
Abstract
This study depicts the drug-resistance and phylogenomic characteristics of 365 Escherichia coli (EC) and 76 Klebsiella pneumoniae (KP) isolated from stray dogs (293) in and around Kolkata, India. Initial screening found 59 isolates, including 48 E. coli and 11 KP multidrug resistant, which included 33 extended-spectrum β-lactamase, 41 AmpC β-lactamase and 18 metallo-β-lactamase producers carrying blaNDM-1 (11) and blaNDM-5 (7) genes. Majority of them had the resistant genes such as blaCTX-M (33), blaTEM (18), blaSHV (4), blaOXA (17), blaFOX (2), blaDHA (2), blaCITM (15), blaCMY-2 (13), blaGES (2) and blaVEB (2), qnrS (15), qnrB (3), aac-6′-Ib-cr (14), tetA (26), tetB (14), sul-1 (25), armA (2) and rmtB (6), in addition to adherence genes such as csgA (33), fimA (27), fliC (13), sdiA (33), rcsA (38), and rpoS (39). They also carried plasmid of diverse replicon types of which IncFIA and FIB were the most frequent. Phylogrouping categorized most of the MDR E. coli in phylogroup A (20), B1 (14), and B2 (6). Enterobacteriaceae repetitive intergenic consensus-polymerase chain reaction (ERIC-PCR) showed genetic diversity of multidrug resistant isolates irrespective of their origin, resistance, and virulence types, differentiating the EC in five clades (A–E) and KP in four clades (A–D). As these stray dogs, which had no history or scope of previous antimicrobial therapy, were found to have contracted potential antimicrobial resistance pathogens, the role of environment in spread of such pathogens and further possibility of human infections cannot be ruled out.
Introduction
Emergence of antimicrobial resistance (AMR) is possibly the biggest threat ever faced by the human civilization. It is well known that the overuse/misuse of antibiotics in human and veterinary treatment, as well as in food production, has aided the growth of multidrug-resistant bacteria. 1 Multidrug-resistant (MDR) Enterobacteriaceae that produce extended-spectrum β-lactamase (ESBL) and carbapenemases have been known for increased mortality among the hospitalized patients; however, they are increasingly reported in community infections and environment. 2 This spread of the bugs beyond hospitals and farms may even affect the sectors with low antimicrobial usage.
Recent study suggested that the backyard goatery, which has negligible antibiotic exposure, could also harbor AMR pathogens. 3 Of late, we have detected quite a few numbers of carbapenem-resistant strains in community pond water indicating their reach beyond health care premises. 4 Thus, environmental spread could also play a vital role to breach the boundaries and carry the pathogens to the take-for-granted safe sectors. 5
In today's society, companion animals such as dogs are considered family members. Various studies suggested that like food animals, dogs may also serve as an important reservoir of AMR pathogens. A significant proportion of registered dog breeds in and around Kolkata (West Bengal, India), who received recent antimicrobial therapy, were found to harbor carbapenem-resistant Enterobacteriaceae. 6 Apart from the registered breeds, India is home to a large number of street dogs, which do not have authentic pedigree records. These dogs, often referred to as “deshi” or “pariah,” usually stay in groups and thrive on the kitchenwaste, hotel refusals, or offals and live with no particular care or comfort.
Despite all the odds, they are very agile, alert, and are considered inherently resistant to many infectious diseases. Unlike the registered domesticated pure breed dogs, they receive little regular care for treatment and vaccination. Occasionally, dogs that are critically ill or grievously injured are brought to clinics by local people or persons involved in such nonprofit humanitarian organizations. Therefore, it is generally presumed that load of AMR pathogens should be low among such dogs and till date, no study has been undertaken to understand the status of such pathogens in nondescript dogs. This study, for the first time, records a detailed investigation on MDR Enterobacteriaceae infections in nondescript dogs in and around Kolkata and peri-urban areas.
Materials and Methods
Sample collection
During the study, which spanned ∼4 years (2018–2022), we collected the samples from 11 veterinary polyclinics and hospitals in and around Kolkata.
The samples collected from diseased nondescript dogs (n = 293) constitute the following: (1) urine sample from animals with urinary tract infection (UTI) and cystitis/urolithiasis through catheterization (n = 33); (2) direct swabs from skin/wound/ear from animals with pyoderma, nonhealing wounds, or otitis (n = 92); (3) swabs from surgical sites, including aspirates from infected bones (from animals with osteomyelitis during surgery; n = 80); (4) direct swabs from rectum from animals with diarrhea; n = 62); and (5) direct vaginal swabs from animals with pyometra and chronic vaginal discharge (n = 26).
Likewise, direct rectal swab (n = 18) and catheterized urine samples (n = 7) were collected from healthy dogs. These dogs were brought for deworming or castration with no apparent illness. Usually, many nongovernment organizations used to bring street dogs for such purpose.
Isolation and identification
Following brief selective enrichment in MacConkey broth, the samples were inoculated onto MacConkey agar, from which 3–5 lactose fermenting colonies were randomly chosen for morphological and biochemical tests for presumptive identification. Further, species-level confirmation of Escherichia coli (EC) and Klebsiella pneumoniae (KP) was made by molecular tests, as stated previously.3,7
Phylogroups of the E. coli isolates
All the confirmed E. coli (n = 365) isolated in this study were subjected to polymerase chain reaction (PCR)-based phylotyping to understand the association of phylotypes with drug resistance and disease conditions. 8
Antibiotic susceptibility testing
Antimicrobial susceptibility profile of the confirmed E. coli and KP isolates was determined by disk diffusion tests using commercially available antibiotic disks (Himedia; cefpodoxime [CPD—10 μg], aztreonam [AZT—30 μg], ceftazidime [CAZ—30 μg], cefotaxime [CTX—30 μg], ceftriaxone [CTR—30 μg], imipenem [IPM—10 μg], amikacin [AK—30 μg], ampicillin [AMP—10 μg], amoxicillin-clavulanate [AMC—30, 20/10 μg], sulfamethoxazole-trimethoprim [TMP—23.75/1.25 μg], nalidixic acid [NA—30 μg], enrofloxacin [ENR—5 μg], chloramphenicol [C—30 μg], tetracycline [TE—30 μg]), and by determining the minimum inhibitory concentrations (MICs) for IPM, ciprofloxacin, and colistin (MICIPM, MICCIP, and MICCOL).
