
Abstract
Background:
Obesity and metabolic syndrome are related to systemic functional microvascular alterations, including a significant reduction in microvessel density. The aim of this study was to investigate the effects of exercise training on functional capillary density in the skeletal muscle and skin of obese rats with metabolic syndrome.
Methods:
We used male Wistar–Kyoto rats that had been fed a standard commercial diet (CON) or high-fat diet (HFD) for 32 weeks. Animals receiving the HFD were randomly divided into sedentary (HFD+SED) and training groups (HFD+TR) at the 20th week. After 12 weeks of aerobic treadmill training, the maximal oxygen uptake (VO2max); hemodynamic, biochemical, and anthropometric parameters; and functional capillary density were assessed. In addition, a maximal exercise test was performed.
Results:
Exercise training increased the VO2max (69±3 mL/kg per min) and exercise tolerance (30±1 min) compared with the HFD+SED (41±6 mL/kg per min, P<0.05 and 16±1 min, P<0.001) and with the CON (52±7 mL/kg per min and 18±1 min, P<0.05) groups. The HFD+TR group also showed reduced retroperitoneal fat (0.03±0.00 vs. 0.05±0.00 gram/gram, P<0.001), epididymal fat (0.01±0.00 vs. 0.02±0.00 gram/gram, P<0.001), and systolic blood pressure (127±2 vs. 150±2 mmHg, P<0.001). The HFD+TR group also demonstrated improved glucose tolerance, as evaluated by an intraperitoneal glucose tolerance test, fasting plasma glucose levels (5.0±0.1 vs. 6.4±0.2 mmol/L, P<0.001) and fasting plasma insulin levels (26.5±2.3 vs. 38.9±3.7 μIU/mL, P<0.05). Glucose tolerance did not differ between HFD+TR and CON groups. Exercise training also increased the number of spontaneously perfused capillaries in the skeletal muscle (252±9 vs. 207±9 capillaries/mm2) of the training group compared with that in the sedentary animals (260±15 capillaries/mm2).
Conclusions:
These results demonstrate that exercise training reverses capillary rarefaction in our experimental model of metabolic syndrome and obesity.
Introduction
O
Metabolic syndrome is a cluster of several cardiovascular disease and T2DM risk factors and is associated with insulin resistance, glucose intolerance, central body fat distribution, dyslipidemia, and hypertension. 4 The metabolic alterations that characterize metabolic syndrome are multifactorial and complex and include both environmental and genetic influences. 4 Rats fed a high-fat diet (HFD) for a long period were recently demonstrated to develop structural and functional microvascular abnormalities characterized by microvascular rarefaction and impaired capillary recruitment. 5 Much evidence indicates that changes in the functional capacity of the endothelium and reduced capillary number contribute to elevated peripheral vascular resistance and directly participate in the hemodynamic changes that lead to arterial hypertension. 6 –10
Physical activity plays a key role in the prevention and treatment of obesity and related disorders. 11 –13 Previous studies have indicated that reducing visceral fat and ectopic fat depots is associated with increased insulin sensitivity 14 and reduced hyperglycemia 15,16 as well as with a reduction in the low-grade systemic chronic inflammation that is observed in obesity. 17,18 Regular physical exercise alters endothelial function and improves the vasodilator mechanisms that are essentially mediated by nitric oxide (NO), 19 thereby resulting in reduced blood pressure. 20,21 Improved NO bioavailability has also been associated with increased microvascular density in an animal model of genetic obesity (obese Zucker rats) and a reduction in inflammatory markers that are associated with microvascular rarefaction. 11 In a study with spontaneously hypertensive rats (SHR), exercise training significantly increased capillary density and the capillary-to-fiber ratio in the skeletal muscle and myocardium, completely reversing microvascular rarefaction and hypertension in these animals. 22 In normotensive rats, both interval and continuous exercise training induced similar functional and structural alterations in the skeletal muscle and myocardium microcirculation. 23
Although the role of exercise in the prevention and treatment of obesity-related metabolic disorders is well documented, 24 –27 the effects of exercise on the functional aspects of the microcirculation in experimental models of diet-induced obesity remain unknown. Thus, the main objective of this study was to evaluate the effects of moderate exercise training on the skeletal muscle and skin microcirculation in HFD-fed rats. We also evaluated the effect of exercise on obesity-induced metabolic dysfunction.
