
Abstract
Background:
Diabetic individuals experience elevated fasting glucose, glycosylated hemoglobin (HbA1c), and plasma insulin and impaired glucose tolerance. Adiponectin is a hormone inversely correlated with insulin resistance. Here we describe the activity of aloesin, an aloe chromone that increases adiponectin production and, when formulated with an aloe polysaccharide composition, improves the insulin sensitivity in db/db and diet-induced obese-diabetic mice.
Methods:
Two aloe chromones, aloesin and aloesinol, were tested in vitro for adiponectin production. Following confirmation of glucose-lowering activity in a high-fat diet (HFD)–induced mouse model, aloesin was formulated with an Aloe vera inner leaf gel powder polysaccharide preparation to yield a composition designated UP780. Efficacy of UP780 was evaluated in HDF-induced and db/db mouse models. GW1929, a synthetic peroxisome proliferator-activated receptor-γ (PPARγ) agonist, was used as a positive control.
Results:
After 3 weeks of treatment of HDF-induced mice, plasma insulin levels were decreased 37.9% and 46.7% by aloesin and aloesinol, respectively. In db/db mice, the chromone- (2% chromone:98% aloe polysaccharide) enriched UP780 aloe composition showed a 33.7% and 46.0% decrease in fasting triglyceride and plasma glucose levels after 10 weeks of oral treatment, respectively. Diabetic mice gavaged with 200 mg/kg of UP780 for 10 weeks showed a 30.3% decrease in fasting blood glucose levels and a 32.2% reduction in plasma insulin. In both animal models, UP780 showed a statistically significant improvement in blood glucose clearance.
Conclusion:
These findings indicate that UP780, a chromone-standardized, aloe-based composition, could potentially be used as a natural product option to facilitate the maintenance of healthy blood glucose levels.
Introduction
In the last decade, advances in pursuit of new molecular targets to treat T2DM resulted in the development of new classes of blood glucose–lowering medications to supplement existing therapies. 5,6 For instance, incretin-based therapies such as glucagon-like peptide-1 (GLP-1) receptor agonists (e.g., exenatide and liraglutide) and dipeptidyl peptidase-4 inhibitors (DPP-4) (e.g., vildagliptin, sitagliptin, saxagliptin, and linagliptin) 7 are remarkably effective in controlling glycemia with low risks of hypoglycemia. Secretion of GLP-1 is glucose dependent and rapidly inactivated by DPP-4. In the presence of glucose, GLP-1 causes pancreatic β-cell activation and release of insulin. Whereas DPP-4 inhibitors are weight neutral, GLP-1 receptor agonists are associated with weight loss. Similarly, sodium glucose co-transporter 2 (SGLT2) inhibitors (e.g., Dapagliflozin) are a class of drugs that improve hyperglycemia by blocking reabsorption of glucose along with sodium from the proximal renal tubule. These drugs can provide benefits in improving glucose control independently of insulin action, provide a low risk of hypoglycemia, have use in both type 1 diabetes and T2DM, and provide subsequent blood pressure control as a result of simultaneous sodium excretion. 8
Nevertheless, long-term clinical applications for some conventional T2DM therapies have still been problematic. Examples include the insulin secretogogues, which have been used as hypoglycemic agents available for T2DM patients for many years. This group of drugs does not address the insulin resistance phenomenon and has poor drug tolerance and undesirable side effects. 9 The thiazolidinedione (TZD) group of drugs is the first in a class that shows antidiabetic activity by ameliorating the insulin resistance of peripheral target tissues. 10 The insulin-sensitizing activity of TZDs is mediated through PPARs, which are a subclass of the nuclear receptor superfamily directly regulating lipid and carbohydrate metabolism. 11 Although these synthetic drugs showed significant therapeutic potential, their use has already been restricted due to severe side effects, including weight gains, enhanced cancer incidence, fluid retention, hemodilution, and heart failure. 12 An associated increased risk of fractures in women taking TZDs for T2DM and increased hepatoxicity led to the withdrawal of one drug in the class, troglitazone, from the market. 13,14 As a result, there remains an urgent need for safer and more effective therapies that are suitable for life-term usage.
Other potential insulin sensitization pathways involve adipokine production and signaling, including tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), C-reactive protein (CRP), plasminogen activator inhibitor-1 (PAI-1), angiotensinogen, resistin, leptin, adiponectin, 15 and RPB-4. 16 These adipokines have profound effects on insulin resistance and vascular homeostasis. Of these proteins, adiponectin is one of the best-characterized hormones from adipocytes, the levels of which have been shown to directly correlate with insulin sensitivity. Treatment with TZDs stimulates adiponectin gene expression and increases circulating adiponectin concentrations in obese mice and insulin-resistant obese humans, 17 thus, the effect of TZDs on adiponectin secretion may explain, at least partially, the glucose-lowering effect of these drugs in patients with T2DM.