While MICIPM and MICCIP were determined using commercially available E-test strips (HiMedia), a broth microdilution assay was used to determine MICCOL. CLSI 2022 breakpoints were used to classify the isolates in WHO-NET software. Isolates that exhibited resistance to three or more groups of antibiotics were categorized as MDR.
Phenotypic tests for conformation of ESBL and AmpC type β‑lactamase production
The isolates with reduced CPD/CAZ/FOX sensitivity were further screened using the combination disk (CD) test, ESBL E-test, cefoxitin-cloxacillin double-disk synergy (CC-DDS) tests to ascertain the ESBL/AmpC β‑lactamase production. 3
Phenotypic tests for confirmation of carbapenemase/metallo‑β‑lactamase production
The carbapenem nonsusceptible isolates were tested using three phenotypic tests: modified carba NP-direct test (CNPt-direct), 9 IPM/EDTA combined disk test (CDT), 10 and MBL-E test using commercially available metallo β-lactamase (MBL) test strips (HiMedia) to confirm carbapenemase/MBL.
Molecular characterization
PCR was used to screen the isolates for different genes encoding carbapenem resistance—blaKPC, blaNDM, blaOXA-48-like, blaIPM, blaVIM, b-lactamases such as—blaCTX-M (group1, 2 and 9), blaTEM, blaSHV, blaOXA, blaGES, blaVEB, and plasmid-mediated AmpC type β-lactamases (ACBLs)—blaFOX, blaCIT, blaMOX, blaCMY-2, blaDHA, blaACC, and blaEBC using different sets of primers.11–13 Furthermore, aforesaid isolates were investigated for the presence of tetracycline-resistance genes—(tetA, tetB, tetC, tetD, tetE), quinolone efflux pumps (qepA, oqxA, and oqxB), aminoglycosides—(armA, rmtB), sulfonamide-resistance (sul-1, sul-2, and sul-3), and plasmid-mediated quinolone-resistance (PMQR) determinants [qnrA, qnrB, qnrS, qnrC, qnrD, aac(6′)-Ib-cr].6,7,14–17
Different genes responsible for adherence and biofilm formations such as csgA, fimA, fliC, rcsA, rpoS, and sdiA were sought by PCR using the published primers.18,19 Details regarding oligonucleotides and control strains are described in Supplementary Table S1.
Plasmid-based replicon typing
Plasmids were isolated from the MDR E. coli and KP isolates using QIAprep Spin Miniprep Kit (Qiagen) following the instructions of the manufacturer and checked for their incompatibility (Inc) types using PCR-based method. 20
Enterobacteriaceae repetitive intergenic consensus PCR
To understand the genetic relatedness, all the MDR E. coli and KP isolates were subjected to Enterobacteriaceae repetitive intergenic consensus (ERIC) PCR as conducted previously. 21
Analysis
Data analysis was conducted in R software (version 4.0.3; R Foundation for Statistical Computing). Relationship among bacterial species, phylotypes, sampling source, resistance and virulence repertoire characteristics of all the MDR E. coli (n = 48) and KP (n = 11) isolates were presented in the form of a heatmap constructed using R software.
The association of particular resistance characteristics (ESBL/ACBL/carbapenemase production) with source of bacterial isolates and their phylotypes was summarized and visualized by multiple correspondence analysis (MCA). The ERIC profile of the MDR isolates was read to estimate the distance matrix by binary method, and Ward's minimum variance (WardD2) was considered to construct the hierarchical structure.
Results and Discussion
This study had been conducted during 2018–2022 spanning a period of 4 years, and it involved sampling from different veterinary hospitals and clinics in and around Kolkata. A total of 365 E. coli (328 isolates from diseased and 37 isolates from healthy dogs), 76 Klebsiella sp. (72 isolates from diseased and 4 isolates from healthy dogs) were isolated from 293 diseased and 25 healthy dogs.
The diseased dogs used for sampling can be classified in seven groups depending on their health status and organ/system involved. Details of sampling sources and pathogens isolated are illustrated in Table 1. Different clinical and pathological exhibits of the dogs, which were included in the study, are summarized in Supplementary Fig. S1.
Clinical Signalments of the Dogs and Details of the Pathogens Isolated During 2018–2022
UTI, urinary tract infection.
E. coli of diverse phylogroups from diseased and healthy dogs
The diverse phylogroups of the E. coli isolated from the diseased and healthy dogs are described in Table 1. Of the 47 E. coli isolated from the dogs with UTIs, cystitis, and urolithiasis (Supplementary Fig. S1), majority belonged to phylogroups A (44.7%) and B1 (44.7%) and B2 (8.5%). Phylogroup A (62.2%) and B1 (35.6%) predominated among the E. coli (90) from skin and ear (pyoderma and nonhealing wounds or ear wounds). The E. coli (n = 60) recovered from surgical sites could be distributed among five phylotypes, of which majority were A (46.7%), B1 (38.3%), and B2 (8.3%) types.
With respect to the 111 E. coli obtained from the rectal swabs of the diarrheic dogs, quite similar trend was observed (phylogroup A [48.6%], B1 [43%], and B2 [5.4%]). Isolates (20) from dogs with pyometra and chronic vaginal discharge also mirrored the same picture—A (45%), B1 (20%), and B2 (30%)—except a little higher frequency of B2 types. Only A and B1 phylogroups were detected among the E. coli from healthy dogs.