Methods
Animals
The handling procedures and testing used in this study have been previously approved by the ethics committee on animal use (CEUA) of the Oswaldo Cruz Foundation–Fiocruz, number LW-21/10 under the provisions of the Brazilian Law #11794/08 for the use of laboratory animals in scientific experiments. The experiments were performed on Wistar–Kyoto rats (WKY; Oswaldo Cruz Foundation Animal Facilities, Brazil) that were kept under controlled humidity (60±10%), temperature (21±2°C), and light (12-hr cycle light/dark) conditions with free access to food and water.
Diet
Previously, we had demonstrated that after 20 weeks of the HFD the rats presented with the main metabolic and hemodynamic characteristics of metabolic syndrome. 5 The commercial diet (CON) (standard chow) contains 23% proteins, 71% carbohydrates, 6% lipids, and 1.3% NaCl, and the HFD contains 14% protein, 56% carbohydrate, 30% lipids, and salt supplementation (standard chow+corn starch+condensed milk+animal fat+0.5% NaCl). The main fat source of HFD was saturated fat (lard). In the present protocol, the animals received the HFD until the end of the training period, completing 32 weeks of this diet.
Training protocol and assessment of maximal oxygen uptake
The training protocol was initiated when the animals had completed 20 weeks of HFD ingestion. The animals were exercised 5 days a week, 1 hr per day, for 12 weeks 28 on a treadmill (Hectron Fitness Equipment, Model HT 2.0, Rio de Janeiro, Brazil). The exercise intensity was equivalent to 65%–70% of the maximal oxygen uptake (VO2max). The VO2max was determined in the pre- and posttraining protocols using a system of open-circuit calorimetry by analyzing the expired gases during a maximal exercise test (MET). The MET was performed on a treadmill coupled to a metabolic box (AQCAD, AVS projects, São Paulo, Brazil) using an inlet flow of air maintained at 3.5 L/min. The system was calibrated using a mixture of 21% O2 and 5% CO2, and the fraction of O2 was measured continuously at an acquisition frequency of 20 Hz. To determine the VO2max, the animals initially remained inside the box for 15 min without exercising to stabilize their oxygen consumption. After this period, the MET began at an initial velocity of 10 meters/min followed by increments of 3 meters/min every 3 min. 29 The test was terminated when the animals were exhausted and remained at the end of the mat on the electrical shock grid for 5 sec. The bars of the shock grid deliver low electrical currents of approximately 2 mA that cause minor discomfort but no harm to the animals.
Hemodynamic measurements
Systolic blood pressure (SBP) and heart rate (HR) were measured using a computerized tail cuff plethysmography system (Visitech Blood Pressure Analysis System, model BP 2000, Apex) in conscious animals in the morning (8 am to 12 am). At least 1 week before recording the arterial pressure, the rats were acclimated for 3 consecutive days using the prewarmed tail cuff device. The platform of the system was adjusted following manufacturer recommendations to maintain temperature around 37°C. Each measurement took around 3 min to be completed and each rat was submitted to two procedures of measurement totalizing 6 min of exposure of the animal to the heating platform. The SBP and HR were measured at the beginning and end of the training protocol.
Metabolic measurements
Intraperitoneal glucose tolerance tests (IGTT) were performed in animals that were fasted for 8 hr. Blood glucose levels were measured 0, 30, 60, and 120 min after the administration of a glucose overload (2.0 grams/kg), 30 and the areas under the glucose curves (AUCs) were calculated. An automatic glucose monitor was used to measure venous blood glucose levels (One Touch Ultra 2®, LifeScan Inc. Johnson & Johnson, Milpitas, CA). Plasma insulin levels were measured using a radioimmunoassay (ImmuChem™ Coated Tube 125I RIA Kit, MP Biomedicals, Santa Ana, CA). Insulin resistance was evaluated using the homeostasis model of assessment of insulin resistance (HOMA-IR) and calculated using the following formula: Fasting glucose (mmol/L)×fasting insulin (μIU/mL)/22.5.
