
Abstract
This study investigates two isolates of Salmonella enterica subspecies enterica serovar Enteritidis (S. Enteritidis), designated Sal B and Sal D, isolated from the blood and pleural fluid, respectively, of the same patient. Drug susceptibility testing revealed significant differences: Sal D exhibited greater resistance to ticarcillin/clavulanate, piperacillin/sulbactam, and ciprofloxacin compared with Sal B. Morphologically, Sal B formed rougher and drier colonies than Sal D at 37°C. Sal B demonstrated significantly stronger biofilm-forming ability and higher adhesion capacity to HaCaT cells than Sal D, whereas Sal D showed superior adaptation to acidic conditions (pH 3.0). Virulence assays indicated no significant differences between the isolates, suggesting comparable pathogenic potential. Comparative genomic analysis showed high gene content conservation but identified two nonsynonymous single-nucleotide polymorphisms (nsSNPs) and an insertion in the envZ and siiE genes. These genetic variations may account for the observed differences in drug susceptibility and biological characteristics. Collectively, these findings suggest that S. Enteritidis can undergo adaptive changes in response to distinct host environments, influencing drug resistance, adhesion, and acid resistance. This knowledge may inform future strategies for the treatment and prevention of Salmonella infections.
Introduction
Non-typhoidal Salmonella (NTS) is a major cause of human diarrhea and self-limiting gastroenteritis, contributing to approximately 93.8 million cases of gastrointestinal illness and 150,000 deaths globally each year. 1 Among NTS serotypes, Salmonella enterica subspecies enterica serovar Enteritidis (S. Enteritidis) is the most frequently isolated worldwide, accounting for approximately 30% of Salmonella infections reported in China. 2 The primary infection associated with S. Enteritidis is acute gastroenteritis, typically resulting from the consumption of contaminated poultry meat and eggs. 3 However, S. Enteritidis can also cause invasive extra-intestinal diseases,4,5 including bacteremia, 6 myocarditis, 7 meningitis, 8 and other focal infections,9,10 often with high mortality rates. Invasive S. Enteritidis disease presents with non-specific clinical features, which can be challenging to differentiate from typhoid fever or other febrile illnesses and are typically unrelated to diarrhea.11,12 Immunocompromised individuals, malnourished children, and the elderly are particularly vulnerable to these infections.8,13–15 Moreover, certain S. Enteritidis isolates exhibit high-level resistance, posing a significant threat to global public health.4,16,17
The pathogenicity of S. Enteritidis is multifaceted, involving multiple virulence factors critical to its infection process. Well-studied virulence factors include fimbriae, flagella, and Salmonella pathogenic islands (SPIs), each contributing to different stages of infection.17–19 SPIs, particularly SPI-1 and SPI-2, are crucial for host interactions, encoding the type 3 secretion system (T3SS) essential for host cell invasion and intracellular survival and replication.20–22 Other virulence factors, such as adhesins, 23 and biofilm-related proteins 24 are implicated in adhesion, invasion, immune evasion, antibiotic resistance, and horizontal transfer of virulence traits. This intricate interplay of virulence factors underscores the sophisticated strategies employed by S. Enteritidis to establish and sustain infection within the host.
In this study, we isolated S. Enteritidis from distinct extra-intestinal sites (blood and pleural fluid) in patients at various time points. We hypothesized that these isolates might be genetically homologous but could have undergone adaptive changes in antibiotic resistance and virulence to survive in different host environments. It is important to acknowledge that S. Enteritidis is characterized by its high clonality, which may complicate the distinction between persistent infection and reinfection. Finally, a comparative genomic analysis was conducted to identify potential genetic variations. These findings will enhance our understanding of the pathogenicity and resistance mechanisms of S. Enteritidis, providing valuable insights for source tracking and disease prevention strategies.
