
Abstract
Objective:
Aminoguanidine, which inhibits the formation of advanced glycosylation end products, can restore the ability of endothelial cells to align and elongate in response to shear stress when that ability is lost during culture in high glucose conditions. This study tests whether aminoguanidine can also restore migratory ability of endothelial cells and whether pyridoxine, a stable form of vitamin B6, can restore migratory ability and ability to align and elongate in response to shear.
Methods:
Human aortic endothelial cells were cultured in normal glucose (5.5 mM), 17.5 mM glucose, and 30.5 mM glucose in the presence or absence of 5 mM aminoguanidine or varying concentrations of pyridoxine (10–1,000 mg/L). Assay of percent closure of a scrape wound after 24 h quantified migratory ability, and alignment and elongation under flow at 10 dynes/cm2 quantified response to shear stress.
Results:
Aminoguanidine (5 mM) fully restores and pyridoxine (100 mg/L, 0.6 μM) partially restores migratory ability of cells cultured in 30.5 mM glucose. Pyridoxine (100 mg/L) fully restores the migratory ability of cells cultured in 17.5 mM glucose. Pyridoxine (100 mg/L) fully restores endothelial cell alignment and elongation and response to shear stress at 30.5 mM glucose.
Conclusions:
Pyridoxine, at dosages known to be safe from previous studies (<250 mg/day) can restore migratory ability and shear stress response to endothelial cells cultured in high-glucose conditions. This indicates that pyridoxine is a potential candidate for treatment of diabetic ulcers and atherosclerosis in diabetes due to the link between these pathologies and endothelial dysfunction in diabetes.
Introduction
Brower et al. then sought to determine the mechanism by which HG hinders these responses. They demonstrated that treatment of the cells with aminoguanidine, an inhibitor of advanced glycation end products (AGEs), blocked the effect of HG on the shear stress response allowing human aortic endothelial cells (HAECs) to align and elongate as if they were in normal media (5.5 mM glucose). Because aminoguanidine restored the ability of HAECs to respond to shear stress, we hypothesize that blocking formation of AGEs with pyridoxine will also restore HAEC migration to normal levels in HG conditions.
We also hypothesize that pyridoxine, a stable form of vitamin B6, will similarly restore endothelial shear stress response and ability to align and elongate. If pyridoxine can do so, it would be a potentially useful method to treat diabetic patients for the purposes of improving wound healing and prevention of atherosclerosis. While aminoguanidine has been highly successful at reducing AGEs in experimentally induced HG conditions, few studies have attempted to translate this success into clinical success. Concerns regarding long-term toxicity of aminoguanidine have largely halted its use as a clinical solution to accumulation of AGEs. 12 Due to these complications it is necessary to find a more biocompatible inhibitor to the formation of AGEs. Pyridoxine, the catalytically inactive form of vitamin B6, is readily converted intracellularly to the active compounds pyridoxamine and pyridoxal phosphate. 13,14 Pyridoxamine is a potent inhibitor of AGE formation and has been shown to reduce nephropathy and retinopathy in diabetic rat models. 15 –17
In this study, we first test whether aminoguanidine can prevent HG inhibition of HAEC migration in a scrape wound experiment. We then perform the same test with pyridoxine to determine whether pyridoxine can restore migratory function of HAECs in HG media. We then measure a dose–response curve of pyridoxine dose versus wound closure at 24 h. Last, we test whether pyridoxine restores response to shear stress (cell elongation and alignment in the direction of flow) in the presence of HG. Our results indicate that pyridoxine may have the potential to be a clinically relevant method of preventing endothelial cell dysfunction in diabetic patients.