Although various studies from India and abroad have documented that companion animals could serve as a potent reservoir of drug-resistant pathogens, we have very limited data on such kind of pathogens from stray dogs—almost an omnipresent creature in the Indian subcontinent. Because of their close contact to human and environments in and around health care institutes and animal farms, they may serve as sentinel to reflect the present scenario of these pathogens in an area.
Phylogroups A (52.6%; 192) and B1 (38.6%; 141) emerged as the most predominant types in this study irrespective of the disease or health conditions of the dogs. Only 6.5% of the isolates belonged to B2 types, and D and F were least frequently detected. Associations between specific phylogenetic groups and pathogenic strains were numerated in several studies. B2 phylotype predominated among the extended-spectrum cephalosporin-resistant E. coli from diseased companion animals (dogs, cats, and horses) with extraintestinal infections, in the United Kingdom. 22
On the contrary, commensal phlotypes were found abundant in blaCTXM-15-producing ESBL E. coli from companion animals in Germany. 23 It appears geographical variation may influence such abundancy. Although majority of the E. coli isolates in this study belonged to commensal population (A and B1), a small proportion (>6%) was associated with virulence phylotypes. However, dogs with genital tract disorders were more frequently (∼30%) colonized with B2 types.
A recent study suggested that the animal E. coli isolates mostly belonged to three main phylogroups—A, B1, and B2. 24 Again, an investigation in Bangladesh reported A as the most predominant phylotype of the E. coli from animals, while isolates from human and environments were mainly of B1 types. 25 In human, several studies indicated predominance of B2 phylotype in urinary infections.26,27
Antibiotic sensitivity profile species and phylogroups-based analysis
The susceptibility profiles of all the isolates (EC [365] and KP [76]) were analyzed in WHO-NET software following CLSI 2022 guidelines. The isolates that exhibited resistance to three or more different antimicrobial classes were categorized as MDR (n = 59). Species and phylotypes-wise resistance patterns are illustrated in Fig. 1.
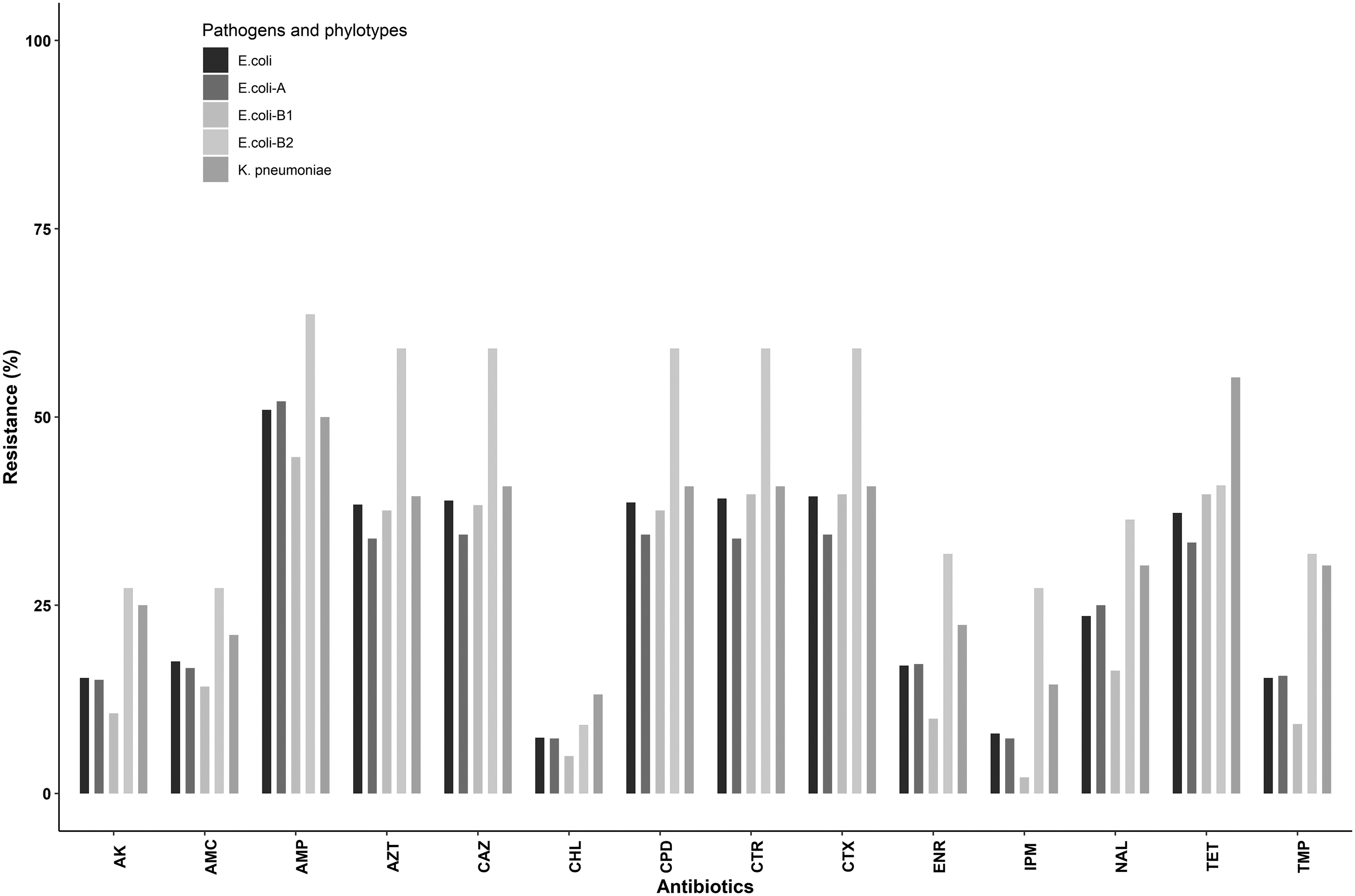
The resistance pattern of the Escherichia coli (365) and Klebsiella pneumoniae (76) isolates from stray dogs in and around Kolkata, India. This figure depicts the resistance profile of the following antibiotics—AK (30 μg), AMC (30 μg, 20/10 μg), AMP (10 μg), AZT (30 μg), CAZ (30 μg), C (30 μg), CPD (10 μg), CTR (30 μg), CTX (30 μg), ENR (5 μg), IPM (10 μg), NA (30 μg), TE (30 μg), TMP (23.75/1.25 μg). AK, amikacin; AMC, amoxicillin-clavulanate; AMP, ampicillin; AZT, aztreonam; C, chloramphenicol; CAZ, ceftazidime; CPD, cefpodoxime; CTR, ceftriaxone; CTX, cefotaxime; ENR, enrofloxacin; IPM, imipenem; NA, nalidixic acid; TE, tetracycline; TMP, sulfamethoxazole-trimethoprim.
