
Abstract
Aeromonas dhakensis is prevalent in aquatic environments in Taiwan and known for its notable antimicrobial resistance. However, comprehensive pan-genomic studies for this species in Taiwan are limited. This study analyzed 28 clinical A. dhakensis isolates using single-molecule real-time sequencing technology, coupled with diverse databases, to elucidate the whole genomes. The focus was on phylogenetic relatedness, antimicrobial resistance genes, and mobile genetic elements. Genomic analysis and multilocus sequence typing were utilized to identify A. dhakensis strains of heterogeneous origins. The detection of various β-lactamase genes (blacphA, blaimiH, blaAQU, blaOXA, blaTEM-1, blaTRU-1, and blaVEB) in clinical A. dhakensis isolates raises concern, especially considering the use of carbapenems and third-generation cephalosporins in patients with severe infections. Notably, most A. dhakensis strains carry chromosome-encoded β-lactamases, including AmpC, metallo-β-lactamase, and oxacillinase, and were susceptible to cefepime in drug susceptibility tests. A. dhakensis strains were also susceptible to aminoglycosides, fluoroquinolones, tigecycline, and trimethoprim/sulfamethoxazole. Three of the 28 A. dhakensis isolates carried plasmids containing an array of drug resistance genes, suggesting this species is likely a recipient or donor of drug resistance genes through horizontal gene transfer. Our findings provide valuable insights into the antimicrobial resistance of A. dhakensis, highlighting the medical implications of its β-lactamase diversity and its potential role in the horizontal gene transfer of drug resistance genes.
Introduction
Aeromonas dhakensis, which was identified as a species in 2013, 1 has been isolated from humans, animals, and the environment globally. A comprehensive review of over 1,800 clinical Aeromonas isolates utilizing molecular methods has revealed that A. dhakensis ranks among the top three species responsible for clinical infections on a global scale. It is responsible for one-fifth of the overall Aeromonas clinical isolates. It is frequently encountered in diverse sources, including wounds, fecal samples, and blood. 1 A. dhakensis is also well known for its resistance to β-lactams.2,3 Several genes encoding β-lactamases located in chromosomes, such as Ambler class B metallo-β-lactamase (MBL), class C, and class D resistance genes, have been detected in this species.4–6
Over the past decade, advancements in next-generation sequencing (NGS) technologies have significantly enhanced our ability to unravel the intricacies of microbial genomes with unprecedented clarity. With the advancement of whole genome sequencing (WGS) techniques, the analysis of Aeromonas has been promoted, enhancing our understanding of this species. Taiwan is one of the important endemic areas of A. dhakensis worldwide. 7 However, WGS studies of A. dhakensis are scarce in Taiwan. In the current study, we collected 28 clinical A. dhakensis isolates sourced from various origins (blood, bile, wound, and sputum) within a medical center situated in southern Taiwan using PacBio sequencing analysis. PacBio sequencing is the third-generation sequencing (TGS) method using the single-molecule real-time (SMRT) technique, developed by Pacific BioSciences®, offering longer read lengths and faster runs with a higher accuracy rate than other NGS techniques. 8 In the present study, we analyzed antimicrobial resistance genes (ARGs) and mobile genetic elements (MGEs) for clinical A. dhakensis isolates and aimed to decipher a clearer picture of genomes in aspects of antimicrobial resistance.
Materials and Methods
rpoD and multilocus sequence typing
Aeromonas identified by MALDI-TOF MS V2.0 (bioMérieux, Marcy-l’Étoile, France) at the study center, collected from January 2020 to December 2021 and stored at −70°C, underwent analysis. Species identification of each isolate was determined based on the partial sequences of rpoD. 9 To elucidate the genetic diversity and evolutionary relationships of A. dhakensis isolates, multilocus sequence typing (MLST) was performed, as described by Martino et al., using six housekeeping genes: gyrB, groL, gltA, metG, ppsA, and recA. 10 Briefly, DNA was extracted from each isolate, followed by PCR amplification of the target genes. The resulting PCR products were sequenced, and the sequences were analyzed using the PubMLST database for Aeromonas species to assign alleles and sequence types (STs). The requirement for informed consent was waived by the Institutional Review Board (A-ER-113-111).