Materials and Methods
Cells and reagents
HAECs were cultured using Medium 200 supplemented with low-serum growth supplement and penicillin/streptomycin (Cascade Biologics) and cultured either in a 12 or 24-well culture plate (migration assay) or on a microscope slide (flow experiment). For HG experimental cultures, when cells reached 70% confluence, the medium was supplemented with an additional 12 mM or 25 mM
For experiments including aminoguanidine, medium was supplemented with 5 mM aminoguanidine when cells reached 70% confluence. For experiments with pyridoxine, a range of concentrations of pyridoxine (10, 25, 50, 75, 100, 150, 200, 300, 500, 600, 700, 800, 1,000 mg/L equivalent molar concentrations: 5.9 × 10−5, 1.5 × 10−4, 3.0 × 10−4, 4.4 × 10−4, 5.9 × 10−4, 8.9 × 10−4, 1.2 × 10−3, 1.8 × 10−3, 3.0 × 10−3, 3.6 × 10−3, 4.1 × 10−3, 4.7 × 10−3, 5.9 × 10−3 M) was added to the endothelial cells at the same time as the increase in glucose for the dose–response curve, and 100 mg/L was added to the cells for other experiments with pyridoxine. Culture medium in all samples was changed every 24 h.
Scrape-wound repair assay
HAECs were seeded (1 × 104 cells/cm2 in 24-well plates for aminoguanidine, 4 × 104 cells/well in 12-well plates for pyridoxine) and grown to 70% confluence using the methods described above. When 70% confluent, the medium was modified as described above for the experimental and control groups. Upon reaching confluence (approximately 48 h), scrape wounds were created using the sharp tip of a sterilized wood stick (aminoguanidine experiments) or a 1,000-μL pipette tip (pyridoxine experiments). Wounded monolayers were photographed immediately using a 5× microscope objective on an inverted microscope (Leica Microsystems). They then were maintained in their respective treatment conditions for 24 h. For the aminoguanidine experiments, after 24 h, cells were fixed with 3.7% formaldehyde, stained with 0.25% Toluidine Blue, and rinsed three times with phosphate-buffered saline (PBS). In both experiments, cells were again photographed using a 5× objective. For the aminoguanidine experiments, the area covered by cells after 24 h was compared to the sample's respective initial wound area using a custom program written in the Matlab programming language. The pyridoxine experiments were analyzed with a simpler method in which the width of the area in which cell coverage is less than 50% (w) is used to determine a percent closure: [(w 0 − w)/w 0]*100, where w 0 is the initial width of the scrape wound.
Flow experiments
NG and HG cells were exposed to 10 dynes/cm2 of fluid shear stress as described by Brower et al. 11 Briefly, individual microscope slides were mounted into a recirculating parallel plate flow chamber device primed with either normal or HG cell culture medium. Temperature of the medium was maintained at 37°C by submerging a medium reservoir, which acts as an upstream pulse dampener, in a 37°C water bath. pH was maintained at 7.4 using 10 mM HEPES (Sigma) in the flowing medium. Flow was driven by a biphasic peristaltic pump (Masterflex L/S, Cole-Parmer). An additional downstream pulse dampener was used in the flow loop to minimize pulsatile effects and ensure steady-state laminar flow. Shear stress at the microscope slide was calculated as τw = 6 μQ/h 2 b, where τw is the shear stress at the slide, μ is the fluid viscosity (8.5 × 10−4 Pa · s), Q is the volumetric flow rate (2.0 × 10−7 m3/s), h is the flow chamber height (2.7 × 10−4 m), and b is the flow chamber width (1.8 × 10−2 m). Cells were exposed to flow for 12 h. Static cultures were kept for all treatment groups to ensure the treatment conditions did not noticeably affect the confluence or overall morphology of the cell cultures.
Slides were removed from the flow chamber, rinsed in prewarmed PBS, fixed for 10 min in 4% formaldehyde in PBS, and rinsed again with PBS. Images were then taken on an inverted microscope with a 10× phase objective. Images were taken on random optical fields away from the cover glass edge. A custom program was written to analyze the images in Matlab. For each image, the user was prompted to manually trace the periphery of whole cells in the image. The length of major and minor axes of each cell in these images was calculated. The aspect ratio (major/minor axis) of each whole cell and the angle difference between the major axis and the flow direction were calculated to give measures of cellular elongation and orientation, respectively, in response to fluid shear stress. Features were measured for 30–60 cells from 10 images in triplicate flow experiments for each treatment group.