In this study, KP isolates from canines were more frequently resistant to majority of the antibiotics than the E. coli isolates (Fig. 1), such as amikacin (EC: 15.3%, KP: 25%), amoxicillin-clavulanate (EC: 17.5%, KP: 21%), chloramphenicol (EC: 7.4%, KP: 13%), enrofloxacin (EC: 17%, KP: 22%), imipenem (EC: 8%, KP: 14.5%), nalidixic acid (EC: 25%, KP: 30.2%), tetracycline (EC: 37%, KP: 55%), and trimethoprim/sulfamethoxazole (EC: 15%, KP: 30%). However, both pathogens exhibited almost parallel resistance pattern toward β-lactams and monobactams such as ampicillin (EC: 51%, KP: 50%), cefpodoxime (EC: 39%, KP: 41%), ceftazidime (EC: 39%, KP: 41%), cefotaxime (EC: 39%, KP: 41%), ceftriaxone (EC: 39.2%, KP: 41%), and aztreonam (EC: 38%, KP: 39.5%).
In Fig. 1, resistance profile of the different phylotypes of E. coli isolates is depicted. As many of the E. coli strains belonged to three phylotypes—A, B1, and B2, these phylotypes were analyzed for estimating and comparison of resistance. Irrespective of the antimicrobials, all the strains of B2 phylotype exhibited higher resistance than A and B1 strains. Further, this difference was more apparent when resistance to cephalosporins was compared (Supplementary Table S2).
Altogether, canine E. coli and KP isolates exhibited frequent resistance to extended-spectrum cephalosporins, nalidixic acid, and tetracycline. We have also observed such resistance pattern among the isolates from food 28 and companion animals 6 of this region. Higher usage of cephalosporins and fluoroquinolones in both human and animal sectors could have led to the spread of the resistance traits in environment, 4 and even among the animals 3 where antimicrobial usage is negligible. Among the E. coli isolates, AMR was relatively more pronounced among the B2 phylotypes. Predominance of B2 phylotype has recently been reported among ESBLs from companion animals in the United Kingdom. 22
B2 phylotypes were previously reported to carry blaCTXM-15 gene in the multidrug-resistant IncFII plasmid. 29 On the contrary, report from Indian patients indicated that E. coli of type B1 were more likely to be drug resistant. 30 Previous studies conducted in a range of food animals indicated that cephalosporin-resistant E. coli often carried virulence characteristics of extraintestinal or uropathogenic E. coli,3,7,28 which mostly belonged to the virulent phylotype B2. This may be a reason for increasing report of B2 phylotypes among ESBLs.
Characteristics of multidrug-resistant E. coli and K. pneumoniae from stray dogs
Based on the AST profile, 59 isolates, which included 48 E. coli and 11 KP, were found multidrug resistant. ESBL was phenotypically evident in 33 isolates of which 29 were E. coli and 4 were KP (Table 2). Phenotypic assays confirmed AmpC β-lactamase production in 36 E. coli and 5 KP, while 12 E. coli and 6 KP were carbapenemase producers. ESBL-producing E. coli belonged to phylogroups B1 (12), A (7), B2 (4), F (3), and D (2). Phylogrouping could not be done for one isolate (CME33). Of the 36 ACBL-producing E. coli, 17 isolates belonged to phylogroup A. Other isolates were from phylogroups B1 (9), B2 (5), F (3), and D (2). Twelve carbapenemase producers were of five phylotypes—A (6), B1 (1), B2 (1), D (1), and F (3).
Characteristics of Multidrug-Resistant Escherichia coli and Klebsiella pneumoniae Isolated from Stray Dogs In and Around Kolkata, India
Based on phenotypic assays.
ACBL, AmpC type β-lactamase; ERIC, Enterobacteriaceae repetitive intergenic consensus; ESBL, extended-spectrum β-lactamase; F, female; M, male; MBL, metallo β-lactamase; ND, not detected; NT, nontypable.
In total, we were able to isolate 48 MDR E. coli and 11 MDR KP isolates in this study, and most of the isolates were ESBL and/or ACBL producers. Further, blaNDM-1 or blaNDM-5–mediated carbapenemase production was confirmed in 18 of them. In the recent past, companion animals emerged as one of the major sentinels of MDR pathogens. Because of multiple reservoirs, complex transmission routes, and presence of asymptomatic carriers, understanding the transmission and spread of ESBL/ACBL-producing MDR isolates in human or companion animals is very difficult, although there is sporadic evidence regarding the role of household transfer of ESBL-/pAmpC-producing Enterobacteriaceae. 31
Virulence factors—iucD, papC, and tsh, what we frequently noted among many of the MDR strains of dogs (data not shown) and serotypes such as O2, O7, O16, and O18, were previously reported in UPEC or ExPEC strains causing UTIs in human. 32 There are limited data on serotypes of the canine E coli isolates, particularly from India. Recently a study in Egypt reported E. coli of serotypes such as O18, O27, O55, O126, O148, O158, and O166 in dogs. 33
The association among species, phylotypes, source of infection, resistance genes, adherence, and virulence in the MDR isolates recovered in this study is presented in Fig. 2. Among the various phylotypes, we found multidrug resistance predominantly among B2 phylotypes (27%) followed by A (10.4%) and B1 (10%). The associations among the phylotypes, source of isolates, and their resistance characteristics are displayed in Fig. 3. It is evident from the correspondence analysis that the isolates of virulent phylotypes (B2, D, and F), particularly those isolated from reproductive tract, were more likely to be ESBL producers, while the isolates of rectal origin belonged to commensal phylotypes (A and B1).
