
Abstract
Background:
Three-month-old neuron-specific lipoprotein lipase (LPL)-depleted mice (NEXLP−/−) mice are preobese and have normal body weight before developing obesity by 4.5 months. This series of experiments investigated responses to novel environment stimuli and acute sleep deprivation in preobese NEXLPL−/−) mice to test the hypothesis that neuron-specific LPL deletion alters normal adaptive metabolic responses to environmental challenges.
Methods:
Three-month-old, age- and weight-matched, male NEXLPL−/−) (n=10) and wild-type (WT) (n=10) mice were housed in individual metabolic chambers with a 12-hr dark cycle. Food and water intake, locomotor activity, and calorimetry data were recorded in 12-min intervals. Novel environmental responses were elicited by first-time introduction to chambers at dark onset, followed by acclimation, baseline recording, and 6-hr of sleep deprivation on subsequent experimental days.
Results:
NEXLPL−/−) mice displayed a 1.5-fold greater increase in activity in response to a novel environment than seen in WT controls (P=0.0308), and a two-fold greater increase in food intake following acute sleep deprivation (P=0.0117). NEXLPL−/−) mice averaged a 27% higher metabolic rate than WT mice throughout the experiments (P<0.0001). Body weight, composition, and temperature did not differ between murine groups throughout the experiments. Levels of free fatty acid, insulin, glucose, and triglycerides were similar between groups at the terminus.
Conclusions:
A deficiency in neuronal LPL signaling disrupts normal responses to novel environmental exposure and acute sleep deprivation, a maladaptive response that may contribute to weight gain in genetically predisposed mice, and perhaps humans.
Introduction
P
In a sense, both sleep and metabolic homeostasis integrate basic autonomic regulatory mechanisms with higher cognitive processing, consolidating multiple internal and external pressures from nutritional status to environmental queues, manifesting in observable changes in behavior. The NEXLPL−/− mouse model used in this experiment has a primary disturbance in metabolic homeostasis due to neuron-specific deletion of LPL, leading to the development of obesity by 4.5 months of age. 1 LPL is a key enzyme in normal lipoprotein metabolism and plays a large role in many metabolic disorders. 9 For this reason, NEXLPL−/− mice serve as a unique model for investigating the effects of metabolic dysregulation, because the primary dysfunction occurs solely within the central nervous system, and the delayed onset of the obese phenotype provides an opportunity to analyze their behavior before the confounding effects of obesity. 10
Such a central disruption in lipoprotein metabolism has many downstream effects, leading to the proposal that mice with neuron-specific LPL deficiency may have attenuated adaptive responses to common environmental challenges due to a disturbance in pathways responsible for energy balance and arousal, possibly contributing to their development of obesity. On the basis of this hypothesis, this study investigated the effects of neuron-specific LPL deletion on behavioral responses to novel environmental stimuli and acute sleep deprivation in mice to determine the role neuronal LPL plays in the overlapping regulatory systems responsible for arousal and energy balance.