Statistical analyses
Migration assay results were obtained by taking three to five images of each well and averaging the results of the images. Experiments were performed in triplicate and data is expressed as mean ± standard error of the mean (SEM). Population means were compared using analysis of variance (ANOVA), and groups were compared using t-tests. Flow data were obtained by performing three experiments for each condition. For each sample, 10 images were taken and analyzed for cell alignment and elongation. Populations were compared using ANOVA and post hoc comparisons between groups using a t-test. Data are presented as mean ± SEM.
Results
Previous work by other investigators has shown that cell migration is inhibited by HG media. 8 –10 We reproduce this result, shown in Fig. 1. HG impairs migration into the wound area relative to control cells (0.50 ± 0.14 times that of control, P < 0.05 vs. NG). We then test whether addition of 5 mM aminoguanidine can restore migratory function to cells in HG. After 24 h, cells in HG + aminoguanidine have similar migratory ability as cells in NG (0.92 ± 0.12 times control, P ∼ 0.67 vs. NG).
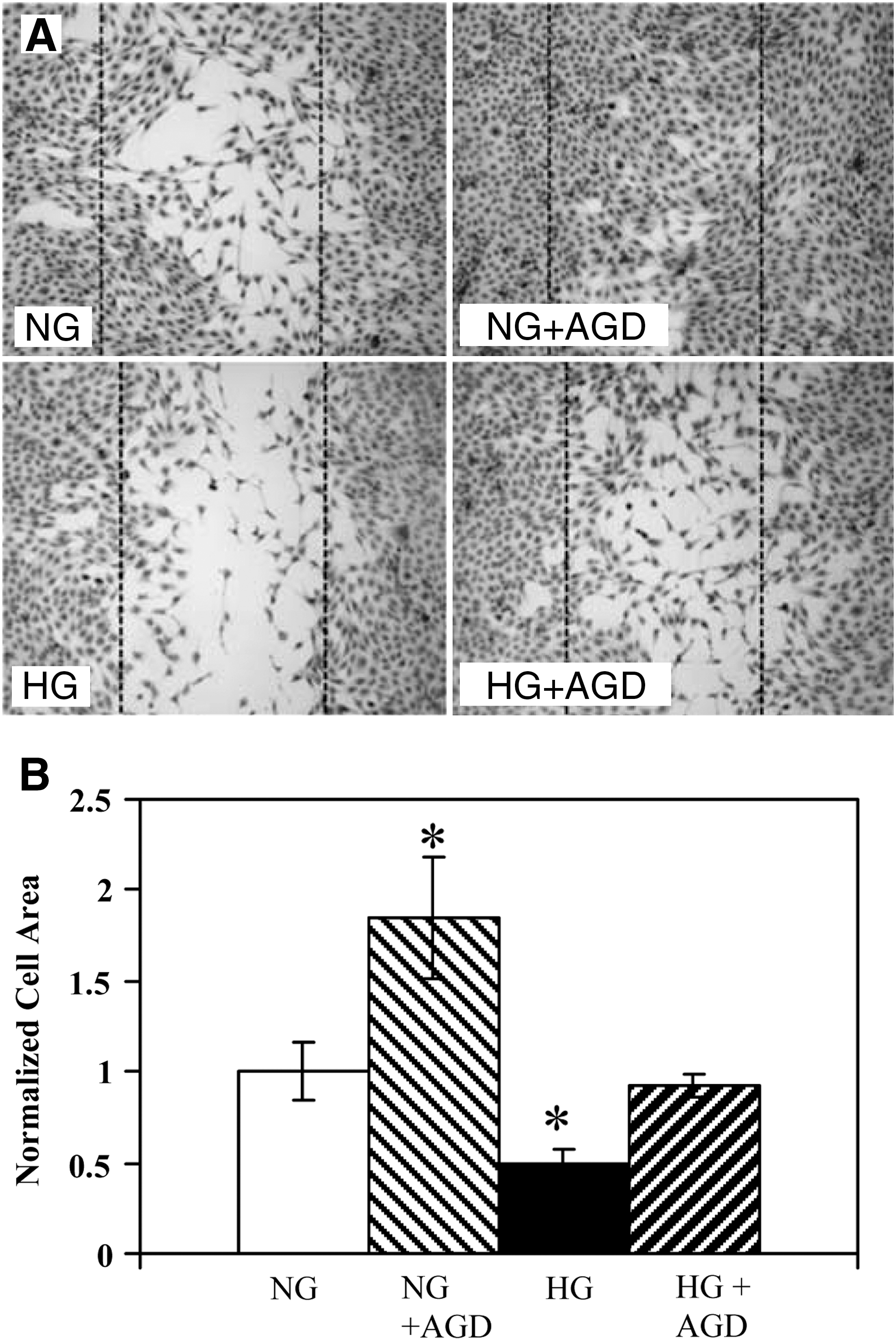
Effects of high glucose and aminoguanidine on the migration of HAECs into an induced wound area.