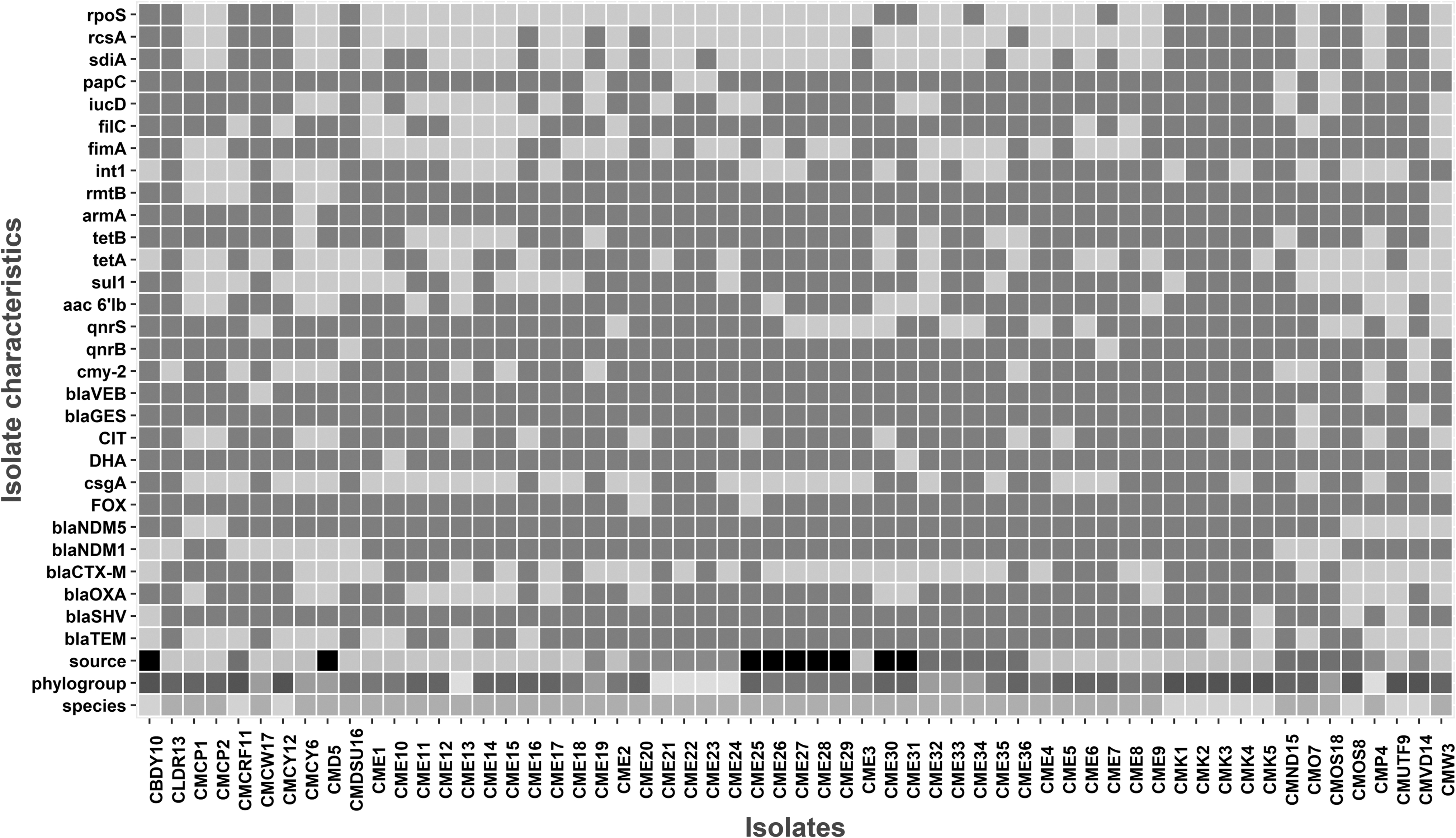
Heatmap of antibiotic resistance, biofilm-associated genes, and virulence determinants in the 48 MDR Escherichia coli and 11 MDR Klebsiella pneumoniae isolates from stray dogs in and around Kolkata, India. Phylogroups of the E. coli, sampling source, species are denoted by multicolor. light grey boxes indicate presence of a respective gene, while dark grey indicates its absence. MDR, multidrug resistant.