Methods and Materials
Case introduction
Figure 1 depicts the clinical timeline of the case. The patient, a 49-year-old male with a history of thoracic aortic aneurysm (TAA) treated with stent implantation 2 months earlier, was admitted to Beijing Friendship Hospital on March 28, 2023, presenting with hemoptysis. He was diagnosed with pneumonia and a thoracic aortic false aneurysm (TAFA). During his hospitalization, he underwent surgical resection of the TAFA and the right lower lobe, with a positive blood culture isolating Sal B. The patient was discharged following clinical improvement. He was subsequently readmitted due to chest pain, during which, another isolate, Sal D, was isolated from pleural drainage. The patient was discharged again thereafter. Notably, stool samples collected during both hospitalizations tested negative for Salmonella.

Timeline of the case. TAA, thoracic aortic aneurysm; TAFA, thoracic aortic false aneurysm; MEC, meropenem; VAN, vancomycin.
Isolation and identification
Bacterial isolates were identified as follows: suspicious colonies on Salmonella-Shigella agar were analyzed using matrix-assisted laser desorption ionization-time of flight mass spectrometry (Biomérieux, France). Biochemical reaction was performed using VITEK2 compact system (Biomérieux, France). Identified Salmonella isolates were serotyped by O and H antigen slide agglutination tests (Tianrunbio, China) according to the White–Kauffmann–Le Minor scheme. All isolates were stored in brain-heart infusion broth with 20% glycerol before experimentation. This study adhered to the Declaration of Helsinki and was approved by the Ethics Committee of Beijing Friendship Hospital, Capital Medical University (L-2021-096).
Antimicrobial susceptibility testing
The minimum inhibitory concentration of each isolate against 14 antibiotics was determined using VITEK2 compact system (Biomérieux, France). The test strains were inoculated onto blood agar plates and incubated overnight at 37°C. The bacterial suspension was prepared to a McFarland standard of 0.5 to 0.63 in 0.45% sodium chloride, using the VITEK DensiChek densitometer. Antibiotics tested included ticarcillin/clavulanate, piperacillin/sulbactam, ceftazidime, cefoperazone/sulbactam, cefepime, aztreonam, imipenem, meropenem, ciprofloxacin, levofloxacin, doxycycline, minocycline, colistin, and trimethoprim-sulfamethoxazole. Susceptibility interpretations followed Clinical and Laboratory Standards Institute guidelines. 25 Escherichia coli ATCC 25922 served as quality control strain. Each bacterial strain was tested in triplicate.
Biological characteristics
Colony morphology assay
Colony morphology was assessed as described previously. 26 Briefly, target isolates were inoculated into Luria-Bertani (LB) liquid medium and cultured overnight at 37°C. One microliter was spotted onto LB agar plates containing 20 µg mL−1 Coomassie Brilliant Blue and 40 µg mL−1 Congo Red, incubated at 26°C and 37°C for 96 hours, and photographed using a digital camera (PowerShot G7X Mark II; Canon).
Acid resistance test
Acid resistance was evaluated as described previously, 27 with minor modifications. Target isolates were inoculated into LB liquid medium and incubated overnight at 37°C until an optical density at 600 nm (A600) reached 2.5. One milliliter of bacterial suspension was centrifuged, resuspended in M9 minimal medium containing 10 mM glucose (M9G, pH 3.0), and incubated at 37°C for 2 hours. Samples were serially diluted with phosphate-buffered saline (PBS), plated onto LB agar, and survival rates were determined.
Biofilm experiment
Biofilm formation was assessed as previously described. 28 Bacterial suspensions in tryptic soy broth (TSB) were prepared from pure colonies. Each well of a 96-well plate received 200 μL of the suspension, achieving in a final concentration of approximately 3 × 105 colony-forming units (CFU)/mL. Negative control wells contained only TSB, while positive controls included Staphylococcus aureus ATCC 29213 in TSB. After 48-hour of incubation at 37°C, wells were washed twice with PBS followed by the addition of 90 μL TSB and 10 μL of MTT reagent for an additional 4-hour incubation. Subsequently, 100 μL formazan solution was added, and absorbance at 490 nm was measured with an ELISA reader. Results were calculated by subtracting the average ODs of negative controls from experimental wells.