Whole genomic analysis by TGS
The genomes of 28 strains from various individuals and infection sites (blood: 9, bile: 6, wound: 11, and sputum: 2) were sequenced using the PacBio Sequel IIe system (Pacific Biosciences Inc., Menlo Park, CA, USA) at the study center. Genomic DNA was extracted and purified using the QIAGEN MagAttract HMW DNA kit (Qiagen, Germany) to obtain high-quality DNA samples. The library construction and sequencing preparation workflow follow the manufacturer’s recommended protocol (PN: 101-696-100, version 8). Sequencing results are subsequently analyzed with genome assembly in the pipeline within the SMRT Link software version 11 (Pacific Biosciences Inc., Menlo Park, CA, USA). The genome sequences of the 28 A. dhakensis strains were submitted to the National Center for Biotechnology Information (NCBI) database under BioProject accession number PRJNA1092388 and genome accession numbers SRX26019577–SRX26019604. All the sequenced datasets were deposited into the NCBI BioSample database (Supplementary Table S1).
Construction of phylogenetic tree
The phylogenomic trees comprising 28 clinical isolates of A. dhakensis, alongside 263 reference genomes of Aeromonas, and Bradyrhizobium diazoefficiens USDA 110 as the outgroup were constructed using GToTree. This was accomplished utilizing 74 bacterial single-copy marker genes identified by the hidden Markov model, and the trees were visualized using iTOL (https://itol.embl.de), sourced from NCBI’s GenBank sequence database.
ARG analysis
The genomes were queried against the Comprehensive Antibiotic Resistance Database (CARD v3.1.4) 11 for the in silico prediction of the possible ARG contained in the genomes. The Resistance Gene Identifier (RGI v5.2.0; default parameters of RGI bwt mode) was employed for gene analysis with CARD. The search criteria were configured for “perfect and strict hits only.”
MGEs investigation
To further investigate the MGE within the whole genome of the isolates and categorize them, MGE was identified using mobileOG-db (Data Version: Beatrix 1.6 v1). 12 Reads were annotated using DIAMOND blastx (v2.015) within the following criteria: e-value < 1e-10, identity percentage >90%, query coverage >90%, and a maximum of two query hits. 13
Drug susceptibility test
All the Aeromonas isolates were stored at −80°C until use. The antimicrobial susceptibility of each A. dhakensis isolate was determined by Sensititre™ Gram Negative GN7F AST plate with an automated plate reading system, Sensititre™ Vizion™ Digital MIC Viewing System (Thermo Fisher Scientific), and the interpretative criteria followed the Clinical and Laboratory Standards Institute (CLSI) recommendations. The breakpoints applied for the tested drugs according to the CLSI M45 A2, 2015, 14 and the CLSI M45 A2, 2010. 15 There are no established breakpoints for minocycline, tigecycline, ceftolozane/tazobactam, ceftazidime/avibactam, and nitrofurantoin.
Results
The TGS of A. dhakensis isolates
A total of 28 Aeromonas isolates from various infection sites (blood: 9, pus: 2, wound: 11, and bile: 6) collected between 2020 and 2021 were identified as A. dhakensis through rpoD sequencing. The results of MLST typing for each isolate are presented in Supplementary Table S2. The findings indicate that there were 28 A. dhakensis isolates distributed across 27 distinct MLST types. We constructed a phylogenomic tree of 28 A. dhakensis isolates along with 263 Aeromonas reference genomes and Bradyrhizobium diazoefficiens USDA 110 as the outgroup sourced from NCBI’s GenBank sequence database. The tree was constructed using 74 bacterial single-copy marker genes. The result showed that the 28 A. dhakensis isolates from different individuals were located in the same cluster, with genetic distance near A. hydrophila, and far away from A. caviae and A. veronii (Fig. 1). Identification of the A. dhakensis using TGS result was matched to that determined by single rpoD gene sequencing. All 28 A. dhakensis isolates were sequenced using the PacBio method, resulting in high-quality sequences. The genomes were assembled into single contigs ranging from 4.4 to 5.1 Mb in size, and plasmids of varying sizes from 14 kb to 367 kb were found in three isolates.