In addition to prescription drug therapies, it is desirable to have safe nutritional approaches for preventing and/or ameliorating insulin resistance. The present study was designed to test in vivo in high-fat diet (HFD)–induced and db/db mouse models to search for safe and efficacious bioactive materials from natural sources that elevate adiponectin secretion from adipose tissues, thereby improving insulin sensitivity.
Materials and Methods
In vitro activity of aloesin and aloesinol
Two compounds, aloesin and aloesinol, were tested at 30 μM for adiponectin induction activity in differentiated 3T3-L1 cells.
High-fat diet–induced diabetes mouse model
A HFD-induced diabetic animal model was developed and employed. Methodology of the diabetic model induction was first explained by Surwit et al. in 1988.
18
In brief, impaired glucose tolerance and T2DM-like symptoms were induced in male C57BL/6J mice (n=7) when fed a HFD (45% kcal) for 8 weeks. Water was provided ad libitum. Animals were maintained in a temperature-controlled room (22.2°C) on a 12-hr light–dark cycle. Body weight, blood glucose, cholesterol, and triglycerides (TGs) were monitored every week. Once induction was confirmed, administration of test articles and controls was initiated. GW1929, N-(2-benzoylphenyl)-O-[2-(methyl-2-pyridinylamino) ethyl]-
db/db mice
Diabetic male C57BL/KsJ (db/db) mice and their litter mates (db/+), purchased from the Jackson Laboratory (Bar Harbor, ME) at 7 weeks old, were housed at 72°F, and 50% relative humidity with a 12-hr light and dark cycle. Mice were maintained on standard rodent chow and were provided with water ad libitum. GW1929 (5 mg/kg) and UP780 (375 mg/kg) were administered to mice (n=8) by daily gavage for 10 weeks. Blood samples were collected from the tail vein. All procedures performed were in compliance with the Animal Welfare Act and US Department of Agriculture regulations.
Glucose and insulin tolerance tests
Animals were fasted overnight (db/db) or for 3 hr (HFD) and received i.p. administration of glucose at a dose of 2 mg/kg (HFD), 3 g/kg (db/db), or insulin (human recombinant expressed in yeast, Sigma, St. Louis, MO, lot #055K1321) at a dose of 0.5 IU/kg. Blood glucose levels were determined at time 0 (before glucose or insulin injection), 30, 60, 90, and 120 min post glucose or insulin delivery. Blood samples were obtained from the tail vein. 21 –23
Assays
Blood glucose levels were measured using the Prestige IQ® Blood Glucose test strips (Walgreen, Home Diagnostics, Inc., Ft. Lauderdale, FL) and monitoring system. Total cholesterol and triglyceride levels were measured using the CardioChek Analyzer with PTS Panels Test Strips (Polymer Technology System, Inc, Indianapolis, IN). Plasma insulin levels were measured with an enzyme-linked immunosorbent assay (ELISA) kit for insulin (Crystal Chem, Chicago, IL). Blood was collected by tail vein and centrifuged for plasma.
Statistical analysis
Data were analyzed using Sigmaplot (Version 11.0). The results were represented as mean±1 standard deviation (SD). Statistical significance between groups was calculated by means of single-factor analysis of variance (ANOVA) followed by a paired t-test. P values less or equal to 0.05 (P≤0.05) were considered as significant. When normality tests failed, data for nonparametric analysis were subjected to Mann–Whitney sum ranks for t-test and Kruskal–Wallis one-way ANOVA on ranks for ANOVA.
Results
Effect of aloesin and aloesinol on adiponectin levels
Purified aloesin and aloesinol at a concentration of 30 μM were active in enhancing adiponectin production, as shown in Fig. 1. Average increases in adiponectin levels of 3.26-fold and 1.79-fold were observed for aloesin and aloesinol, respectively.

Effect of UP394 (aloesin) and UP396 (aloesinol) on adiponectin levels secreted into the medium of differentiated 3T3-L1 cells. 3T3-L1 cells were induced to differentiate then treated with UP394 (aloesin) and UP396 (aloesinol) at concentrations of 30 μM for 48 hr. Adiponectin concentrations in the culture medium were determined with an enzyme-linked immunosorbent assay (ELISA) kit for adiponectin.
Effect of aloesin and aloesinol in vivo
As seen on Fig. 2, significantly improved glucose use was observed in a glucose tolerance test at times 60, 90, and 120 min after i.p. glucose load for GW1929 and aloesin-treated diet–induced diabetic C57BL/6J mice. Animals treated with aloesinol, however, did not show improvement of glucose clearance versus vehicle treatment at the P≤0.05 level, although a clear trend of improvement was observed. Similarly, for the insulin tolerance test, a significant glucose clearance was observed at time points of 30, 60, and 90 min after i.p. insulin for aloesin, aloesinol, and GW1929 when compared to vehicle (Fig. 3).
