, representing the USA300 clone. Overall detection rate of the USA300 clone in this study (1.8% of all isolates, 5/270) may be comparable to that observed in our present study (2.5%; 7/279), although the prevalence of ST8-MRSA-IV among all the BSI isolates was considerably lower (8.6%; 24/279). The prevalence of the USA300 clone as well as CC8-MRSA-IV among BSI isolates in our study (2019–2021) was slightly lower than that in 2017–2019 without significant difference. This may imply that the USA300 clone in BSI is persisting at a low level in northern Japan.
A remarkable finding was identification of the presumptive variant of ST8-USA300 clone (ΨUSA300), strain R3-8, which lacks ACME. Whole genomic analysis revealed that it is closer to USA300 strain TCH1516 than FPR3757 with respect to the position of ICE6013 in the chromosome, and having a plasmid that is similar to that of TCH1516. Accordingly, strain R3-8 is presumed to be derived from strain TCH1516, a thorough genomic alteration to lose ACME. USA300 FPR3757 was isolated from abscess of an HIV-positive adult patient representing a highly resistant strain, while TCH1516 (USA300-HOU-MR) was derived from an adolescent patient with sepsis and purely acquired in the community in the United States.23,49 These strains are genetically highly similar and thus suggested that their subtle genomic difference is implicated in their pathogenesis. 49 Therefore, further molecular epidemiological study to identify different strains among USA300 clone may be of value to presume the virulence and probability to evolve into a new variant, such as strain R3-8.
Recently in Japan, some distinct variants of the USA300 clone (ΨUSA300) have been identified, including the strain having an altered ccrB2, 57 strains with lineage-specific mutations, 58 and a Latin American variant (USA300-LV/J), which carries the copper and mercury resistance mobile element instead of the ACME in southern Japan. 59 Furthermore, in northern Japan, PVL-positive MRSA-IVa belonging to ST6562, a single-locus variant of ST8, with or without ACME was identified in severe pediatric infection, 60 and also in skin/oral cavity of healthy adults.61,62 The presence of such distinct ΨUSA300 strains may suggest that the USA300 clones have been uniquely evolving in Japan, being adapted to the community.
The present study revealed increased prevalence of CC1-MRSA-IV MRSA and occurrence of a novel variant of the USA300 clone among BSI isolates in northern Japan, whereas CC8-MRSA-IV appears to be persisting at a low level. Further epidemiological study may be necessary to monitor the trend of CC1 and CC8 MRSA, and to clarify the diversity and prevalence of USA300 variant and their pathogenic significance.
Ethics Statement
In this study, we used isolates routinely cultured from clinical specimens from hospitals and clinics, and no human participants were directly involved. Hence, clearance of human ethics was not required.
Footnotes
Authors' Contributions
Conceptualization: M.S.A., and N.K.; methodology: M.S.A., N.U., and M.K.; formal analysis: M.S.A., N.O., M.H., and N.K; data curation: M.S.A. and N.K.; resources: Y.K., K.K., and M.I.; writing—original draft preparation: M.S.A; writing—review and editing: M.S.A. and N.K.; supervision and project administration: N.K.; funding acquisition: M.S.A., N.U, and NK.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by JSPS (Japan Society for the Promotion of Science) KAKENHI grant number JP20H03933, JP21K10401, and JP21K10417.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.