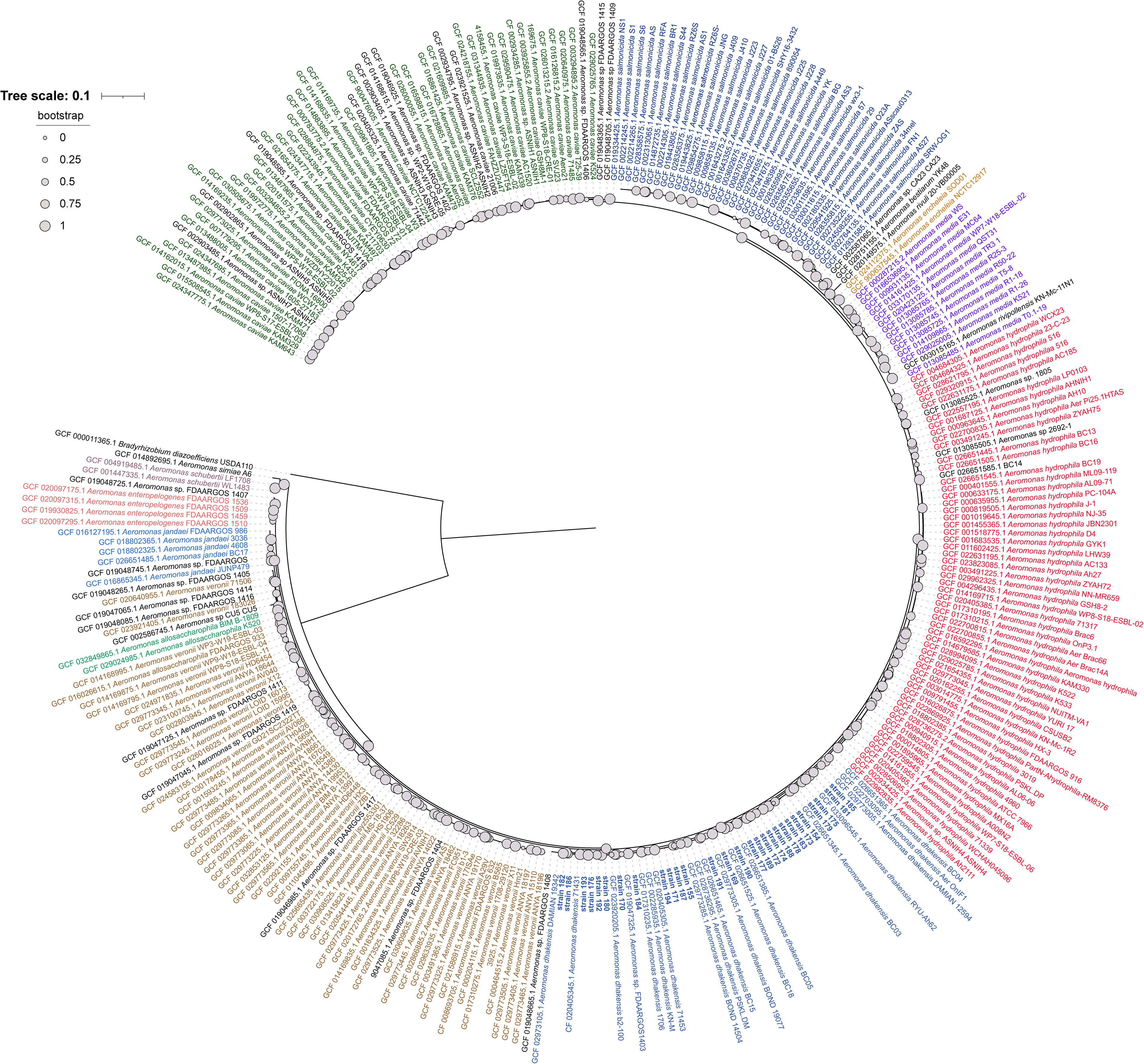
Phylogenomic trees of the 28 Aeromonas dhakensis strains (▲), along with 263 Aeromonas reference genomes, and Bradyrhizobium diazoefficiens USDA 110 as the outgroup sourced from NCBI’s GenBank sequence database. The tree was constructed using GToTree with 74 bacterial single-copy marker genes identified by a hidden Markov model (HMM) and visualized using iTOL (https://itol.embl.de). The scale bar represents the number of nucleotide substitutions per sequence site.