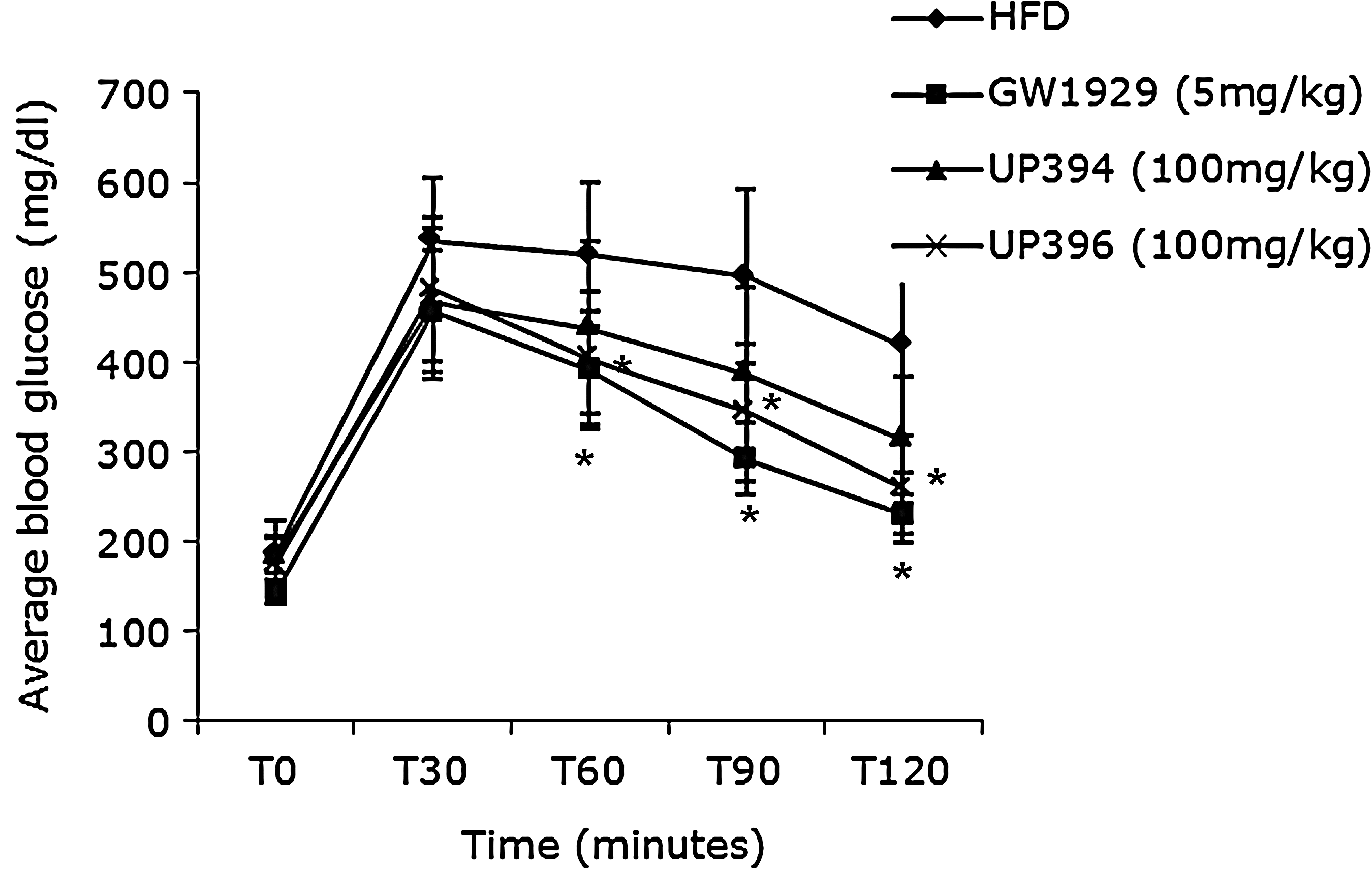
Glucose tolerance test. An intraperitoneal (i.p.) glucose tolerance test was conducted on C57BL/6J mice at a dose of 2 grams/kg on day 18 after treatment. Mice were treated with GW1929 (5 mg/kg), UP394 (100 mg/kg), UP396 (100 mg/kg), and vehicle i.p. Blood glucose levels were measured at time 0, 30, 60, 90, and 120 min. Treatment lasted for 4 weeks. The data are means±standard deviation (SD), n=7, P<0.05 (*). HFD, high-fat diet.

Insulin tolerance test. An intraperitoneal (i.p.) insulin tolerance test was performed on C57BL/6J mice at a dose of 0.5 u/kg on day 24 of active treatment. Animals were fasted for 3 hr before insulin injection. Mice were treated with GW1929, UP394, UP396, and vehicle for 24 days. Blood glucose levels were measured at time 0, 30, 60, 90, and 120 min. Treatment lasted for 4 weeks. The data are means±standard deviation (SD), n=7, P<0.05 (*). HFD, high-fat diet.