Adhesion assay
The cell adhesion experiment was carried out as previously described. 29 Overnight cultures in the LB medium were utilized to inoculate fresh LB media, resulting in an initial optical density (OD600) of 0.05. Cultures were incubated at 37°C for 3 hours with orbital shaking at 200 rpm. Bacteria were collected, washed with cooled PBS, and used to infect HaCaT cells seeded in 96-well plates with Dulbecco’s Modified Eagle Medium at 37°C and 5% CO2. When the cell density reached 70%, HaCaT cells were infected with bacteria in the exponential phase at a multiplicity of infection of 10 and incubated for another 2 hours in 24-well plates. Supernatants were discarded, cells were washed with sterile PBS to remove non-adherent bacteria, lysed with 0.1% Triton X-100, and bacterial CFUs were counted by plating on LB agar. Experiments were performed in triplicate.
Motility assay
The motility assay followed previously established methods. 30 A single colony was inoculated into 3 mL of LB liquid medium for overnight culture. Two microliters of bacterial suspension were spotted onto LB semi-solid medium (containing 0.3% agar). Motility was assessed by measuring the diameter of the bacterial migration zone from the edge of the inoculation point after incubation at 37°C for 7 hours. Images were captured using a gel imaging system. Each plate was inoculated with both Sal B and Sal D simultaneously, with three plates for parallel control.
Galleria mellonella infection assay
The Galleria mellonella infection assays were performed as previously described, with slight modifications. 31 Final-instar larvae, used within 3 days of receipt, were divided into three weight-matched groups of ten larvae each: a control group treated with 10 μL 0.45% saline, and two treatment groups received 10 μL bacterial suspension (105 CFU of Sal B and Sal D). Injections were administered into the last proleg of each larva’s right side using a 10 µL Hamilton syringe. Larvae were maintained in darkness at 37°C and observed every 12 hours for 72 hours, with survival status recorded and a survival curve plotted.
DNA extraction
Genomic DNA was extracted from each isolate using the QIAamp DNA Mini Kit, according to the manufacturer’s instructions. 32 Briefly, bacterial cultures were centrifuged, and the pellets were resuspended in buffer GA with Proteinase K, followed by incubation with buffer GB. Ethanol was then added, and the mixture was transferred to an adsorption column for further processing. Samples were washed with buffers GD and PW before eluting the DNA with TE buffer. Following centrifugation, the eluate was collected, and DNA was eluted with TE buffer. After centrifugation, the eluent was collected, quantified using a Thermo Scientific Qubit® 2.0 Fluorometer, and stored at −20°C for downstream applications. DNA integrity was verified by agarose gel electrophoresis.
Genome sequencing and bioinformatic analysis
Whole-genome sequencing was performed using Illumina NovaSeq PE150 at Beijing Novogene Bioinformatics Technology Co., Ltd. Approximately 0.2 μg of DNA per sample was used for library preparation using NEB Next® Ultra™ DNA Library Prep Kit for Illumina. Genomic DNA was sonicated to 350 bp fragments, end-polished, A-tailed, and ligated with Illumina adapters, followed by PCR amplification. PCR products were purified, DNA concentration was measured by Qubit®3.0 Fluorometer, and libraries were analyzed by Agilent 2100 Bioanalyzer and quantified by real-time PCR (>2 nM). Index-coded samples were clustered on a cBot Cluster Generation System with Illumina PE Cluster Kit. The DNA libraries were then sequenced on an Illumina platform to generate 150 bp paired end reads. Gene functions predicted using the Kyoto Encyclopedia of Genes and Genomes (KEGG) 33 and Clusters of Orthologous Groups (COG) 34 were utilized. Pathogenicity and drug resistance analyses were performed using Virulence Factors of Pathogenic Bacteria (VFDB) 35 and Antibiotic Resistance Genes Database (ARDB). 36 In silico serotyping and sequence typing were performed as reported literatures.37–40
Comparative genomic analysis
Comparative genomic analysis between Sal B (reference strain) and Sal D was performed using MUMmer. 41
Accession numbers
Nucleotide sequence data generated in this study are deposited in the GenBank database with the following accession numbers: SAMN44286458, SAMN44286459 (BioProject: PRJNA1172950).