ARGs in A. dhakensis isolates
The genomic analysis annotated 33 ARGs in 13 drug classes using the CARD v3.1.4 for these A. dhakensis strains (Supplementary Table S3). The ARGs responsible for resistance against aminoglycosides, carbapenems, penems, cephalosporins, diaminopyrimidines, elfamycins, fluoroquinolones, macrolides, phenicols, sulfonamides, and tetracyclines were prevalent among these 28 strains (Supplementary Table S4).
We further elucidated the distribution of various ARGs among 28 A. dhakensis isolates (Fig. 2). Genomic locations of ARGs were analyzed based on RGI/CARD database annotations first. In addition, we employed PlasFlow (v1.1), a sophisticated machine learning-based tool, to analyze the entire genome. This tool is designed to predict whether genomic fragments are of chromosomal or plasmid origin. We also utilized PlasmidFinder 2.1, 16 a highly specific tool for identifying known plasmid replicons. The results showed that all the ARGs were located on chromosomes. However, three isolates (strains 155, 179, and 187) were found to carry plasmids that encode an array of resistance genes, including MBL blaImi and blacphA2, oxacillinase blaOXA-726, Amp-C blaTRU-1, tet(E), adeF, rsmA, EF-Tu, and adeF (Table 1). Three genes encoding carbapenemases, including blaImi, blaOXA, and blacphA2, were detected. Twenty-seven of them harbored at least one of three genes encoding carbapenemases, including blaImi, blaOXA, and blacphA2. Interestingly, one isolate (strain 176) exhibited a carbapenem-resistance phenotype, characterized by decreased susceptibility to ertapenem, imipenem, and meropenem, despite the absence of detectable genes encoding carbapenemases. This phenomenon indicates that the isolate possesses a carbapenem resistance mechanism other than these carbapenemases and is worthy of further investigation.

Heatmap illustrating the distribution and genomic context of antimicrobial resistance genes (ARGs) in 28 Aeromonas dhakensis strains. The color intensity represents ARG copy numbers. All ARGs shown in this figure are chromosomally located based on complete genome analysis.
Analysis of Antimicrobial Resistance Genes on Plasmids from Three Aeromonas dhakensis Isolates
Chromosomal inducible Ambler class B MBL genes, blacphA-like, and blaImiH genes were detected in 14 and 12 isolates, respectively. blaAQU-like genes, which encode class C AmpC β-lactamases, were detected in 14 A. dhakensis isolates. Twenty-seven isolates possessed genes encoding class D β-lactamase, OXA-726, with one of them carrying genes for both OXA-726 and OXA-10 β-lactamases. Tetracycline resistance genes, including tet (A), tet (E), and tet (C), were discovered in 10 isolates, and sulfonamide resistance genes, including sul1 and sul2, were detected in 4 isolates. Chromosome-associated aminoglycoside resistance genes, including APH (3″)-Ib, APH (6)-Id, AAC-Ib-cr6, ANT(2″)-Ia, AAC(6′)-Id, APH(4)-Ia, and AAC(3)-IV, were distributed in three isolates.
