
Abstract
Background:
The alarming increase in the prevalence of metabolic syndrome globally demands serious attention. Recently, we reported increased cellular dysfunction of subcutaneous adipose tissue and monocytes contributing to the insulin resistance and increased inflammation in nascent metabolic syndrome. The major circulating leukocyte is the neutrophil (polymorphonuclear neutrophil, PMN). The aim of the study was to evaluate leukocyte levels in nascent metabolic syndrome uncomplicated by diabetes or cardiovascular disease (CVD).
Methods:
In this study, we compared nascent metabolic syndrome (n=63) versus control subjects (n=44). Fasting blood was collected for complete blood count, insulin, glucose, high-sensitivity C-reactive protein (hsCRP), and nitrotyrosine.
Results:
The white blood cell (WBC) and neutrophil counts were significantly increased in metabolic syndrome compared to controls following adjustment for age, gender, and adiposity. There were significant correlations between neutrophil counts with homeostasis model assessment of insulin resistance (HOMA-IR) and hsCRP, (r=0.29, P=0.004 and r=0.35, P=0.0004, respectively). Similarly the correlations of WBC counts were significant with HOMA-IR and hsCRP (r=0.22, P=0.03 and r=0.31, P=0.0014, respectively).
Conclusion:
These findings suggest that significantly elevated WBCs due to increased neutrophil counts in nascent metabolic syndrome could contribute to the increased risk for both diabetes and CVD.
Introduction
Neutrophils (polymorphonuclear neutrophils, PMNs) are the prominent type of circulating leukocytes (white blood cells, WBCs) constituting approximately 50%–70% of all WBCs and is a critical phagocyte that plays a major role in acute infections. PMNs undergo activation when sensing infection or various proinflammatory mediators and migrate to the site of injury, producing reactive oxygen metabolites involving nicotinamide adenine dinucleotide phosphate (NADPH) oxidase pathway 3,4 and release of cytokines and chemokines. 5,6 Activated neutrophils display strong phagocytic and antimicrobial activity generating numerous reactive oxidants and radicals that can oxidatively damage proteins, lipoproteins, DNA, and lipids of the target cells. 7 Also PMNs possesses a large arsenal of biomediators of inflammation and could contribute to the proinflammatory state of metabolic syndrome.
Previously, we have reported increased monocyte activity in nascent metabolic syndrome, including increased biomediators of inflammation such as interleukin (IL)-1β, IL-6, IL-8, soluble tumor necrosis factor receptor 1 (sTNFR1), Toll-like receptor 2 (TLR2), TLR4, as well as increased biomarkers of oxidative stress such as oxidized low-density lipoprotein (LDL) and nitrotyrosine. 8,9 We have also provided evidence of increased monocytic superoxide anion release and NADPH oxidase activity in metabolic syndrome. 8 However the role of neutrophils contributing toward increased oxidative stress and the inflammatory state in the nascent metabolic syndrome state remains unexplored. In this preliminary report, we determined if PMN counts were increased in metabolic syndrome and correlated with biomarkers of inflammation, insulin resistance, and oxidative stress.
Methods
The subjects were enrolled from Sacramento County through fliers and advertisements in newspapers. Metabolic syndrome was defined using the modified criteria of the National Cholesterol Education Program Adult Treatment Panel III (NCEP ATP III), as described previously. 8,9 The subjects classified as having metabolic syndrome had at least three features to sustain the diagnosis, including central obesity, hypertension or on antihypertensive medications, dyslipidemia [low high-density lipoprotein (HDL), high triglycerides (TG)], and plasma glucose between 100 and 125 mg/dL. The enrolled control subjects had two or less features of metabolic syndrome and were not on any blood pressure (BP) medications. Other exclusion criteria for controls were a fasting plasma glucose (FPG) >100 mg/dL) and TG >200 mg/dL.
Other exclusion criteria for both groups were: Diabetes [FPG >125 mg/dL and/or glycosylated hemoglobin (HbA1c) >6.5], clinical atherosclerosis (coronary artery disease, peripheral artery disease, CVD, etc.), smoking, hypo- or hyperthyroidism, malabsorption, anticoagulant therapy, steroid therapy, anti-inflammatory drugs, statins and other hypolipidemic therapy, hypoglycemic agents, angiotensin receptor blockers (ARBs), TG >400 mg/dL (for metabolic syndrome subjects), oral contraceptives, use of antioxidant supplements in the past 6 months, pregnancy, abnormal complete blood count including increased WBCs, alcohol consumption >1 oz/day, consumption of n-3 polyunsaturated fattay acids (PUFA), postmenopausal women on estrogen replacement therapy, active wounds, retinopathy, recent surgery, inflammatory or malignant disease C-reactive protein (CRP) >10 mg/L, and chronic high-intensity exercisers (exercise >100 min/week).