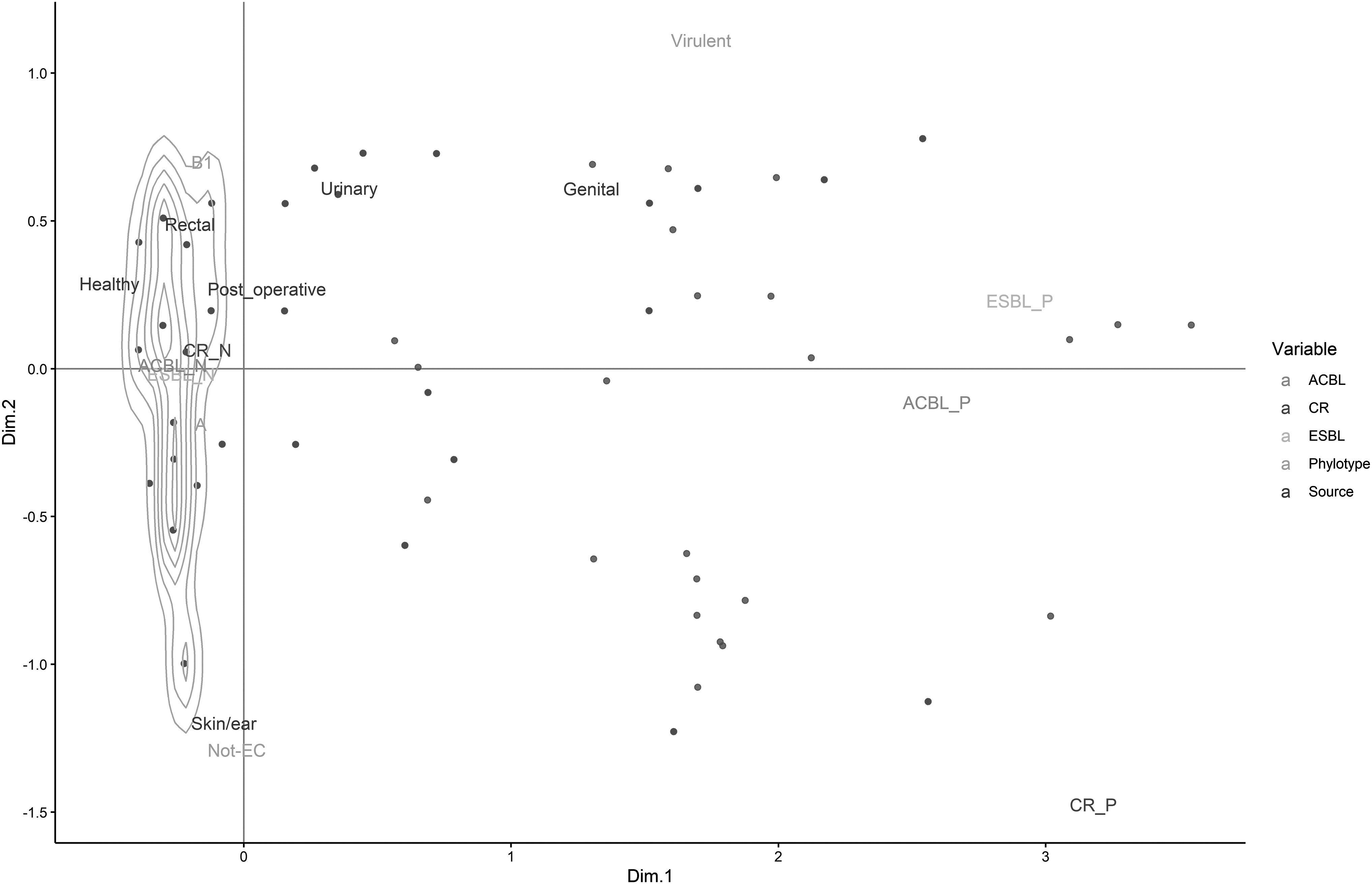
Multiple component analysis of antibiotic resistance characteristics (ESBL/ACBL or carbapenem resistance) for isolates from mongrels with their phylotypes (commensal—A and B1 vs. virulent—B2, D, and F) and sources. The loadings of each of these categories and variables on the first two principal components are shown. ACBL, AmpC type β-lactamases; ESBL, extended-spectrum β-lactamase.
We observed that ∼18% of the B2 isolates were also ESBL producing. B2 types were reported for their association with virulence traits causing UTIs. 34 Many of the B2 phylotypes in this study also carried extraintestinal virulence factors (data not shown), indicating that dogs could have contracted infections from human sources. 35 Previous workers from Spain suggested that the canine MDR E. coli were mostly of B1, D, and E phylogroups. 36 On the contrary, a report from Brazil suggested that canine E. coli were found mostly of B1, B2, and E types. 37 Of the 59 MDR strains recovered in this study, majority were from urogenital tract (24), skin or ear infection (16), and postsurgical infections sites (10).
More than 75% of the ESBL isolates were also from these sites of infections. We do not have any previous records to correlate the sites of infections with recovery of resistant strains in dogs or domestic animals. Our previous reports were mainly focused on recovery of such resistant strains from bovine milk or rectal/cloacal swabs of other animals. However, in human, UTI caused by MDR uropathogenic E. coli has become endemic in certain parts of India. 38 Five of the MDR isolates from urogenital tract infections in this study had carbapenem-resistance gene (NDM-1 or NDM-5), which is an emerging problem in human. 39
Of late, a study conducted in eastern India revealed that ∼43% of the samples from community-acquired UTI tested positive for ESBL infections, 40 and cefixime and ciprofloxacin were among the antibiotics against which resistance was frequently noted. In our study, many of the strains isolated from the urogenital tracts carried fimbria protein or adhesins such as fimA, filC, and csgA, which help them adhere to the epithelial cells of the tract only to reinforce their pathogenicity. 41 These cell surface proteins have the ability to promote motility, enzymatic breakdown, adhesion, colonization, and biofilm formation. 42
Further, a considerable number of isolates carried sdiA, rcsA, and rpoS genes, of which rpoS, an RNA polymerase sigma subunit, is responsible for controlling the expression of several other interconnected genes, which can drive the bacteria into stationary phase, particularly during stress like ultraviolet radiation, acid, temperature, osmotic shock, oxidative stress, nutrient deprivation, and exposure of antimicrobials. 43 The quorum sensing gene, sdiA, controls the adaptability of the bacteria to survive in the intestine and to facilitate systemic invasion in addition to making communications with other bacteria. 44
Distribution of ESBL genes among multidrug-resistant strains recovered from dogs
Among the 48 MDR E. coli isolates, blaCTXM-1 was detected in 29 isolates (Accession No. OP432544), whereas blaCTXM-2 (Accession No. OM898922) and blaCTXM-27 (Accession No. OM898921) were detected in 1 isolate each (2.08%). Eleven E. coli were positive for blaTEM-1 gene (Accession No. OL744243), and none had blaSHV gene. Fifteen of them possessed blaOXA gene (Accession No. OL744247). Of the 11 MDR KP, 4 were positive for both blaCTXM-1 and blaSHV (Accession No. OL744251) genes, whereas 7 had blaTEM (Accession No. OL744252). The gene blaOXA was detected in two KP isolates. Altogether, we detected ESBL production in 33 isolates.