Antimicrobial susceptibility of A. dhakensis isolates
Antimicrobial susceptibility test results are demonstrated in Table 2. All the isolates showed resistance to ampicillin/sulbactam and cefazolin. One-third of the strains (10 out of 28) were resistant to tetracycline. Tetracycline resistance genes (tet) were present in all 10 isolates. Specifically, tet(E) was detected in seven isolates, tet(A) in two isolates, and one isolate contained both tet(E) and tet(C). Tet genes were not detected in any strains without tetracycline resistance. One-third of the isolates were resistant to piperacillin/tazobactam. Susceptibility to third-generation cephalosporins, specifically ceftriaxone and ceftazidime, was found to be decreased in 53.6% and 67.9% of the isolates, respectively. The susceptibility to the fourth-generation cephalosporin cefepime remained at 82.1%. The A. dhakensis isolates showed substantial resistance to carbapenem, being less susceptible to imipenem (60.7%), followed by ertapenem (82.1%), doripenem (82.1%), and meropenem (85.7%). The isolates demonstrated substantial susceptibility to amikacin (100%), gentamicin (96.4%), aztreonam (82.1%), ciprofloxacin (82.1%), and trimethoprim/sulfamethoxazole (85.7%). Sixty-four percent of A. dhakensis isolates were susceptible to tetracycline. Although there are no established criteria, all the isolates exhibited high susceptibility to tigecycline, with minimum inhibitory concentration (MIC) levels below 2 μg/mL. Ceftolozane/tazobactam and ceftazidime/avibactam are advanced-generation cephalosporins combined with β-lactamase inhibitors, which are active against drug-resistant gram-negative bacteria. 17 Despite the absence of established drug susceptibility standards for Aeromonas species, the majority of strains (24 out of 28) exhibited an MIC level of ≤2 μg/mL for ceftazidime/avibactam, and 22 of them had an MIC level of <2 μg/mL for ceftolozane/tazobactam.
Drug Susceptibility Determined Minimal Inhibitory Concentration (MIC, μg/mL) Level for the 28 Aeromonas dhakensis Isolates
Strains 155, 179, and 187 carried plasmid-associated antimicrobial resistance genes.
AMK, amikacin; Amp, ampicillin; Amp/sul, ampicillin/sulbactam 2:1 ratio; Azt, aztreonam; CAZ, ceftazidime; CEF, cefepime; CEZ, cefazolin; Cez/avi, ceftazidime/avibactam; Cez/Taz, ceftolozane/tazobactam; CIP, ciprofloxacin; CRX, ceftriaxone; dori, doripenem; Ert, ertapenem; GM, gentamicin; Imi, imipenem; Mer, meropenem; MIN, minocycline; NC, no criteria; Nit, nitrofurantoin; Pip/tazo, piperacillin/tazobactam; TET, tetracycline; TIG, tigecycline; TRM/SMX, trimethoprim/sulfamethoxazole.
Mobile genomic elements in A. dhakensis isolates
To identify the presence of MGEs among the A. dhakensis isolates, the TGS data were analyzed using the mobileOG-db database comprising over 700,000 deduplicated sequences encompassing 5 major mobileOG categories and more than 50 minor categories. 12 A total of 3,966 genes, distributed across 34 minor categories within the 5 major MGE categories, were identified in the 28 A. dhakensis isolates. The counts of genes varied among the categories: phage (620), transfer (1,197), replication/recombination/repair (1,621), stability/transfer/defense (150), and integration/excision (378; Fig. 3). Each strain possessed a significant number of MGE genes, ranging from 104 to 258 (Fig. 4).

The 28 Aeromonas dhakensis strains are distributed within the 5 major mobile OG categories. Each major category contains various minor categories. Bubble size denotes the number of gene counts. NA, category not classified.