Body composition assessment by dual-energy X-ray absorptiometry
The body fat percentage of the animals was determined after 32 weeks of the HFD using the dual-energy X-ray absorptiometry (DEXA) methodology (Lunar IDXA ME, GE Health Care, Little Chalfont, UK) and quantified using Encore 2008 (GE Health Care Version 12.20) software. The equipment calibration followed the manufacturer's recommendations and was performed on each day of the evaluation. For the analysis, the animals were anesthetized via intraperitoneal sodium pentobarbital (35 mg/kg) administration and then positioned under the scanning area of the equipment in ventral decubitus. The scanning was performed following a sagittal line that was demarcated by the device, which passed under the center of certain anatomical points such as the skull, spine and pelvis.
Intravital fluorescence video microscopy
The animals were anesthetized with pentobarbital [75 mg/kg, intraperitoneally (i.p.)] associated with fentanyl (3 μg/kg); the anesthesia was complemented with an intravenous (i.v.) injection of 5 mg/kg pentobarbital immediately before the administration of the neuromuscular-blocking agent. Fentanyl was injected into the peritoneal cavity 5 min before the beginning of the surgical procedures. The rats were intubated with a polyethylene tube via a tracheotomy, immobilized with pancuronium (1 mg/kg i.v.), and artificially ventilated with room air using a small animal ventilator (Ugo Basile, Varese, Italy). The right jugular vein was catheterized to permit injection of the anesthetic agents and fluorescent dye. The rats' central temperature was monitored with a rectal probe, and their body temperature was maintained at 38±0.5°C using a homeothermic blanket system (Harvard Apparatus, Boston, MA).
The ear skin was scraped, and the animals were placed prone on a Plexiglas pad. The gracilis muscle was exposed via an incision in the right thigh and covered with oxygen-impermeable plastic wrap. The animals were then placed under an upright fixed-stage intravital microscope (Olympus BX51/WI, Center Valley, PA) coupled to a CCD digital video camera system (Optronics, Goleta, CA). An Olympus objective with 10× magnification was used in the experiments, resulting in a total magnification of 100× at the video monitor. After the intravenous injection of 0.15 mL of 5% fluorescein isothiocyanate (FITC)-labeled dextran (molecular weight 150,000), microscopic images of the muscle and skin were successively obtained in real time to count the capillaries using Saisam software (Microvision, Evry, France). Functional capillary density, defined as the total number of spontaneously perfused capillaries per square millimeter of surface area (1 mm2), was determined in random microscopic fields over a 4-min period. Intravital fluorescence video microscopy can be used to identify the capillary network in tissues such as skeletal muscle and skin. In the skin, the capillaries are arranged as interconnected networks, and capillary loops are oriented perpendicularly to the skin surface. In skeletal muscles, the capillaries are arranged along the longitudinal axis of the muscle fibers; thus, the capillaries run parallel to each other.
Quantification of retroperitoneal and epididymal fat
The animals were euthanized using an intravenous administration of pentobarbital overdose, according to ethical committee specifications. After the death of the animals, the retroperitoneal fat was dissected and weighed on an analytical balance accurate to 0.01 gram. Retroperitoneal fat was considered as the fat depots attached to the posterior abdominal wall, around the kidneys, and the abdominal portion of the ureter. Abdominal fat located in the lower abdomen and attached to the epididymis was considered as epididymal fat.
Study design
At 6 weeks of age, the animals (n=54) were randomly divided into two groups and were fed either a CON (SCD, Nuvilab–CR1, Nuvital Nutrients Ltd., Colombo, Brazil) (n=15) or HFD (n=39) for 32 weeks. Saturated fat (lard) was the main fat source in the HFD. At the 20th week, the animals were submitted to hemodynamic and VO2max assessments, and the HFD group was subdivided into sedentary (HFD+SED, n=15) and trained (HFD+TR, n=24) groups. At the end of the experimental protocol (32 weeks), all animals were submitted to hemodynamic, VO2max, metabolic and DEXA assessments, intravital fluorescent video microscopy, and quantification of retroperitoneal and epididymal fat. Figure 1 illustrates our experimental protocol.