We then tested whether pyridoxine would similarly restore migratory function to cells in HG. Figure 2, A–F, shows representative images of HAECs under each experimental condition. Images at t = 0 and 24 h time points are shown for 5.5 mM glucose (Fig. 2A, B), 30.5 mM glucose (Fig. 2G, H), and 30.5 mM glucose with 100 mg/L pyridoxine (Fig. 2I, J). Quantitative analysis of the percent closure of the wound after 24 h showed that HAECs grown in NG achieved 82.0 ± 2.8% closure. Cells in HG only achieved 48.8% ± 3.1% closure (P < 1 × 10−6 vs. NG). Migration was significantly restored upon the addition of 100 mg/L pyridoxine. Wound closure increased to 62.7% ± 7.6% (P < 0.05 vs. HG); however, treatment with pyridoxine did not cause the cells to fully recover their normal migratory function (P < 0.0005 vs. NG).
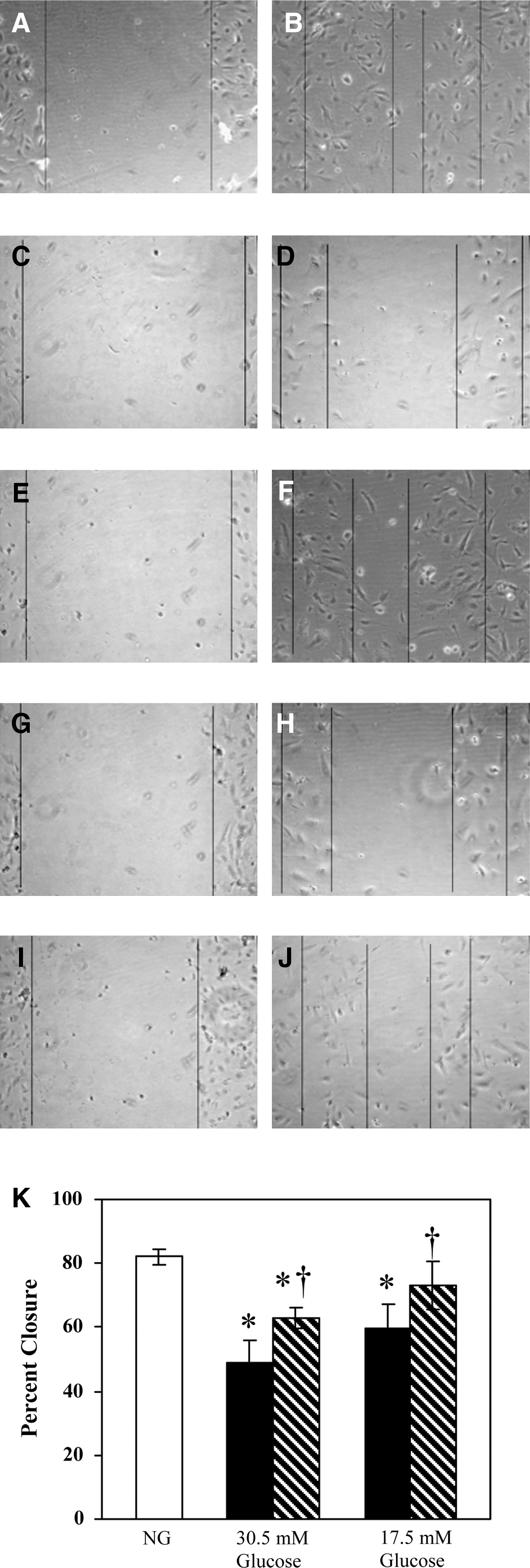
Ability of pyridoxine to restore migration of human aortic endothelial cells
To determine whether the treatment with pyridoxine could fully restore HAEC migration in conditions of less severely increased glucose concentrations, the migration assay was performed in the presence of 17.5 mM glucose. Figure 2, C and D, shows representative images of cells in 17.5 mM glucose without pyridoxine, and Fig. 2, E and F, shows images of HAECs in 17.5 mM glucose + 100 mg/L pyridoxine. Figure 2K quantifies the results showing that cells in 17.5 mM glucose migrate significantly less than cells in NG (59.5% ± 6.1%, P < 0.0001 vs. NG). Pyridoxine restores most of this lost migratory function (73.2% ± 7.4%, P < 0.05 vs. 17.5 mM glucose); the restored value is not statistically different from NG (P ≃ 0.06 vs. NG).