The insulin-sensitizing activity of aloesin and aloesinol was further demonstrated by the activity of these compounds to lower plasma insulin levels in the treated animals. The reference compound GW1929, as expected, decreased insulin levels significantly by 50.2% compared to vehicle. Similarly, aloesin and aloesinol also decreased insulin levels significantly by 37.9% and 46.7%, respectively, compared to that in the vehicle-treated mice (Fig. 4).
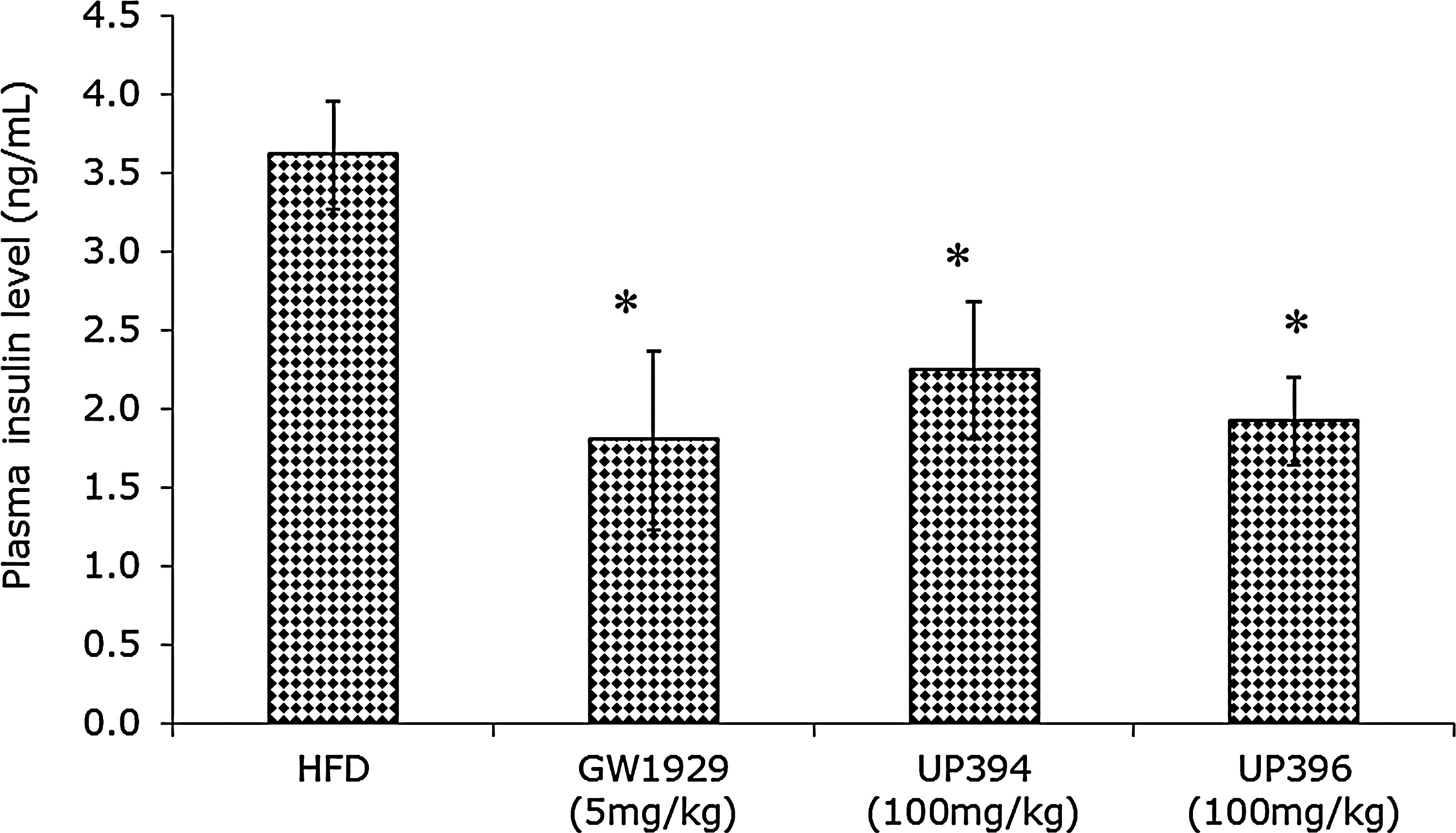
Effect of UP394 and UP396 on plasma insulin levels in high-fat diet (HFD) –induced diabetes model. Animals (n=7) were treated with GW1929 (5 mg/kg), UP394 (100 mg/kg), UP396 (100 mg/kg), and vehicle intraperitoneally (i.p.) for 4 weeks. Blood was collected by tail vein and centrifuged for plasma after 4 weeks of treatment. The plasma insulin levels were measured with an enzyme-linked immunosorbent assay (ELISA) kit for insulin (Crystal Chem, Chicago, IL). P<0.05 (*).