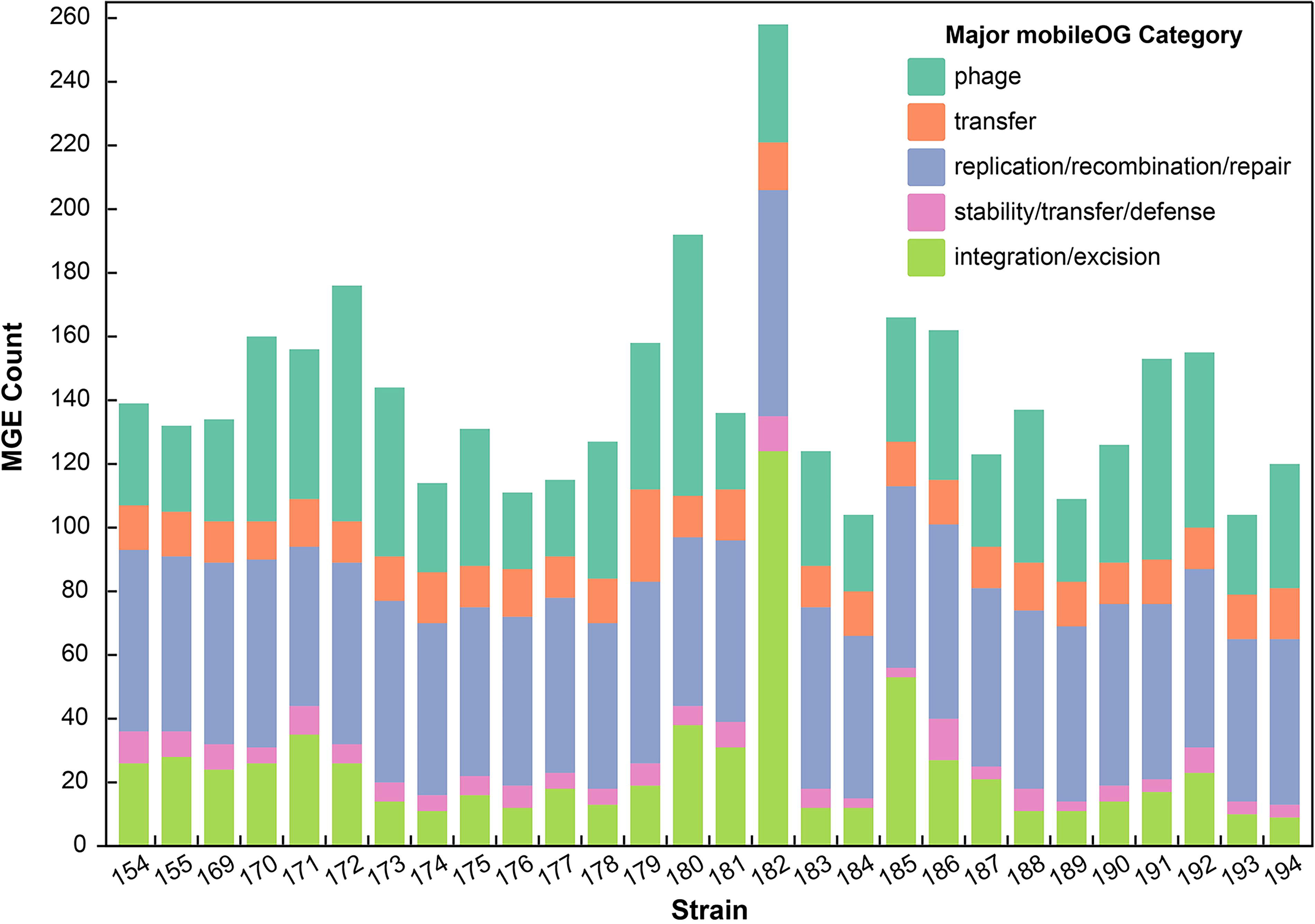
The mobile genetic element (MGE) counts within 5 major categories: phage, transfer, replication/recombination/repair, stability/transfer/defense, and integration/excision in 28 Aeromonas dhakensis strains.