Statistical analysis
Statistical analysis was carried out using SPSS 22.0 (IBM Inc., Chicago, IL). One-way ANOVA and unpaired t tests were employed to assess statistical significance, with a significance level set at p < 0.05. When appropriate, Bonferroni post tests were conducted to compare the means of different experiments directly.
Results
Isolation and identification
Both bacterial isolates, isolated from the patient’s blood and pleural fluid at different times, were identified as S. Enteritidis (including slide agglutination and in silico serotyping). Biochemical characteristics were largely similar, except for
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing results (Table 1) indicated that both isolates shared the similar susceptibility profiles for most antibiotics, with notable differences in ticarcillin/clavulanate (intermediate vs. resistant), piperacillin/sulbactam (susceptible vs. intermediate), and fluoroquinolones ciprofloxacin (susceptible vs. resistant) between Sal B and Sal D, respectively.
The Results of Antimicrobial Susceptibility Testing (MIC, μg/mL)
Ssusceptible; Iintermediate; Rresistant.
ATM, aztreonam; CAZ, ceftazidime; CIP, ciprofloxacin; COL, colistin; CSL, cefoperazone/sulbactam; DOX, doxycycline; FEP, cefepime; IPM, imipenem; LVX, levofloxacin; MEM, meropenem; MIC, minimum inhibitory concentration; MNO, minocycline; SXT, trimethoprim-sulfamethoxazole; TCC, ticarcillin/clavulanate; TZP, piperacillin/sulbactam.
Biochemical Characteristics of the Isolates
Differential biochemical characteristics.
5KG, 5-keto-glucoside; AGAL, α-galactosidase; AGLTp, glutamine arylamidase; APPA, alanine-phenylalanine-proline arylamidase; Balap, β-alanine arylamidase; BGAL, β-galactosidase; BGLU, β-glucosyl enzyme; BNAG, β-N-acetyl amino glycosidase enzymes; CIT, citrate; CMT, COURMARATE; dCEL, d-cellobiose; dGLU,
Biological characteristics
To explore differences in the biological characteristics of the two S. Enteritidis isolates from distinct sites within the same patient, we conducted assays for morphology, biofilm formation, adhesion, motility, acid tolerance, and Galleria mellonella infection (Fig. 2).