Efficacy of UP780 in db/db mice
Fasting glucose levels of vehicle-treated db/db mice increased over time during the 10-week treatment period, and, as expected, GW1929 treatment maintained glucose at the pretreatment baseline level. Treatment with the aloesin–aloe polysaccharide preparation UP780 (375 mg/kg) also reduced glucose levels substantially starting from the week 5 of treatment. Fasting blood glucose levels were significantly lower in the UP780-treated group at weeks 6, 7, 9, and 10, when compared to vehicle-treated group. After 10 weeks of treatment, the glucose level in the animals treated with UP780 was reduced to 54% of that in the animals treated with vehicle (Fig. 5).

Fasting blood glucose level. Weekly fasting glucose levels of male db/db mice treated with GW1929, UP780, and vehicle for 10 weeks. Animals (n=8) were provided with T2018 rodent diet ad libitum except when fasting. Animals were fasted overnight before measurements were taken. The values indicated are means±standard deviation (SD). Fasting blood glucose levels were significantly lower for GW1929 and UP780 at weeks 6, 7, 9, and 10, when compared to vehicle P<0.05 (*).
The effect of UP780 treatment on insulin resistance was demonstrated with the oral glucose tolerance test (OGTT). Significant glucose use was observed at times 0, 60 (UP780 treatment only), 90, and 120 min for both GW1929-treated and UP780-treated mice when compared to vehicle-treated animals (Table 1).
An oral glucose tolerance test was conducted on db/db mice (n=8) at a dose of 3 grams/kg after 10 weeks of treatment. Animals were fasted overnight before glucose load. Mice were treated with GW1929, UP780, and vehicle for 10 weeks. Lean control db/+ mice (n=8) treated with vehicle were included in this study. Blood glucose levels were measured at time 0, 30, 60, 90, and 120 min.
Data are expressed as mean±standard deviation (SD).
P<0.05.
UP780 reduced TGs significantly compared to the vehicle group after treatment for 10 weeks. A 34% reduction of TGs was observed in the animals treated with UP780, and 43% reduction was observed in the reference compound group as compared to vehicle only treated animals (Fig. 6).

Fasting blood triglyceride (TG) levels. Fasting TG levels of male db/db mice (n=8) treated with GW1929 (5 mg/kg), UP780 (375 mg/kg), and vehicle were measured at baseline and week 9. Animals were fasted overnight before measurements were taken. Data are expressed as mean±standard deviation (SD). A significant reduction in TG levels was found in animals treated with GW1929 and UP780 when compared to vehicle after 9 weeks of treatment, P<0.05 (*).
Efficacy of UP780 in HFD diabetes model
As depicted on Table 2, diet-induced diabetic C57BL/6J mice treated with 200 mg/kg UP780 and 5 mg/kg GW1929 showed statistically significant reductions of fasting blood glucose levels at all measurement times compared to vehicle-treated mice.
High-fat diet (HFD)–fed mice (n=7) were administered GW1929 (5 mg/kg), UP780 (200 mg/kg), and vehicle orally for 10 weeks. Regular rodent diet–fed mice (n=7) treated with vehicle were included in this study. Fasting blood glucose level was measured at baseline, and weeks 4, 6, 8, and 10 using the Prestige IQ Test Strips. Data are expressed as mean±standard deviation (SD).
Values in parentheses indicate percent decrease of HFD.
P≤0.05.
P≤0.001.
When the glucose or insulin tolerance test was conducted after 6 or 10 weeks of daily oral treatment, respectively, a statistically significant improvement in glucose clearance was observed at all measurement times post i.p. glucose or insulin load for animals treated with 200 mg/kg UP780 and 5 mg/kg GW1929 compared to diabetic animals treated with vehicle (Figs. 7 and 8).

Glucose tolerance test. An intraperitoneal (i.p.) glucose tolerance test was conducted on week 6 after onset of treatment. High-fat diet (HFD)–fed mice (n=7) were administered GW1929 (5 mg/kg), UP780 (200 mg/kg), and vehicle orally for 10 weeks. Regular rodent diet–fed mice (n=7) treated with vehicle were included in this study. Mice were injected glucose at a dose of 2 grams/kg after 3 hr of fasting. Blood glucose levels were measured at time 0, 30, 60, 90, and 120 min. Data are expressed as mean±standard deviation (SD). *P≤0.05.