Majority of the MDR strains recovered in this study were ESBL producers (n = 33). Van Den Bunt et al. reported on occurrence of ESBL producers in >10% of the dog population in the Netherlands, and most of them had blaCTXM-group 1. 45 Again, Benavides et al. recorded blaCTXM-1 type ESBL producers in 24% of the dogs in Chile. 46 In Taiwan, blaCTX-M-1 and blaCTX-M-2 were the most frequent ESBL types found in dogs. 47 In India, blaCTXM-1 appeared overwhelmingly prevalent, particularly among the hospitalized patients. 48
In concordance to our previous observation in hospitalized pets, 6 it seems that blaCTXM-1 emerged as the main ESBL gene in human and animals, at least in this part of the world. Several workers reported on the predominance of blaCTX-M-15 gene among Indian ESBL isolates. 49 The blaCTX-M-27 was detected in a single isolate. It is an infrequent global blaCTX-M allele that differs by only one nucleotide from blaCTX-M-14, resulting in the change of amino acid at position 240. 50 A study conducted in Germany on ESBL producers from livestock, humans, companion animals, food, and the environment detected blaCTX-M allele only in human isolate. 51
Similar to our observation, blaOXA was frequently reported in human patients in and around Kolkata. 52 The blaOXA gene is responsible for mediating resistance to ampicillin and cephalosporins, and some variants are also responsible for reducing susceptibility toward reserved antibiotics such as carbapenem. 53 Of late, there is a report on spillover of blaOXA-181–encoding plasmids to Enterobacter hormaechei and Serratia nevei causing canine and feline opportunistic infections in Thailand. 54 Further, blaOXA-48 encoding IncL plasmid was reported in dogs from France, suggesting that dogs can also play a vital role in transmission of these resistance genes.
This study also recorded presence of blaVEB (Accession No. OM977027) and blaGES (Accession No. OM898923) type ESBLs altogether in three E. coli and one KP isolates. Both the genes—blaVEB and blaGES—are relatively rare and under-reported in animals. Since the first report of blaVEB-1 from Vietnam in 1998, it continued to be sporadically reported from various parts of southeast Asia and other parts of the world. In India, it was reported in an outbreak of neonatal sepsis caused by MDR Proteus mirabilis 55 and in patients with recurrent UTI from north eastern states of India. 56
Plasmid-mediated ACBL and carbapenemase genes among multidrug-resistant strains recovered from dogs
Of the 48 MDR E. coli isolates, blaCMY2 (Accession No. OL744245) were detected in 10 isolates, while blaDHA and blaFOX genes were detected in 2 isolates, each. Among the 11 MDR KP blaCMY2 (Accession No. OL744238) detected in 3 isolates. In total, 18 isolates had carbapenemase gene blaNDM (blaNDM-1 [11] and blaNDM-5 [7]).
Quite similar to this study, Manoharan et al. also reported predominance of plasmid-mediated ACBL like blaCMY, blaDHA, and blaFOX in human from five different tertiary care centers of India. 57 A different study conducted in Iran also showcased the predominance of these resistance determinants 58 among hospitalized patients. Plasmid-mediated AmpC b-lactamases, which are actually derived from chromosomally encoded AmpC, may vary in their prevalence depending on the geographical location and the type of hospital. 59 Isolates with the pAmpC genes can be transferred more easily across the species, and are often MDR to cause therapeutic failures in nosocomial infections and even to complicate the treatment of community-acquired infections.60,61
The blaNDM-producing MDR strains were detected in ∼3.9% of the dogs in China. 62 Even we also found several carbapenem-resistant strains from hospitalized pets. 6 Since carbapenems are not used in dogs in India, the detection of carbapenem-resistant pathogens is quite unlikely. However, most of the carbapenem-resistant strains carried plasmids of multiple replicon types—IncFIA, IncFrepB, IncFIB, and IncFIC. All these plasmids are usually occupied with MDR cassettes, and selection pressure due to some other antibiotics could have led to the acquisition and dissemination of carbapenem-resistance genes. It is not unlikely that these plasmids, which are commonly detected among human or environmental pathogens, 63 have been picked up by the microbiota of stray dogs.
Other resistance determinants among multidrug-resistant strains recovered from dogs
Other resistant determinants such as sulfonamide-resistant gene sul-1 gene were detected in 19 of the MDR E. coli (Accession No. OL744244) and 6 of the MDR KP (Accession No. OL744250) isolates. This study also detected the tetracycline-resistance genes such as tetA (Accession No. OL699944) and tetB in 22 and 14 MDR E. coli isolates, respectively. In addition, four KP isolates had tetA gene (OL744242).
Sulfonamide- and tetracycline-resistance genes have been frequently detected in human, nonhuman primates, and domestic animals suggesting the role of common ecological niche for transmission of AMR pathogens and genes. 64 It is not impossible that prolonged persistence of antibiotics (or their residues) such as sulfonamide, tetracycline, and quinolone in environment and soil because of their slow degradation 65 could have facilitated the spread of their resistance factors in the stray dogs.
Quinolone drug-resistant determinants such as qnrB, qnrS, and aac-6′-lb-cr were also found in this study. The qnrS were detected in 13 E. coli (Accession No. OM977024) and 2 KP isolates, qnrB gene was found in 2 E. coli (Accession No. OM977028) and in 1 KP (Accession No. OM977029), and aac-6′-lb-cr gene in 13 E. coli (Accession No. OM977026) and 1 KP isolate, only.