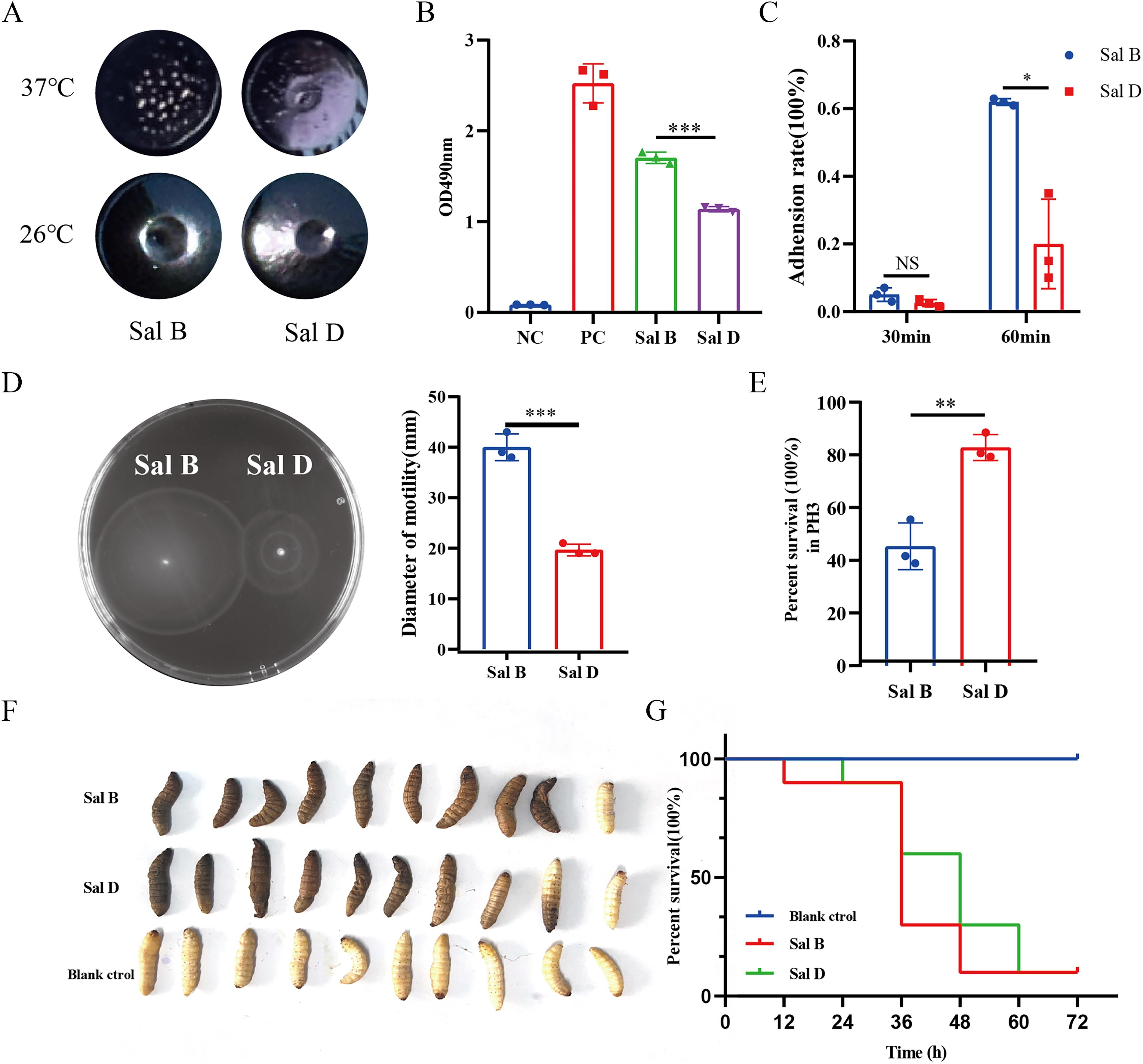
Characteristics contrast of Sal B and Sal D.
Colony morphology differed at different temperatures: at 26°C, both isolates formed smooth colonies with sunken centers in concentric circles, showing minimal differences. At 37°C, however, colonies were rougher, with Sal B forming notably rougher and drier colonies than Sal D. In addition, biofilm formation ability was significantly higher in Sal B than in Sal D (p ≤ 0.001). Moreover, adhesion capacity to HaCat cells showed no difference at 30 minutes but were significantly higher for Sal B than it was for Sal D at 60 minutes (p ≤ 0.05). Motility, assessed by bacterial migration zone diameter, was stronger in Sal B than in Sal D (p ≤ 0.001). Acid adaptation at pH 3.0 was superior in Sal D compared with Sal B (p ≤ 0.01). Nevertheless, the Galleria mellonella infection assay revealed no significant differences in virulence between the two isolates.