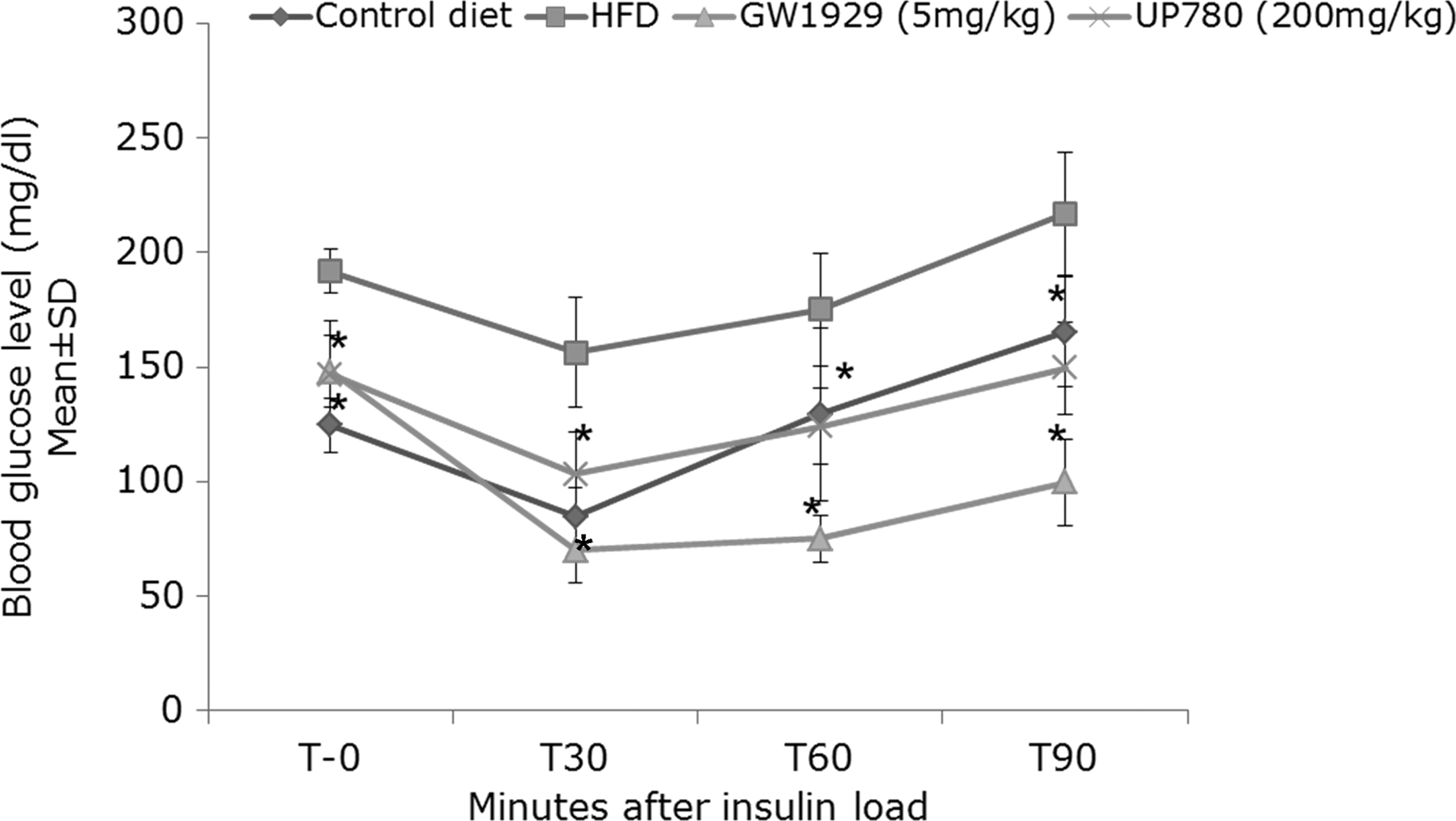
Insulin tolerance test. An intraperitoneal (i.p.) insulin tolerance test was conducted on week 10 after onset of treatment. High-fat diet (HFD)–fed mice (n=7) were administered GW1929 (5 mg/kg), UP780 (200 mg/kg), and vehicle orally for 10 weeks. Regular rodent diet–fed mice (n=7) treated with vehicle were included in this study. Mice were injected insulin at a dose of 0.5 u/kg after 3 hr of fasting. Blood glucose levels were measured at times 0, 30, 60, 90, and 120 min. Data are expressed as mean±standard deviation (SD). *P≤0.05.
Plasma insulin levels were determined at baseline, week 4, and week 10, and as seen in Fig. 9, a progressive increase in plasma insulin level was observed for vehicle-treated mice, whereas GW1929- and UP780-treated mice showed reductions in plasma insulin level of 37.3% and 28.5% at week 4 and 79.9% and 32.2% at week 10 when compared to respective pretreatment baseline insulin levels.