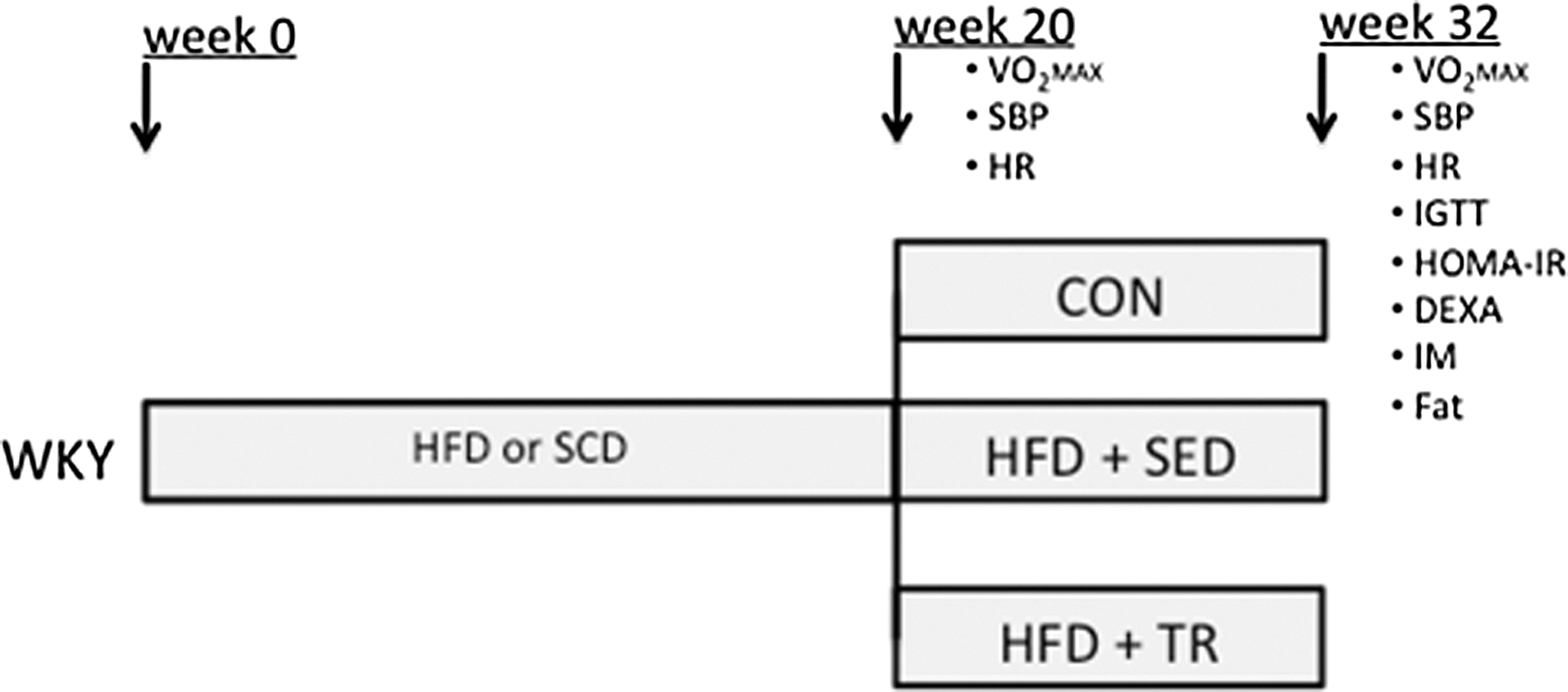
Schematic representation of the experimental protocol. WKY, Wistar–Kyoto rats; HFD, high-fat diet; SCD, standard commercial diet; CON, control group; SED, sedentary; TR, trained; VO2max, maximal oxygen uptake; SBP, systolic blood pressure; HR, heart rate; IGTT, intraperitoneal glucose tolerance test; HOMA-IR, homeostatic model assessment of insulin resistance; DEXA, assessment of body composition by dual-energy X-ray absorptiometry; IM, intravital fluorescent video-microscopy; Fat, quantification of retroperitoneal and epididymal fat.