Materials and Methods
Animals
This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Committee on the Ethics of Animal Experiments of the University of Colorado [protocol number B-36110(04)1E]. The generation of NEXLPL−/− mice has been described previously and is accomplished by crossing LPL loxP mice with transgenic mice exhibiting brain-specific expression of cre recombinases. 1 For all sacrifices, mice were anesthetized with intraperitoneal administration of Avertin (2,2,2-tribromoethanol, 0.4 mL/gram), and all efforts were made to minimize suffering. Three-month-old male wild-type (WT) and NEXLPL−/− mice were used, because at this age the NEXLPL−/− mice are not obese, remaining subject to the pathophysiological effects of the gene knockout, but not affected by the physiological consequences of obesity on sleep and activity. 1
A total of 20 mice were used (10 per genotype), requiring five runs of the protocol. Each run used two age- and weight-matched mice per genotype, housed individually, which is the capacity of the environmental chamber. Sample size was selected based on prior experiments with NEXLPL−/− mice using the same calorimetry apparatus, in addition to prior experiments using similar sleep-deprivation and novel environmental stimulus methods, in which similar numbers of mice were sufficient to achieve statistical significance for observed differences. Calculations using statistical power analyses (SigmaStat v.2.01) with assumptions for expected differences between groups based on the literature and preliminary data, α=0.05, and power=0.8, have previously determined that eight animals per group were necessary for this type of experiment, with up to four additional animals per group acceptable to account for the possibility of measurement and recording errors as a result of instrumentation fluctuation or other accidents. 1,11
Apparatus and data collection
The experimental apparatus consisted of a temperature-controlled environmental chamber maintained at 22°C, with a 12-hr light cycle from 08:00 to 20:00 located within a designated laboratory room. Four individual calorimetry chambers within the larger unit were each equipped with food and water dispensers connected to recording equipment and external activity monitors. The chambers were sealed so that air exchange occurred only through the calorimetry recording equipment. To calculate respiratory quotient (RQ) and metabolic rate (MR) and to quantify locomotor activity, an open-ended indirect calorimetry system with a Columbus Instruments Opto M3 multichannel activity monitor was used. This measured food and water intake, oxygen consumption (O2), carbon dioxide (CO2) production, and activity in 12-min intervals for individual mice. Locomotor activity was quantified for each mouse by summing the number of infrared beam breaks caused by movements along the x, y, and z axes.
Experimental protocol
To observe responses to a novel environment, recording began upon first-time introduction to the experimental chambers at the onset of the dark period. Mice were transferred to individual chambers that maintained a 12-hr light cycle from 8:00 to 20:00, a consistent ambient temperature, and ad libitum access to food and water, to which mice were previously accustomed. This provided a new environmental stimulus for the mice, wherein calorimetry measurements, food consumption, and water intake, were recorded to evaluate responses. This method is based on rodent behavior studies and has been validated in subsequent investigations as an effective method of promoting transient wakefulness and increased activity in mice. 12 –14 Mice acclimatized to the chambers over several hours following introduction, and baseline measurements began during the subsequent dark period. After baseline recording, the sleep-deprivation intervention was carried out.
The sleep-deprivation intervention entailed removing the mice for 6 hr prior to dark-onset (14:00–20:00) and maintaining wakefulness by the gentle handling method, a well-established method for maintaining wakefulness in mice, which involved gently touching the mice with a brush or introducing new objects or bedding whenever behavioral signs of sleep were observed. 11,15,16 Mice were not handled beyond this during the sleep-deprivation period, had ad libitum access to food and water, and were not disturbed in any way while eating or drinking. Mice were reintroduced to individual chambers at 20:00, and data collection resumed for the dark period, after which mice were sacrificed for sample collection.
Sample collection
In the first three sets of experimental runs, core body temperature, body weight, and plasma samples were taken from the mice at 20:00 and 8:00 throughout experimental days (n=6 per genotype). Core body temperature was determined via rectal thermometer, and plasma was collected via tail blood sampling. Calorimetry data collection for the first three runs was unsuccessful due to data collection software malfunction, and for this reason two additional experimental runs (n=4 per genotype) were completed. For these final runs, only body weights were recorded at 20:00 during the experimental days to limit the number of interventions during recording days and maximize the amount of calorimetry data available for these groups.
Mice were sacrificed at the end of the experiment and anesthetized as described above. Prior to dissection, body composition was measured by dual-energy X-ray absorptiometry using a mouse densitometer (PIXImus2, Lunar Corp., Madison, WI). Mice were then decapitated for plasma collection, and metabolic parameters were measured as previously described. 9 Specific brain regions and peripheral tissues were also removed for future investigation. Brain tissue was preserved in RNALater, and stored at 6°C; peripheral tissues were flash-frozen in liquid nitrogen, and stored at −80°C. Sample collection time averaged between 15 and 30 min per mouse from anesthetization to complete dissection.