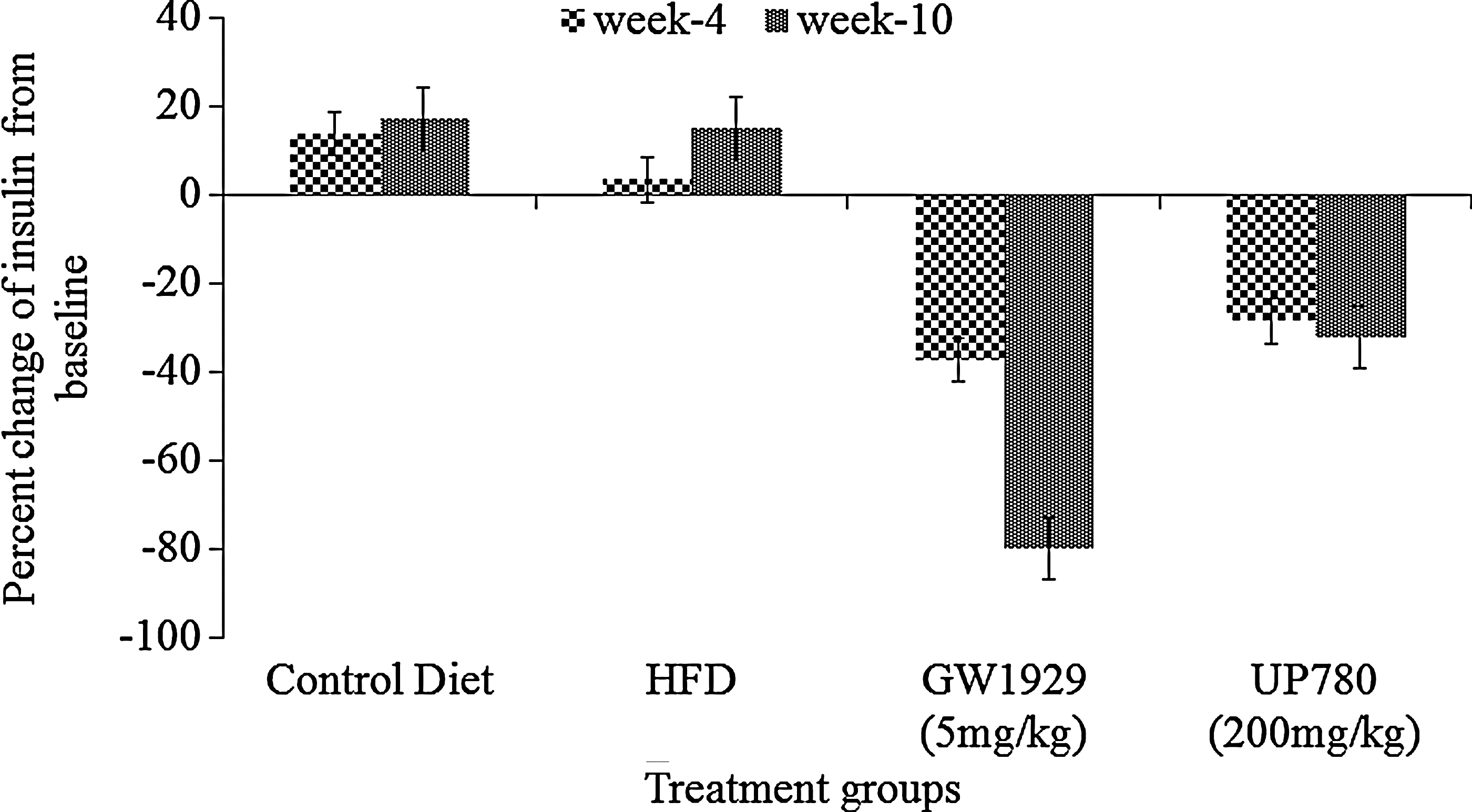
Plasma insulin level compared to baseline. High-fat diet (HFD)–fed mice (n=7) were administered orally with GW1929 (5 mg/kg), UP780 (200 mg/kg), and vehicle for 10 weeks. Control diet–fed mice (n=7) treated with vehicle were included in this study. A blood sample was collected from a tail vein from 3-hr fasted mice at baseline, week 4, and week 10 after treatment. Plasma insulin levels were measured with an enzyme-linked immunosorbent assay (ELISA) kit for insulin. Data are expressed as percent change of insulin from baseline.
Fasting blood triglyceride and total cholesterol were measured at pretreatment baseline, and at week 2 and week 10 after treatment. When compared to vehicle-treated mice, a reduction in blood triglyceride levels of 17.5% and 7.9% was observed at week 2 and 23.9% and 5.4% at week 10 for GW1929 and UP780, respectively (Table 3). These changes did not achieve significance at the P≤0.05 level.
Fasting total cholesterol and triglyceride levels were measured at baseline, week 2, and week 10 using 15- to 20-μL blood samples obtained from the tail veins of mice fasted for 5 hr (n=7) using the CardioChek Analyzer with PTS Panels Test Strips. Data are expressed as mean±standard deviation (SD).
Discussion
Insulin resistance is now recognized as a major determinant in the pathogenesis of metabolic disorders, including T2DM, which co-exists with other disorders such as obesity, dyslipidemia, hypertension, and cardiovascular disease, collectively termed insulin resistance–associated disorders (IRAD). 1 A linkage of insulin resistance to adiponectin is suggested by the findings that patients with T2DM experience low levels of circulating adiponectin, 24 –26 and the observation that plasma adiponectin levels from diabetic mouse models are increased in response to PPARγ agonist treatment, which improves insulin sensitivity. 27
Other studies have reported beneficial effects of aloe and aloe components with metabolic disease animal models, including alloxan-induced diabetes, 28 streptozotocin-induced diabetic rats, 29 –35 and db/db obese-diabetic mice. 36 However, in all of these studies, the principal active components and the mechanism(s) of action were not fully established.