General genomic features
To characterize the genetic characteristics, draft genome sequences were obtained for two isolates. With regard to the draft genome sequences and the predictions of coding genes, whole-genome maps of S. Enteritidis were constructed (Table 3). The final assemblies of two genomes of S. Enteritidis showed that the genomes were similar in size and G+C contents. Genome annotation yielded 4,581 and 4,085 genes for Sal B and Sal D, respectively.
Basic Genomic Information
GIs, Genomic islands; TR, Tandem repeat.
The predicted genes were annotated using KEGG metabolic pathway categorization and COG categories (Fig. 3 and Supplementary Fig. S1). The distributions of annotations in Sal B and Sal D were found to be comparable. Taking Sal B as an example, in terms of KEGG pathways, metabolism had more genes than any other category. Notably, carbohydrate metabolism (336 genes), amino acid metabolism (214 genes), and cofactor and vitamin metabolism (199 genes) were the most abundant. The top three categories of COG-annotated genes were carbohydrate transport and metabolism (406 genes), amino acid transport and metabolism (383 genes), and transcription (338 genes).

KEGG metabolic pathway classification
Resistance genes and virulence genes in Sal B and Sal D
The Sal B and Sal D isolates were annotated with resistance and virulence genes using CARD, ARDB, VFDB, and T3SS databases. 42 Both isolates encoded for the same 36 resistance and 153 virulence genes (Table 4). Resistance genes were classified into 11 categories: resistance-nodulation-cell division transporter (RND), major facilitator superfamily transporter (MFS), ATP-binding cassette superfamily (ABC), multidrug and toxic compound extrusion family (MATE), small multidrug resistance family, β-lactamase, β-lactamase/RND, RND/ABC, RND/MFS/ABC, RND/MFS, and others. Genes such as basitem-1, marA (β-lactamase), acrA, acrB, sdiA, marR, rsmA, CRP, adeF (RND), emrB, emrR (MFS), mdtK (MATE), marA (β-lactamase/RND), and soxS (RND) have been linked to differences in Sal B versus Sal D resistance phenotypes (ticarcillin/clavulanate, piperacillin/sulbactam, and ciprofloxacin). The virulence genes were mainly classified into eight categories: effector delivery system, adhesins, flagella and chemotaxis, enterobactin synthesis and transport, lipopolysaccharide, exotoxin, stress survival, antimicrobial activity/competitive advantage, and resistance to complement killing. It was found that the effector delivery system possessed the largest number of virulence factors, consisting mainly of T3SS, T6SS, and type 1 secretion system (T1SS). The second highest number of virulence factors belonged to adhesions. Both of them played major roles in Salmonella invasion and infection.
Comparative genomic analysis
In order to reveal the reasons for the differences in the biological characteristics of the two isolates, including virulence and resistance, we conducted comparative genomic analyses between Sal B (reference strain) and Sal D. Both the chromosomal and plasmid genomes of Sal B and Sal D exhibit high similarity (Fig. 4), with 39 SNPs and 3 insertions/deletions identified (see Supplementary Fig. S1). After excluding synonymous mutations and non-coding region variants, 15 nonsynonymous single-nucleotide polymorphisms (nsSNPs, affecting envE and siiE) and 1 insertion (encoding putative transposase) were retained (Table 5).