All the PMQR genes including aminoglycoside acetyltransferase [AAC-(69)-Ib-cr] are known to confer low level of quinolone resistance, and thus isolates having PMQR genes are often missed in disk diffusion test. PMQR genes often play the pivotal role for selection of isolates with higher resistance to fluoroquinolones, a frontline therapy in UTI and typhoidal illness; thus, their detection remained an essential part of AMR surveillance. Three isolates had both qnrS and aac-6′-lb-cr; cocarriage of qnr and aac-6′-lb-cr was previously described in clinical isolates (KP) with higher resistance to drugs such as aminoglycosides, cephalosporins, and quinolones. 66
In this study too, we found these three isolates as ESBL producers, and one of them carried armA and rmtB genes, indicating the possibility of these resistance determinants in single drug resistance cassette. In total, we detected class I integron in 17 E. coli and 7 KP isolates, and armA and rmtB genes in 2 and 5 MDR E. coli, respectively; most of them were ESBL producers. Many of the ESBL producers were frequently reported to carry armA and rmtB because of their location on the same plasmid along with ESBL genes. 67
Diversity of plasmid replicon types of the multidrug-resistant strains recovered from dogs
Plasmid replicon typing showed predominance of IncF replicon types. FIA (EC: 36; KP: 4), FIB (EC: 18; KP: 4), FIC (EC: 7 KP: 1), FIIA (KP: 2), and FrepB (n = 33) were frequent among the E. coli isolates. Besides, I1γ plasmid types were detected in 20 MDR E. coli and 3 KP isolates. A few isolates had HI-1 (EC: 2 KP: 1), A/C (EC: 3 KP: 1), and N (KP: 1) replicon types.
Similar to our observation, plasmids of diverse Inc groups are recorded among the drug-resistant strains such as blaCTXM-1 type ESBL producers or quinolone-resistant E. coli from various domestic animals, including dogs. 68 Most of the environmental pathogens were reported to carry multiple IncF family plasmids of FIA, FIB, and FII combination types resulting from higher selection pressure associated with cumulative antimicrobial exposure. 69 Of late, we have detected such multidrug-resistant plasmid in caprine 3 and canine E. coli 6 isolates. Multidrug-resistant plasmids are common in food animals because of their close interaction with human, other animals, and environment, which may serve as the source of diverse pathogens and their resistant determinants. 70
Although most of the isolates in this study carried more than one plasmid, it was not possible to ascertain whether the resistance genes were located on one or multiple plasmids. Nevertheless, IncF plasmids particularly FIA and FrepB types were previously reported as major vehicle to ferry resistance genes. 71 The carriage of drug resistance genes on transposable mobile elements and mobile plasmids renders the resistome highly dynamic, and that can be transmitted to other animals, in-contact human and spread to distance through environment.
ERIC PCR differentiated the E. coli strains in five clades A–E. Clade C contained majority of the isolates (n = 14) followed by E (n = 11) and A (n = 10) (Table 2 and Supplementary Fig. S2). Clades D and B consisted of eight and five isolates, respectively. All the isolates were typeable and yielded different fingerprinting profiles. We could not find any relation of the ERIC clades of the isolates with respect to the source of sampling, phylotypes, and resistome. The 11 MDR KP isolates were divided among 4 clusters A–D. While A, B, and D consisted of 2 isolates each, cluster C contained 5 isolates.
In general, it appears from the ERIC PCR that the MDR E. coli isolates from stray dogs were genetically diverse with little evidence of any similarity with respect to sample source, resistome, or phylogroups, except the fact that majority of the isolates in cluster E belonged to phylotype A. Conversely, we could notice two carbapenemase-producing KP isolates from surgical sites grouped together in cluster A. Most of the UTI isolates clustered together either in B or in a subcluster of C. Therefore, it seems that sampling source could have a bearing upon the genomic relatedness among the MDR KP isolates. However, such conclusion may be misleading, as we had limited number of MDR KP isolates.
Previous studies indicated that ERIC PCR could distinguish isolates from typical sampling sites or a particular species. It had been also reported that isolates from a particular farm, food, or from a particular region could be recognized using ERIC PCR. 72 However, it is only possible provided that the isolates have a high degree of sequence similarity. Nonetheless, ERIC PCR revealed a wide genomic diversity among the MDR isolates in this study, which is not unusual, as the stray dogs could have contracted the infections from diverse sources.
Although we usually focused more on food animals for carrying out the AMR surveillance, increasing detection of drug-resistant pathogens in pet animals such as dogs has emerged as a serious cause of concern because of their proximity to human. This study pointed out the spread of the resistance determinants in stray dogs with no previous direct exposure to antibiotics, suggesting a vital role of environment in dissemination of antimicrobial-resistant pathogens and resistance genes. Most of the isolates also harbored plasmids of multiple incompatibility types, indicating the potential role of these mobile genetic elements in transmission of AMR genes.
Most of the multidrug-resistant isolates, including the ESBL producers, were isolated from reproductive or urinary sources and belonged to virulent phylotypes. This study indicated that the mongrels may act as a potential reservoir of drug-resistant pathogens, including the carbapenemase producers. Emergence of such resistance even against the reserved drugs is quite alarming; therefore, multidimensional tracing of these pathogens in veterinary setting, human hospitals, community, and other in-contact animals is necessary for establishment and strengthening of antimicrobial stewardship and control policy.
Ethical Approval
The study was conducted following the guidelines prescribed by Institutional Animal Ethical Committee.
Footnotes
Acknowledgments
The authors are thankful to Director, ICAR-IVRI, Izatnagar for providing necessary support for carrying out the work. They acknowledge the inputs of Dr. A. Sen, HoRC, ICAR-IVRI (Kolkata, India) for revising the article.
Authors' Contributions
J.B., S. Biswas, D.B., and M.H. performed investigation. J.B. and S. Batabyal provided methodology and conceptualization. S. Bandyopadhyay assisted with data curation, formal analysis, visualization, software, funding acquisition, project administration, and supervision. A.K.D., P.K.N., I.S., P.D., and S. Bandyopadhyay contributed to writing—review and editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
The research grant received from DBT-TANUVAS CRC&N (BT/ADV/CANINE HEALTH/TANUVAS/2017) project is thankfully acknowledged.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.