In the current study, biochemical and biological activities of two aloe chromones and a defined composition UP780 were addressed. These compounds possess metabolic disease–modifying activity as assessed in animal models. Although identified through their adiponectin stimulation activity, it is likely that both aloesin and aloesinol possess other activities, as indicated by the pleiotropic effects observed in the animal models reported here and the multiple pathways effected as reported in the companion paper Tseng-Crank et al. (in preparation). Due to the low content of aloesinol in the aloe plant and being economically unfeasible to obtain a standardized extract to deliver biologically relevant dosage, we proceeded with aloesin for formulation and subsequent tests.
The insulin-sensitizing activity of the aloe chromones was demonstrated by improved glucose and insulin tolerance and reduced fasting glucose and plasma insulin levels in diet-induced diabetic mice. These beneficial metabolic effects were also documented for the aloesin–aloe polysaccharide composition UP780 in both diet-induced diabetic mice and db/db genetically obese-diabetic mice. The beneficial effects on glucose metabolism observed with the UP780 aloe composition are comparable to those found with reference compound GW1929, a selective PPARγ agonist. Treatment of diet-induced diabetic mice and db/db obese-diabetic mice with UP780 or with GW1929 also reduced triglyceride levels. Thus, UP780 appears to possess the beneficial effects like TZD insulin sensitizer compounds. We have found, however, that treatment with UP780 does not produce the weight gain (data not shown) documented for TZDs. 37 Members of thiazolidiones (e.g., troglitazone) are effective in improving glycemic control and insulin sensitivity in patients with T2DM despite their potential to cause weight gain. Besides a favorable increase in adiponectin concentrations, it is also believed that with TZD treatment there is a positive shift in fat distribution from visceral to subcutaneous adipose depots that are associated with improvements in hepatic and peripheral tissue sensitivity. However, as Tang et al. have shown, 38 mice treated with troglitazone had markedly increased recruiting and progression of adipose stem cells into new adipocytes than those treated with placebo. This finding could partially explain why patients taking these drugs tend to put on some weight. On the other hand, in our study, mice treated with UP780 did not show an increase in weight gain at the end of study when compared to untreated mice.
Although our screen for bioactive plant compounds was based on in vitro elevation of adiponectin production from adipocytes, we were unable to show an increase in the plasma level of adiponectin for any of the mice treated with aloesin, aloesinol, or UP780 (data not shown). We hypothesize that aloesin, aloesinol, and the UP780 composition act through additional (or other) pathways. Therefore, we used microarray and qPCR to study the gene expression in liver and adipose tissue modulated by UP780 in diet-induced diabetic mice. In liver, the most profound changes induced by UP780 were reduced expression of lipid synthesis genes and increased expression of genes encoding fatty acid–binding proteins, suggesting a lower systemic availability of free fatty acids, known as inducer of insulin resistance in adipose tissue and muscle. In adipose tissue, multiple genes (including the adiponectin gene) involved in the insulin signaling pathway were induced by UP780, which may account for the enhanced insulin sensitivity and glucose usage observed in our animal studies Tseng-Crank et al. (in preparation). These findings perhaps explain why UP780 showed significant glucose-lowering activity. However, the complete mechanism by which UP780 increased insulin sensitivity without associated body weight gain remains unclear and needs further investigation.
In summary, we have characterized and demonstrated the efficacy of the novel and natural agents aloe chromones aloesin and aloesinol for lowering glucose levels and improving insulin sensitivity in two distinct widely used mouse diabetes models. Specifically, aloesin (UP394) and aloesinol (UP396) from whole leaves of Aloe ferox showed in vitro adiponectin elevation activity and in vivo insulin-sensitizing activity. We also documented that enriched aloe chromone composition in the inner leaf gel powder of A. vera, has a profound glucose-lowering effect as well as enhanced insulin sensitivity.
In conclusion, on the basis of data compiled in the present study and the pilot double-blind placebo-controlled human clinical trial that showed a statistically significant reduction in HbA1C, fasting blood glucose, fructosamine, and plasma insulin level, 39 UP780, a standardized aloe chromone-based composition, could possibly be used as a natural alternative for maintaining a healthy blood glucose level.
Footnotes
Acknowledgments
The authors would like to express their gratitude to Mr. Ken Jones from Aloecorp, Dr. Yuan Zhao, and Dr. Padmapriya Abeysinghe, Dr. Wenwen Ma, Ms. Jennifer Homan, Ms. Julia Foo, and Dr. Julie Tseng-Crank from Unigen for their invaluable support for the completion of this project. The authors thank Dr. Ed Cannon and Dr. Doug Bradley for a critical reading of this manuscript.
Author Disclosure Statement
M.Y., B.C., LB and Q.J. are currently Unigen employees, and therefore with financial interest. M.P. and J.Z. are past Unigen employees without financial interest.