Discussion
Utilizing TGS analysis, clinical A. dhakensis isolates were found to be polyclonal in nature and harbor various types of β-lactamase genes, including blacphA, blaImiH, blaAQU, blaOXA, blaTEM-1, blaTRU-1, and blaVEB. Previous studies revealed that A. dhakensis harbors genes encoding AmpC (blaAQU), MBL (blacphA and blaImiH), and oxacillinase (blaOXA).18–21 The widespread presence of various types of β-lactamase genes would preclude the selection of first-generation cephalosporins, second-generation cephalosporins, and carbapenems as effective therapeutic agents against A. dhakensis infection. However, it has been reported that there is often a lack of concordance between the actual resistance profile and the presence of various types of β-lactamase genes in Aeromonas species.22–24
Of the 28 A. dhakensis isolates, 27 carried genes encoding AmpC β-lactamases. These genes were categorized into two groups: blaTRU-1 and blaAQU. Among the 27 isolates positive for AmpC β-lactamase genes, 14 A. dhakensis isolates harbored blaAQU-related genes (1 blaAQU-1, 7 with blaAQU-2, and 6 with blaAQU-3), whereas the other 13 isolates possessed the blaTRU-1 gene. None of the isolates carried both genes. The blaAQU-related genes have been reported from A. dhakensis and are unique to this species. 25 The blaTRU-1 gene that belongs to the AmpC family was reported from A. enteropelogenes 26 but rarely reported in A. dhakensis. The wild-type blaAQU gene confers resistance against cephamycins, third-generation cephalosporins, and β-lactam inhibitors such as clavulanate, tazobactam, and sulbactam. However, it cannot hydrolyze penicillins or carbapenems. 25 The constitutive production of TRU-1 β-lactamase may be inducible in A. enteropelogenes, leading to resistance against third-generation cephalosporins while maintaining susceptibility to cefepime, aztreonam, and imipenem. 26
blaOXA-associated genes have been discovered in Aeromonas isolates from diverse resources.6,24,27,28 Among 28 A. dhakensis isolates, 27 carried the blaOXA-726 gene, whereas one possessed the blaOXA-10 gene. OXA-726-encoding A. dhakensis Igbk (ST 1171) was isolated from an edible snail traded in Nigeria, 29 suggesting the widespread presence of this gene worldwide and its potential threat to humans. The blaOXA-726 gene detected in most of the A. dhakensis isolates is genetically close to blaOXA-830, blaOXA-724, blaOXA-725, and blaOXA-12. 27 Most of our isolates exhibiting carbapenem resistance phenotypically carried an additional gene encoding MBL carbapenemase, alongside blaOXA-726.
The extended-spectrum-lactamase (ESBL) gene, blaVEB-1, belongs to class A β-lactamase and was discovered in one A. dhakensis isolate. A variety of ESBL genes, including blaVEB-1a, blaSHV-12, blaPER-1, blaPER-6, blaTLA-2, and blaGES-7 detected in Aeromonas isolates from the Seine River, France, was reported, 30 indicating that the Aeromonas genus is a reservoir for these ESBL genes. The blaVEB-1 has been reported prevalent among Aeromonas species resistant to or with reduced susceptibility to ceftazidime and inhibited by the β-lactamase inhibitor clavulanic acid.30–32 Taken together, the presence of AmpC, MBL, and OXA in A. dhakensis makes cefepime a reasonable choice for severe A. dhakensis infections unless the causative isolates are EBSL producers.