Informed consent approved by the Institutional Review Board at the University of California at Davis was obtained from participants in the study. After a history was obtained and physical examination, fasting blood was obtained. A complete blood count was determined using the Beckman Coulter LH780 autoanalyzer. Plasma lipid and lipoprotein profile, urea nitrogen, creatinine, aspartate aminotransferase, alanine aminotransferase, glucose, and thyroid-stimulating hormone (TSH) were assayed by standard laboratory techniques in the Clinical Pathology Laboratory. Insulin levels were assayed by enzyme-linked immunosorbent assay (ELISA; Linco Biosystems) and homeostasis model assessment of insuling resistance (HOMA-IR) was calculated from glucose and insulin levels as described previously. 8 Nitrotyrosine was assayed as described previously. 8 Briefly, nitrotyrosine levels were measured in plasma in duplicates using the BIOXYTECH® Nitrotyrosine-EIA kit (Oxis Research ™ International, Inc., Foster City, CA), and the coefficient of variation (CV) for this assay was 4.7%. 8
Data are expressed as mean±standard deviation (SD) or, for skewed variables, as median and interquartile range. Log transformations were applied to skewed data prior to parametric analyses. Comparisons between the control and metabolic syndrome groups were made with two-sample t-tests and analysis of covariance (ANCOVA) to adjust for waist circumference or body mass index (BMI). Spearman rank correlation coefficients were computed to assess the association between relevant variables. Data were analyzed using SAS, version 9.2 (SAS Institute, Cary, NC).
Results
Table 1 depicts the salient characteristic features of the enrolled subjects. The groups were matched for age and gender. All five features of metabolic syndrome were significantly different between the two groups. Additionally, there was a significant increase in BMI in metabolic syndrome patients. There were also significantly increased levels of hsCRP and HOMA-IR in metabolic syndrome patients compared to controls (Table 1).
Results are presented as mean±standard deviation or median (25th percentile, 75th percentile).
P value adjusted for sex, age, and BMI.
BMI, body mass index; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; hsCRP, high-sensitivity C-reactive protein; HOMA-IR, homeostasis model assessment of insulin resistance; WBCs, white blood cells.
It is important to emphasize that all volunteers had no evidence of macroinflammation because a hsCRP value <10 mg/L and a WBC count (WCC) in the reference range were strict inclusion criteria. The WCC was significantly increased in metabolic syndrome compared to controls (P=0.0006). This increase persisted following adjustment for age, sex, BMI, and waist circumference (P=0.01). Similarly, there was a significant increase in neutrophil counts in metabolic syndrome compared to controls (P=0.001). The increase in PMNs also persisted following adjustment for age, sex, BMI, and waist circumference (P=0.01). These adjusted levels of increased WCC and PMNs are shown in Fig. 1.
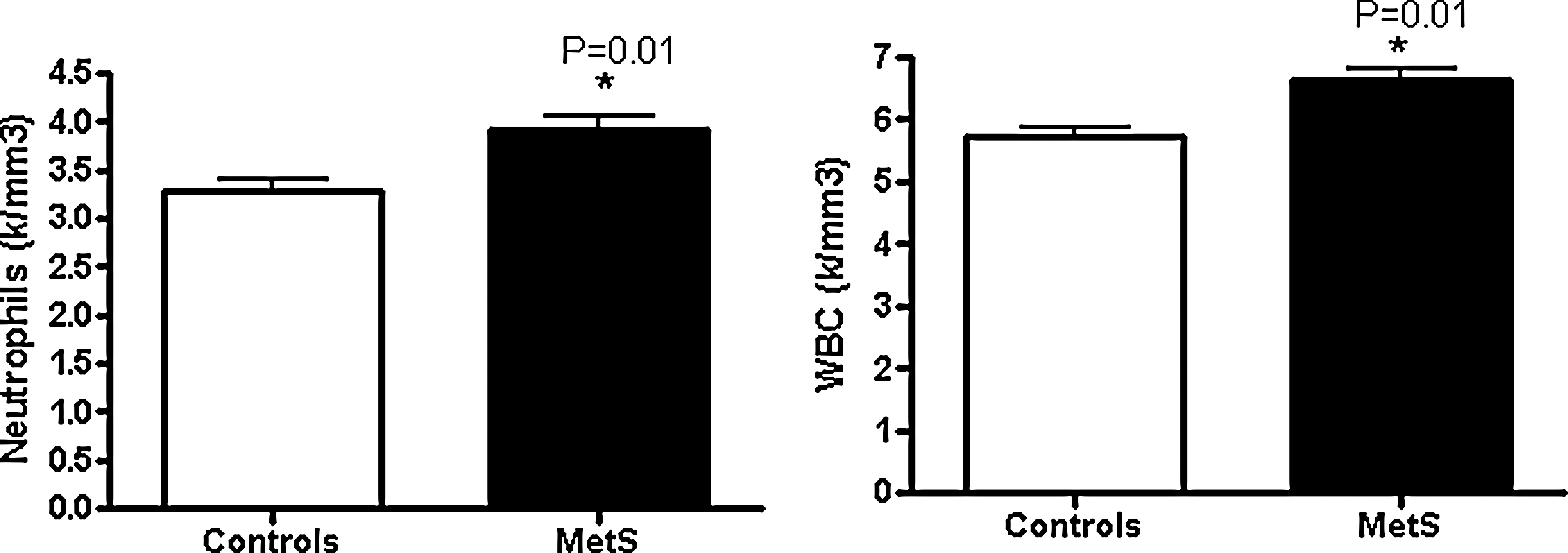
Comparison of white blood cell (WBC) and neutrophil counts in nascent metabolic syndrome (n=63) and controls (n=44). (*) P value adjusted for age, gender, and body mass index (BMI). MetS, metabolic syndrome.