We initially used 100 mg/L pyridoxine because this is a potentially clinically viable dose. The result in Fig. 2 (showing that migratory function is not fully restored) led us to determine if, by increasing the concentration of pyridoxine, the migration of HAECs in 17.5 mM glucose could be fully restored. Figure 3 shows the dose–response of HAECs to pyridoxine in the presence of 17.5 mM glucose. Concentrations up to 10 mg/L do not significantly improve cell migration (P < 0.05 vs. 100 mg/mL). Concentrations between 25 mg/L and 500 mg/L restore migration similar to the response shown in Fig. 2K for 100 mg/L pyridoxine (ANOVA P < 0.05, no difference within 25–500 mg/L upon post hoc Tukey test). Above this concentration (600–1,000 mg/L pyridoxine), the cell migration response returns to the response seen in 17.5 mM glucose without pyridoxine (the post hoc Tukey test finds no difference with 0 mg/L pyridoxine).
HG has also been shown to effect alignment and elongation of HAECs in response to flow, and aminoguanidine has been shown to restore their function. 11 Figure 4, A–D, shows representative images of cells in static conditions in 5.5 mM glucose (Fig. 4A), in 5.5 mM glucose exposed to 10 dynes/cm2 shear stress for 12 h (Fig. 4B), in 30.5 mM glucose exposed to shear stress (Fig. 4C), and in 30.5 mM glucose with 100 mg/L pyridoxine exposed to shear stress (Fig. 4D). Quantification of these results (Fig. 4E, F) show that static cultures are randomly oriented (58.7° ± 19.3°, random orientation of 45° is within the 95% confidence interval) and mostly rounded (aspect ratio = 1.7 ± 0.5). After 12 h of flow, NG cells elongate (aspect ratio = 3.9 ± 1.1) and align (20.1° ± 13.3°) in the direction of fluid flow, but HG conditions (30.5 mM) inhibit this effect (aspect ratio, 2.7 ± 0.8, P < 1 × 10−5 vs. NG; alignment angle, 34.0° ± 14.1°, P < 0.001 vs. NG). Pyridoxine at 100 mg/L fully restores elongation (4.2 ± 0.9, P < 1 × 10−11 vs. HG) and alignment (18.2° ± 9.2°, P < 1 × 10−6 vs. HG) of cells cultured in HG supplemented with 100 mg/L pyridoxine. Cell elongation and alignment in HG + 100 mg/L pyridoxine are similar to those for NG controls (P ∼ 0.2 and P ∼ 0.5, respectively).