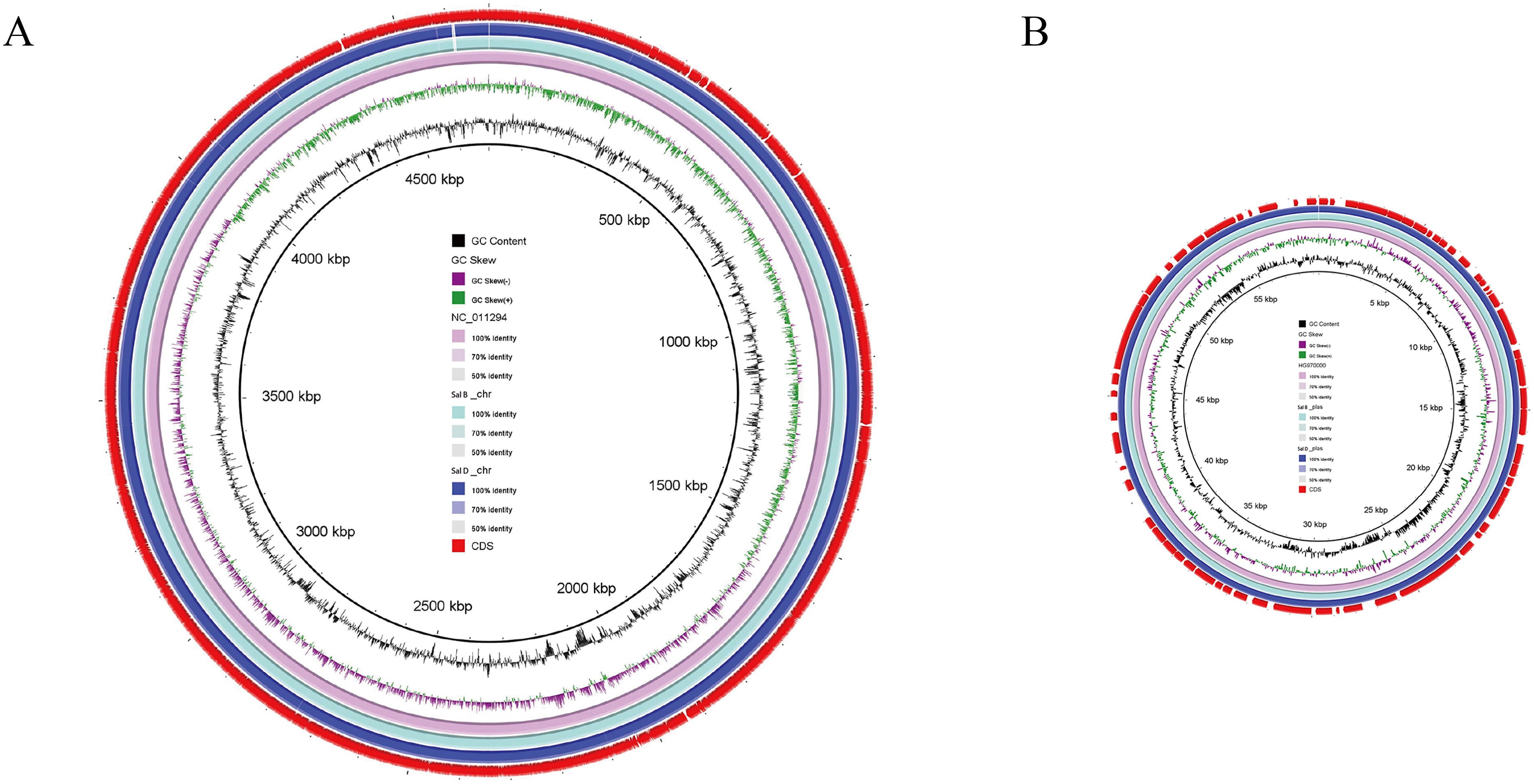
Comparison of chromosomal genomes
Resistance Genes and Virulence Genes in Sal B and Sal D
T1SS, type 1 secretion system; T3SS, type 3 secretion system.
INDELs and nsSNPs in Coding Sequences Between Sal B (Reference Isolate) and Sal D
INDELs, insertions/deletions; nsSNPs, nonsynonymous single-nucleotide polymorphism.