Ten A. dhakensis isolates demonstrated reduced susceptibility to tetracycline with MIC levels >8 mg/dL. Despite the absence of an established breakpoint for tigecycline, all 28 A. dhakensis isolates showed good susceptibility with MIC levels <1 mg/dL. Similar results were observed in other studies testing Aeromonas isolates,33,34 suggesting that tigecycline is one of the drug choices for treating Aeromonas infections. All of these isolates possessed tet genes, which encode drug efflux pumps (2 with tet(A), 7 with tet(E), and 1 with tet(E) and tet(C)) based on the TGS analysis using CARD v3.1.4. Tet(C) was detected in plasmid RA53 from A. salmonicida. 35 To our knowledge, this is the first finding of tet(C) existing among A. dhakensis strains.
In our study, we identified 378 gene counts in the integration/excision category of the mobileOG-db database, which are linked to transposon function, across 28 A. dhakensis isolates. However, no ARGs were discovered on transposons. Using the TGS method, we identified 3 out of 28 A. dhakensis isolates that carried plasmids. Important ARGs, such as AmpC (blaTRU-1), MBL (blaImiH and blacphA2), oxacillinase (blaOXA), and tet(E), were located on these plasmids (Table 1). These three strains carrying plasmids demonstrated resistance to carbapenems and decreased susceptibility to the third-generation cephalosporins and the fourth-generation cephalosporins (Table 2). A detailed sequence comparison of the three plasmids identified in strains 155, 179, and 187 was performed (Supplementary Table S5). No MGEs were detected in these plasmids using mobileOG-db analysis. Sequence comparison between these three plasmids showed limited overall similarity beyond the ARG regions, indicating they likely represent distinct plasmid types rather than variants of a common plasmid. This observation, combined with the absence of MGEs on the plasmids themselves, suggests that the distribution of ARGs among A. dhakensis strains might involve more complex mechanisms than direct plasmid sharing.
Conclusion
Leveraging the SMRT sequencing technique of TGS, alongside diverse databases, can accelerate the comprehension of the genomes of pathogenic strains. With the aid of TGS, we identified drug resistance genes blaTRU-1, blaOXA-726 blaVEB-1, and tet(C), which were first reported in A. dhakensis isolates from Taiwan. The presence of various drug-resistance genes in clinical A. dhakensis suggests that this species serves as a potential vector for transmitting ARGs through horizontal gene transfer in different environments, warranting continuous surveillance.
Footnotes
Acknowledgments
We extend our gratitude to the Diagnostic Microbiology and Antimicrobial Resistance Laboratory at National Cheng Kung University Hospital for their technical assistance in bacterial culture and drug susceptibility testing for the isolates. We specially thank Dr. Guann-An Chen of Blossom Biotechnologies Inc., Taiwan, for his invaluable assistance in designing the WGS workflow and Yen-Ling Peng of Blossom Biotechnologies Inc., Taiwan, for her contributions to big data curation. We thank the National Core Facility for Biopharmaceuticals and the National Center for High-performance Computing of National Applied Research Laboratories of Taiwan for providing computational resources and storage resources.
Authors’ Contributions
Y.-W.C.: Project administration and funding acquisition. Y.-C.S.: Investigation, data curation, and writing—review and editing. W.-Y.C.: Formal analysis, methodology, and writing—review and editing. J.-H.W.: Conceptualization and supervision. J.-W.C.: Conceptualization and supervision. S.-L.S.: Investigation and methodology. C.-S.C.: Resources and supervision. P.-F.T.: Resources and methodology. W.-C.K.: Supervision, funding acquisition, and resources. P.-L.C.: Conceptualization, funding acquisition, project administration, and writing—review and editing.
Data Availability Statement
The data that support the findings of this study are available upon reasonable request from the corresponding author.
Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article. All the authors declare that they have no conflict of interest.
Funding Information
The study received support from research grants provided by the Ministry of Science and Technology, Taiwan (MOST 111-2314-B-006-048; 111-2221-E-006-022-MY3; 110-2923-E-006-004-MY3; NSTC 113-2321-B-006-007; NSTC 113-2314-B-006-045) and National Cheng Kung University Hospital (NCKUH-11109029; NCKUH-11301006; NCKUH-11401002).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.