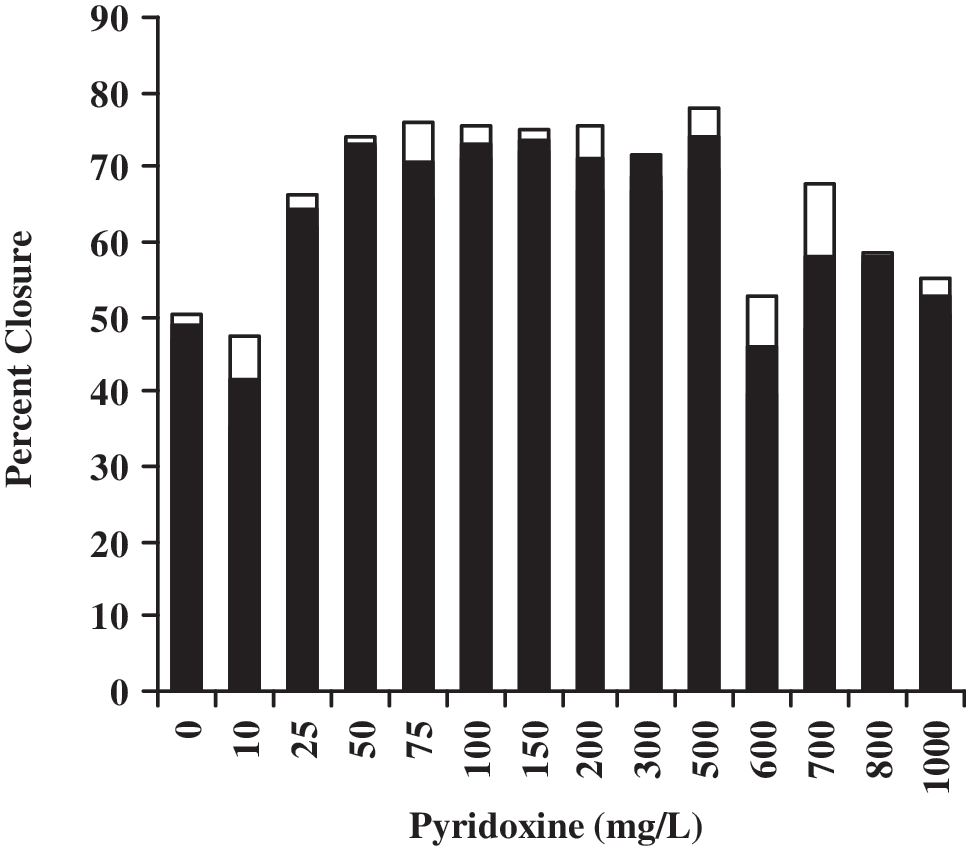
Dose–response of pyridoxine on human aortic endothelial cell (HAEC) migration in the presence of 17.5 mM glucose. Bars are mean (filled) ± standard error of the mean (SEM) (not filled). Analysis of variance (ANOVA) with post hoc Tukey test (P < 0.05) groups 0, 10, 600–1,000 separately from 25–500.

Representative images of human aortic endothelial cells (HAECs) grown under each culture condition.
Discussion
Previous work has shown that aminoguanidine and pyridoxine prevent the formation of AGEs. 15 –17 Our laboratory has previously demonstrated that aminoguanidine restores the ability of HAECs to align and elongate when cultured in HG medium. 11 In this study, we also show that pyridoxine can restore these functions when HAECs are grown in HG conditions. Furthermore, we have shown that both aminoguanidine and pyridoxine can restore migratory function of HAECs cultured in HG media to the level of function exhibited by the cells in normal (5.5 mM glucose) medium. An additional, and interesting, finding is that aminoguanidine can enhance the migratory ability of HAECs cultured in normal medium as well (P < 0.5 vs. NG).