Discussion
S. Enteritidis is a significant pathogen capable of causing severe invasive infections beyond the gastrointestinal tract. 4 While acute enteritis is the most common presentation, S. Enteritidis can progress to systemic diseases, such as bacteremia and focal infections. 43 In the case, two S. Enteritidis isolates were isolated from the blood and pleural fluid of a middle-aged male patient with a history of stent implantation, separately. Moreover, both isolates belong to the most prevalent serogroup O:9, 44 and they shared the same sequence typing, ST11. The uniqueness of this case lies in the fact that the isolate was first isolated from blood rather than from the typical fecal or contaminated food sources. 3 Although the infection source of S. Enteritidis remains unclear, the possibility that S. Enteritidis contributed to TAA cannot be excluded.45,46 Notably, the isolates exhibited differences in resistance profiles, morphology, biofilm formation, adhesion capabilities, and acid adaptation, suggesting adaptive evolution in response to distinct host environments (blood vs. pleural fluid).
The question of whether the Salmonella infecting the patient represents a homogeneous population or heterogeneous subpopulations is complex. Given the patient’s prolonged antibiotic treatment (over 2 months) and the observed phenotypic differences, we hypothesize that selective antibiotic pressure drove adaptive mutations. This hypothesis was preliminarily supported by analyzing multiple colonies (>3 colonies) during experiments. It is worth noting that S. Enteritidis is known for its high clonality, which may complicate the differentiation between persistent infection and reinfection. While future studies will rigorously explore clonal expansion versus polyclonal selection, current genomic data confirm the homologous origin of Sal B and Sal D, validating subsequent mechanistic analyses of strain-specific variations.
The differences in antibiotic resistance profiles between Sal B and Sal D have implications for clinical treatment. Sal D’s reduced susceptibility to β-lactams and fluoroquinolones may reflect adaptive evolution under host immune pressure and antibiotic exposure. This adaptive evolution has allowed Sal D to gradually acquire resistance to certain antibiotics during its interactions with the host immune system and antibiotics. We speculate that the single-base mutation in the sensor kinase envZ affects its cognate response regulator OmpR expression, which in turn leads to a reduction in OmpC or OmpF porins, ultimately increasing resistance to β-lactams and fluoroquinolones. 47
EnvZ/OmpR, a critical two-component regulatory system in S. Enteritidis, not only modulates antibiotic resistance but also senses and responds to environmental cues. 48 In this case, the mutation in envZ appeared to affect colony morphology, biofilm formation, adhesion capacity, motility, and acid resistance. In terms of colony morphology, the two showed differences at 37°C, while there was almost no difference at 25°C. Sal B formed rougher and drier colonies at 37°C than Sal D. Also, Sal B had more biofilm-forming ability, motility, and adhesion than Sal D. These findings were consistent with previous studies.30,49–51 In addition, bacterial differences in adhesion may also be related to mutations in siiE, which is secreted by a T1SS that is also encoded by genes within SPI-4.52,53 These results suggest that S. Enteritidis in different environments (blood vs. pleural fluid) may further affect the expression of related proteins to adapt to the environment through mutations in related genes. These genes were closely related to bacterial signal transduction and phenotypic traits. Future studies could further explore the roles of these genes in the pathogenic mechanisms of S. Enteritidis, providing insights for the development of targeted therapeutic strategies.
Overall, this study reveals the adaptive changes and biological characteristic differences of S. Enteritidis in different environments, within the same host, emphasizing the importance of genomic analysis in elucidating bacterial pathogenesis. These findings provide a new perspective for the clinical diagnosis and treatment of S. Enteritidis infections, as well as laying a foundation for future research and public health strategies. Future investigations need to further explore how various environmental factors influence S. Enteritidis adaptability and host interactions to develop targeted intervention.
Footnotes
Availability of Data and Materials
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Authors’ Contributions
X.C. contributed to the study’s conception and design, conducted experiments, analyzed and interpreted the data, elaborated the figures, and drafted the article. X.C. contributed to conducting experiments, acquiring data, analyzing the data, and drafting the article. L.M. contributed to the data acquisition, and statistical analysis, and revised the drafted article. J.W., Y.X., Y.L., S.M., and J.L. contributed to collecting isolates and conducting experiments. W.S. contributed to the conception and design of the study, revised the drafted article, and supervised the whole process. All contributors approved the final version of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.