Statistical analysis
Results are reported as means±standard errors of the mean (SEMs), with error bars representing the SEMs. P values <0.05 were considered significant. JMP® 11.0.0 was used to perform all statistical analyses. For analysis of novel environment responses, data recorded for activity, food intake, MR, and RQ were calculated for the average of the first 2 hr of the dark phase on the baseline day and on the new environment day, as well as being analyzed in 1-hr block averages over 24 hr for each day. Two-way mixed analysis of variance (ANOVA) was performed (independent measure, genotype; repeated measure, experimental day). For analysis of sleep-deprivation responses, data recorded for activity, food intake, MR, and RQ were calculated for the average of the first 4 hr of the dark phase on the baseline day and on the post sleep-deprivation day, as well as being analyzed in 1-hr block averages over 24 hr for each day. Two-way mixed ANOVA was performed (independent measure, genotype; repeated measure, experimental day). Figures include the sample size used for each group in analysis. Data for any mouse that did not include complete baseline and experimental day values due to recording instrument error were excluded for analysis. For any ANOVA displaying significant effects, post hoc paired t-tests (for repeated measures) or Student t-tests (for independent measures) were performed.
Results
Novel environment response
By comparing responses to acute novel environment exposure between WT and NEXLPL−/− mice, we were able to investigate whether or not arousal-promoting mechanisms in NEXLPL−/− mice were affected by the gene deletion. Mice characteristically display increased arousal for hours following exposure to a novel environment. 14,17 –19 In this experiment, responses of both genotypes reflected characteristic increases in activity and decreases in food intake, with corresponding decreases in RQ reflective of these changes. However, these changes were dissimilar between genotypes, with NEXLPL−/− mice displaying a 1.5-fold greater increase from baseline activity over the first 2 hr in comparison to WT mice (P=0.0308), and an attenuated decease in food consumption during this time period that failed to reach significance (WT decreased by 70%, P=0.006; NEXLPL−/− decreased by 52%, P=0.055) (Fig. 1).

Response to a novel environment in WT (left) and NEXLPL−/− (right) mice. Activity is displayed in the first row as 1-hr averages beginning at dark onset for experimental and baseline days. (Dashed lines) novel environment day; (solid lines) baseline recording day. In the following rows, bar graphs represent averages of the first 2 hr following dark onset for food intake, metabolic rate (MR), and respiratory quotient (RG). (Solid bars) baseline recording day; (shaded bars) novel environment recording day. Error bars, standard error. Asterisks denote within-group differences, brackets with asterisks represent between-group differences, and the diamond represents trend. See statistical analysis in the Materials and Methods section for details.
Additionally, genotypes displayed characteristic energy expenditure responses following exposure to a novel environment, having similar decreases in RQ (WT decreased 6.2%, P=0.0022; NEXLPL−/− decreased 7.3%, P=0.0003), and increases in MR (WT increased 1.9%, P=0.002; NEXLPL−/− increased 7.1%, P=0.0001) during the first 2 hr of exposure, correlating with exploratory behavior during this time and reflective of changes in physical activity and food intake. Although responses were similar between genotypes, NEXLPL−/− mice maintained a metabolic rate averaging 27% higher than WT mice throughout the interventions (P<0.0001).
Acute sleep-deprivation response
Both genotypes displayed characteristic decreases from baseline activity during the dark period following 6 hr of sleep deprivation, with WT mice decreasing 46.6% (P=0.043) and NEXLPL−/− decreasing 44.8% (P=0.029) (Fig. 2). However, NEXLPL−/− mice displayed a 33% increase in food intake from baseline during this period (P=0.007), a two-fold greater increase compared to control mice (P=0.0117). In addition to the increase in food consumption, NEXLPL−/− mice displayed a two-fold greater decrease in MR compared to WT following the acute sleep-deprivation period (P=0.0408) (Fig. 2).