The mechanism by which aminoguanidine and pyridoxine restore function is less well understood. A previous study in our laboratory showed that the expression of heparan sulfate proteoglycans (HSPG) was decreased in cells grown in HG medium, but HSPG was partially restored when these cells were treated with aminoguanidine. 21 HSPG is known to be a shear stress sensing mechanism in endothelial cells, so this may be the mechanism by which shear stress response is lost in cells cultured in HG and likewise is how aminoguanidine (and possibly also pyridoxine) restores the alignment and elongation responses. It is not currently known whether this mechanism or another mechanism controls the loss and subsequent restoration of migratory ability in these cells. Another possible mechanism affecting migration rate is the expression of integrins. Brower measured the expression of the α3-integrin subunit (using fluorescence activated cell sorting) and found that its expression was less when cells were exposed to high glucose and that its activity was restored to normal levels upon addition of aminoguanidine. 22 Many studies have shown a biphasic effect of cell adhesivity on migration rate. 23 Decreasing integrin expression will decrease adhesivity, and this can result in decreased migration if the cell was near the optimal adhesivity for migration.
Regardless of the mechanism, this study indicates that pyridoxine may be clinically useful as a treatment for diabetes. Pyridoxine is safe up to approximately 250 mg/day. Side effects of doses at or below 250 mg/day are minor, including mostly nausea and/or loss of appetite but also include dizziness, diarrhea, dermatitis, and flushing of the skin. 24,25 The 100 mg/L concentration used in this study is greater than this dose; total body water of approximately 0.6 L/kg body weight equates to a required dose significantly greater than 250 mg/day. However, because the effect desired is upon the endothelium lining blood vessels, it is possible that a lower dose can achieve transient concentrations in the blood close to the concentrations used in this study; and, if the endothelial cells internalize the pyridoxine, this concentration could be sustained intracellularly for sufficient duration to achieve the desired effect. Also, the dose–response curve (Fig. 3) shows that 25 mg/L can provide similar restoration of function, and this equates to a dose closer to the range of pyridoxine doses that have been used safely with minimal side effects.
There are two pathologies associated with diabetes that we believe may be ameliorated by treatment with pyridoxine—diabetic ulcers and atherosclerosis in diabetes. The migration data shown here indicate that endothelial cells in HG conditions are less able to migrate, and this may partially explain the slow rate of healing of diabetic ulcers (of course, damaged nerve response is also partially responsible due to repeated injury of these wounds). In this study, we show that pyridoxine restores migratory ability of cells in HG conditions. If some of the problem in healing of diabetic ulcers is due to inability of new blood vessels to grow into the healing wound, treatment with pyridoxine may enhance the rate of wound healing in patients with diabetes.
Other researchers have also observed that atherosclerosis, which is normally located in regions of low shear stress (such as arterial branch points), is widely distributed in the vasculature of diabetic patients. In previous reports, we have proposed that this may be due to endothelial cells being less able to sense shear stress when growing in HG conditions (e.g., diabetes). We have further proposed that plaque formation may be allowed because of impaired ability of endothelial cells to align and elongate with flow. In this study, we have shown that pyridoxine can restore the ability of endothelial cells grown in HG conditions to align and elongate with flow; therefore, there is reason to believe that pyridoxine may be a potential treatment to prevent wide-spread atherosclerosis in diabetic patients.
Although the clinical efficacy of pyridoxine must be established by clinical trials of pyridoxine in these two patient populations (patients with diabetic ulcers; prediabetic or newly diagnosed patients for prevention of atherosclerosis), the in vitro evidence shown here indicates that pyridoxine is a promising candidate for such clinical studies. Because pyridoxine is safe at the proposed dose, is very inexpensive, and is widely available, this is an exciting finding that may provide an easy, safe, and inexpensive way to improve the treatment of diabetes.
Footnotes
Acknowledgments
The authors would like to thank the Palm Foundation for funding portions of this work. M.R.C. and B.G.K. were funded by the American Heart Association and Arizona Biomedical Research Commission contract #0903.
Author Disclosure Statement
There is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.