Monocyte counts were also significantly increased in metabolic syndrome patients compared to controls; however, the differences did not reach significance when adjusted for age, gender, and BMI. Lymphocyte counts were not significantly different between the two groups. Both WCC and PMNs were correlated with the accepted biomarkers of insulin resistance (HOMA-IR) and the prototypic biomarker of inflammation, hsCRP, because both inflammation and insulin resistance are typical features of metabolic syndrome. Correlational analyses showed significant positive correlations between neutrophil counts with HOMA-IR and hsCRP (r=0.29, P=0.004 and r=0.35, P=0.0004, respectively). Similarly the correlations of WBC counts were significant with HOMA-IR and hsCRP (r=0.22, P=0.03 and r=0.31, P=0.0014, respectively). Whereas WCC correlated significantly with nitrotyrosine, a biomarker of oxidative stress (r=0.32, P=0.03), there was a trend to a positive correlation between PMNs and nitrotyrosine (r=0.28, P=0.06).
Discussion
The metabolic syndrome state is characterized by increased levels of various inflammatory markers such as increased CRP, tumor necrosis factor (TNF), leptin, plasminogen activator inhibitor-1 (PAI-1), monocyte chemoattractant protein-1 (MCP-1), IL-1, IL-6, retinol binding protein-4 (RBP-4), serum amyloid A (SAA), and decreased IL-10 and adiponectin levels. 2 Several studies have reported increased levels of inflammatory biomarkers in metabolic syndrome. 2,8,9 Previously, we have shown that there is adipose tissue dysregulation and increased monocyte activity in nascent metabolic syndrome without the confounding of diabetes and/or CVD. Both of these cellular dysfunctions contribute to the proinflammatory state and increase oxidative stress and insulin resistance. 8 –10
Because the PMN is the major circulating leukocyte and could contribute to both increased oxidative stress and inflammation, we examined the WBC and its subpopulations in our cohort. We show that both total WBCs and PMNs are increased in metabolic syndrome and that this increase persisted following adjustment for age, sex, BMI, and waist circumference. However, although we have previously reported an increase in monocytic activity, the absolute monocyte count was not increased following adjustment for adiposity. Thus, one can conclude that the increase in total WBCs is largely driven by an increase in PMNs in metabolic syndrome, because both monocyte and lymphocyte counts were not significantly different and the increase in PMNs persisted following all adjustments.
Furthermore, we showed that both WBCs and PMNs correlate significantly with insulin resistance and inflammation as evidenced by HOMA and hsCRP. In addition, we showed that WBCs correlated with nitrotyrosine and there was a positive trend with PMN. Thus, it appears that the increase PMNs could also be a participant in the insulin resistance, increased inflammation, and oxidative stress of metabolic syndrome. However, only studies examining PMN activity in metabolic syndrome and controls can determine this.
Previously, Vozarova et al. 11 measured WBC counts in 352 nondiabetic Pima Indians and reported that higher WBC counts were associated with worsening of insulin action and presaged the development of type 2 diabetes. Other groups have also reported increases in PMN and WBC counts in metabolic syndrome. Kim et al. 12 showed, in a Korean population, increased WBC and PMN counts in male but not female metabolic syndrome patients, unlike our study. However, they did not report on correlations with CRP, insulin resistance, or oxidative stress biomarkers. In accord with our study, Dixon and O'Brien 13 showed increased WBCs and PMNs, but not lymphocyte counts, following adjustment for age, sex, and BMI in metabolic syndrome, but did not undertake correlations like the present study with biomarkers of inflammation, insulin resistance, and oxidative stress. Finally, in a recent report, Buyukkaya et al. 14 showed increases in PMNs and PMN/lymphocyte ratios in metabolic syndrome. However, they did not correct for adiposity, as in our study, and hence cannot conclude that the increase in PMNs is a feature of metabolic syndrome.
In conclusion, we demonstrate that in nascent metabolic syndrome, without the confounding of diabetes or CVD, the increase in WBCs is due to an increase in PMNs. Also this increase in PMNs is correlated with both insulin resistance and inflammation, suggesting that, in addition to adipose tissue dysregulation and increased monocytic activity, neutrophils could also contribute to the pathophysiology of metabolic syndrome by promoting insulin resistance, increased inflammation, and oxidative stress. Hence, future studies are urgently needed that investigate PMN function in metabolic syndrome and matched controls to determine if PMNs have a role in metabolic syndrome and if this role could be modulated to prevent the risk for diabetes and CVD.
Footnotes
Acknowledgments
This study was supported by a grant from American Diabetes Association to I. Jialal.
Author Disclosure Statement
No competing financial interests exist.