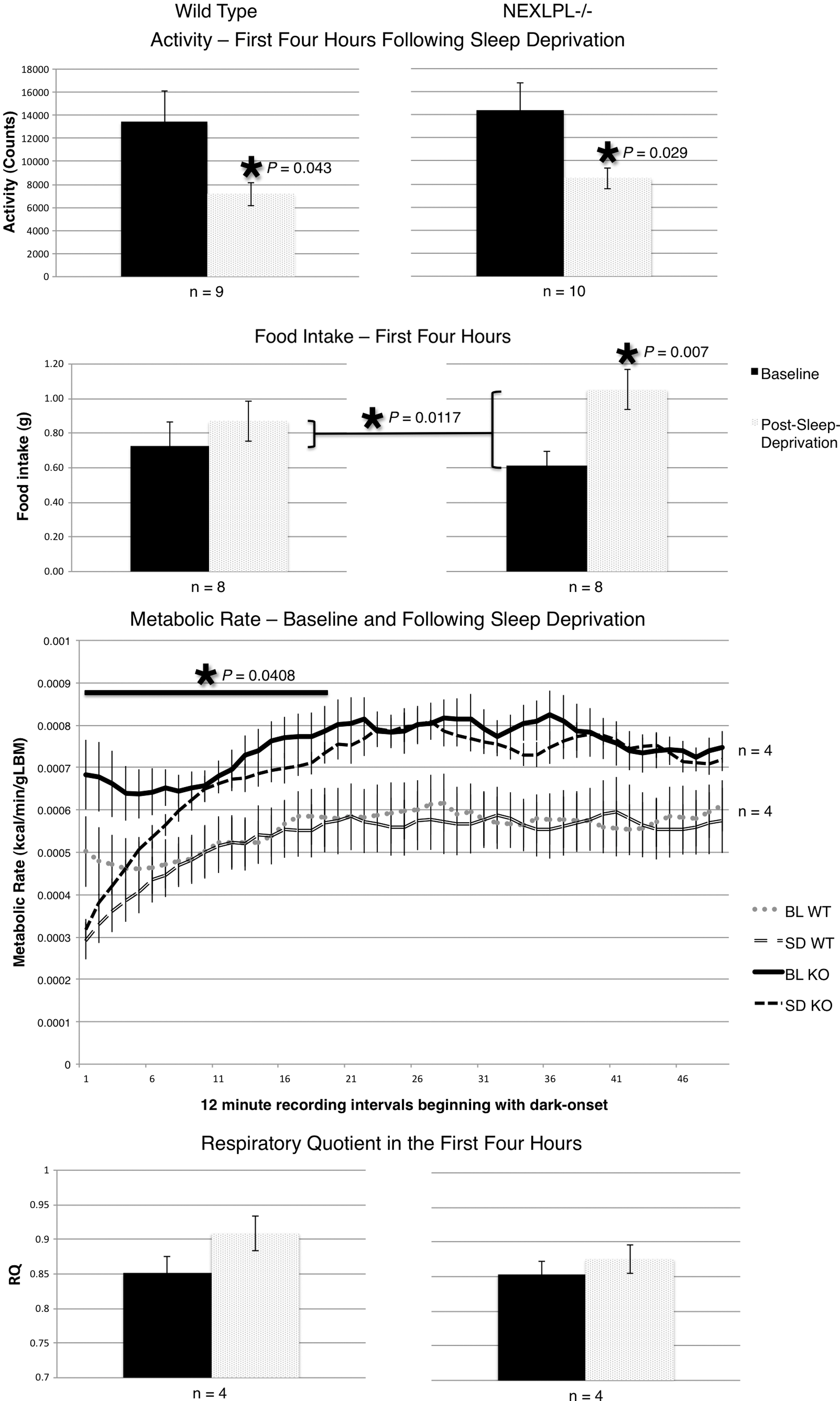
Response to acute sleep deprivation in WT (left) and NEXLPL−/− (right) mice. Bar graphs represent averages of the first 4 hr of dark onset following 6 hr of sleep deprivation for activity, food intake, and respiratory quotient (RQ). See Fig. 1 legend for details. This figure displays metabolic rate (MR) as continuous data collection with time points representing 12-min intervals. (Gray lines) wild-type (WT) mice (baseline: dotted, sleep deprivation response: split); (black lines) NEXLPL−/− mice (baseline: solid, sleep deprivation response: dashed); (solid bar and asterisk) a significant difference between group responses over the first 4 hr of dark onset. BL WT, baseline for wild type; SD WT, standard deviation for wild type; BL KO, baseline for knockout; SD KO, standard deviation for knockout.
Combined experimental genotype characteristics
Although NEXLPL−/− mice displayed baseline nonfasting dark-onset free fatty acid (FFA) levels that were higher than WT mice (P=0.0418), genotype comparisons of nonfasting plasma levels of FFA, insulin, glucose, and triglylcerides (TGs) following the post sleep-deprivation dark period (end-plasma) did not show significant differences between groups. Body weights were similar between groups of mice throughout the experiments, as well as body compositions following the experiments. Both genotypes displayed similar increases in body temperature in response to a novel environment, as well as during the period following 6 hr of sleep deprivation compared to baseline dark-onset body temperatures (Table 1).
WT, wild type; NEXLPL, neuron-specific lipoprotein lipase (LPL)-depleted mice; SE, standard error; FFA, free fatty acids; TGs, triglycerides.
Discussion
Mice with disrupted brain lipoprotein metabolism (NEXLPL−/−) display maladaptive responses to environmental challenges, indicating that intact brain lipoprotein metabolism is necessary for these processes, potentially playing a role in lipid-sensing pathways that mediate the integration of arousal and energy balance. This finding supports the strong connection found between various pathways mediating energy balance and vigilance states, such as the role clock genes play in regulating many physiological and behavioral patterns. 20 Overlapping pathways in the homeostatic regulation of sleep and metabolism are observable by creating disruptions in one system that lead to changes in another, such as in the metabolic consequences of circadian disruptions. 3 However, layers of communication remain undiscovered that are important for characterizing the interaction between peripheral and central regulatory mechanisms responsible for successful adaptive responses in a changing environment.
In our experiment, NEXLPL−/− mice displayed maladaptive behavior in response to environmental stimuli. The exaggerated response to a novel environment could be due to higher baseline levels of anxiety leading to elevated stress and increased activity in response to environmental change. Such hyperactivity in response to a novel environment has been observed in a separate mouse model with a circadian clock transcription factor gene mutation, in which the mice displayed impaired habituation related to impaired memory formation. 21 In this light, the hyperactivity of NEXLPL−/− mice may reflect a similar anxious phenotype resulting from disrupted memory formation, a finding that would correlate anatomically with the pronounced deficiency of LPL in the hippocampus of NEXLPL−/− mice. 10 This may also explain the increased MR NEXLPL−/− mice displayed throughout the experiment while maintaining similar body weights to controls, corresponding to an acutely exaggerated response to the experimental protocol, in contrast to the long-term positive energy balance these mice display. 1 Compared to normal response patterns to environmental novelty characterized broadly in the literature, 22 the hyperactive behavior seen in NEXLPL−/− mice is maladaptive and potentially related to impaired memory formation, a finding being supported by additional data currently being analyzed.
It is possible that pathways responsible for integrating arousal state and energy balance are dysfunctional in NEXLPL−/− mice, i.e., that brain lipoprotein metabolism is a means of lipid sensing and may mediate the interaction between arousal and metabolic pathways. In this sense, the apparent disconnect between arousal and energy balance in NEXLPL−/− mice may be a reflection of a more general impairment in homeostatic signal integration due to deficiency of a key enzyme (LPL) employed for this purpose.
In addition to displaying maladaptive behavior when confronted with a novel environment, NEXLPL−/− mice displayed maladaptive responses to acute sleep deprivation. In this experiment, mice were subjected to a single 6-hr period of sleep deprivation at the end of the light period. Studies using longer periods of sleep deprivation in mice have shown hypothalamic–pituitary–axis activation and loss of body weight despite increased food intake, and it was anticipated in this experiment that WT mice would display a short-lived adaptive response to a single 6-hr sleep-deprivation challenge. 23 Indeed, WT mice displayed a trend toward increased food intake, although this change did not reach significance (Fig. 2). WT mice also displayed characteristically decreased activity from baseline during the beginning of the dark period following sleep deprivation. Although NEXLPL−/− mice displayed similarly decreased activity, they additionally increased food consumption, while having greater attenuation of MR than WT controls. This reveals that mice deficient in neuronal LPL display maladaptive responses to acute sleep deprivation, reflecting a positive energy state and potentially contributing to the development of an obese phenotype later in life. 1
The absence of neuronal LPL may increase sensitivity to both sleep disruption and metabolic disturbance and may disrupt communication between peripheral and central regulatory mechanisms. Prior experiments have shown that sleep restriction leads to metabolic disruptions, and alterations in feeding time have an additive effect through disrupting peripheral clocks. 24 Circadian clock studies have also revealed multiple connections between the regulation of arousal states and energy balance; however, there remain gaps in understanding the distinct connections between the central and peripheral mechanisms regulating these pathways. 4 The findings presented in this study point to an important area of investigation regarding the communication between these central and peripheral mechanisms.
Moreover, understanding the ways in which peripheral and central regulatory pathways communicate is important for the study of human disease. Multiple physiologic systems contain intrinsic circadian activity, and there is evidence to support the link between the uncoupling of these rhythms and disease. 5 For instance, evidence in humans shows that short-term sleep restriction leads to measurable changes in glucose metabolism and adrenal activity that can lead to disease when sustained. 25 Effective adaptation to environmental change involves multiple regulatory pathways, and both intrinsic and extrinsic disruptions in these pathways can play a role in disease development. Extrinsically, sleep disruption correlates with the development of disease states, including obesity, insulin resistance, and hypertension in humans, just as alterations in arousal states such as hypersomnia and insomnia can be a product of intrinsic disease states. 26 –31 Additional studies support the overlap between circadian regulatory pathways, metabolism, and immune function, with disruptions along these pathways leading to a variety of disease states. 32 –36 Similar disruptions are correlated with the development of metabolic syndrome and metabolic disorders, and investigating these pathways has increasing potential for developing new therapeutics. 37,38 For this reason, understanding the ways in which peripheral and central regulatory systems communicate is invaluable for further understanding human disease.
Conclusion
In conclusion, mice with neuron-specific deletion of LPL display maladaptive responses to novel environment exposure and acute sleep deprivation, likely resulting from disruption in multiple neural pathways due to the absence of LPL activity. Although the precise mechanism remains unknown, this suggests maladaptive responses to novel environments and sleep disturbance could contribute to weight gain in genetically predisposed mice, and perhaps humans.
Footnotes
Acknowledgments
Funding for this study was provided through University of Colorado School of Medicine Research Track (grant no. R25 MH080859), the American Diabetes Association Basic Science Research Award (#1-10-BS-45) awarded to Robert H. Eckel (R.H.E.), National Institutes of Health (NIH) grants DK089309 (R.H.E.) and NIH-K12 HD057022 (Principal Investigator, Judith Regensteiner), and a mentored Building Interdisciplinary Research Careers in Women's Health (BIRCWH) training grant awarded to Hong Wang.
Author Disclosure Statement
No competing financial interests exist.