Drugs
The following drugs were used: sodium pentobarbital, pancuronium bromide and FITC-labeled dextran. All these drugs were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Statistical analysis
The data are expressed as the means±standard error of the mean (SEM). Morphological variables (weight, weight gain, fat weight, and percentage body fat) and microcirculatory parameters (functional capillary density) were compared using analysis of variance (one-way ANOVA). The MET (time to exhaustion), VO2max, and hemodynamic (SBP and HR) variables were compared at the pre- and posttraining times using two-way ANOVA. The post hoc Tukey test was used when P<0.05. All the calculations were performed using a commercially available, computer-based statistical package (GraphPad InStat 5.0, GraphPad Software, La Jolla, CA).
Results
Body weight and body composition
To determine the impact of exercise on the body weight of the HFD-fed animals, the animals were weighed at the end of the 32-week study period. We observed that the HFD significantly increased the body weight of the animals in the HFD+SED group (571±20 grams; P<0.001) compared with that in the CON group (497±10 grams). Physical activity promoted a significant reduction in the body weight of the HFD+TR group animals (479±10 grams; P<0.001), and there were no significant differences between this group and the CON group (Fig. 2A). There were no significant differences in daily food intake between different groups (CON, 15.83±6.33; HFD+SED, 15.09±5.69; HFD+TR, 12.46±4.93 grams/day). DEXA methodology (Fig. 2B) and retroperitoneal (Fig. 2C) and epididymal (Fig. 2D) fat depot weights were used for the body composition assessment. Using DEXA, we observed a significant increase in the body fat percentage of the HFD+SED animals (62±4%; P<0.001) compared with the CON animals (32±3%). Similarly, the HFD+SED group demonstrated a significant increase in the retroperitoneal (0.05±0.00 gram/gram; P<0.001) and epididymal (0.02±0.00 gram/gram; P<0.001) fat depots compared with the CON group (0.02±0.00 and 0.01±0.00 gram/gram, respectively). Physical activity promoted a significant reduction in the retroperitoneal (0.03±0.00 gram/gram; P<0.001) and epididymal (0.01±0.00 gram/gram; P<0.001) fat depots, but the values for the HFD+TR group were still significantly higher than those of the CON group (P<0.05).

Body weight
Evaluation of the time to exhaustion and maximal oxygen uptake
A MET was performed before and after 12 weeks of training. During the MET, the total running time until exhaustion and VO2max (Table 1) was evaluated. No significant baseline differences were observed for the two variables. After 12 weeks of training, the HFD+TR group demonstrated significantly higher VO2max values (70.8±2.4 mL/kg per min; P<0.05) and time to exhaustion (29.8±1.1 min; P<0.001) compared with the CON group (51.8±9.1 mL/kg per min and 17.9±0.6 min, respectively) and with the HFD+SED group (41.5±5.5 mL/kg per min and 16.3±1.0 min, respectively).
The values represent the means±standard error of the mean (SEM).
P<0.05 versus control.
P<0.05 versus HFD+SED.
P<0.05 versus Pre.
HFD+SED, high-fat diet–fed sedentary animals; HFD+TR, high-fat diet–fed trained animals; Pre, before 12 weeks of training in the control group; Post, after 12 weeks of training in the control group; SBP, systolic blood pressure; HR, heart rate; bpm. beats per minute; TTE, time to exhaustion; VO2max, maximal oxygen uptake.
Hemodynamic parameters
SBP and HR were evaluated both before and after 12 weeks of training. We observed that the SBP had significantly higher baseline values in the HFD+SED (152±2 mmHg; P<0.001) and HFD+TR (152±2 mmHg, P<0.001) groups compared with the CON group (133±1 mmHg) (Table 1). The same pattern was observed for HR [HFD+SED 364±5 beats per minute (bpm) and HFD+TR 375±4 bpm; P<0.05] in the comparison with the CON group (341±6 bpm) (Table 1). After 12 weeks of training, SBP and HR were significantly reduced in the HFD+TR group (PAS 127±2 mmHg; P<0.0001, HR 345±3 bpm; P<0.05) compared with the HFD+SED (151±2 mmHg and 367±5 bpm, respectively) and CON (137±2 mmHg and 335±5 bpm, respectively; P<0.05) groups. No significant differences were observed between the HFD+TR and CON groups after the training period.
Evaluation of glucose metabolism and insulin sensitivity
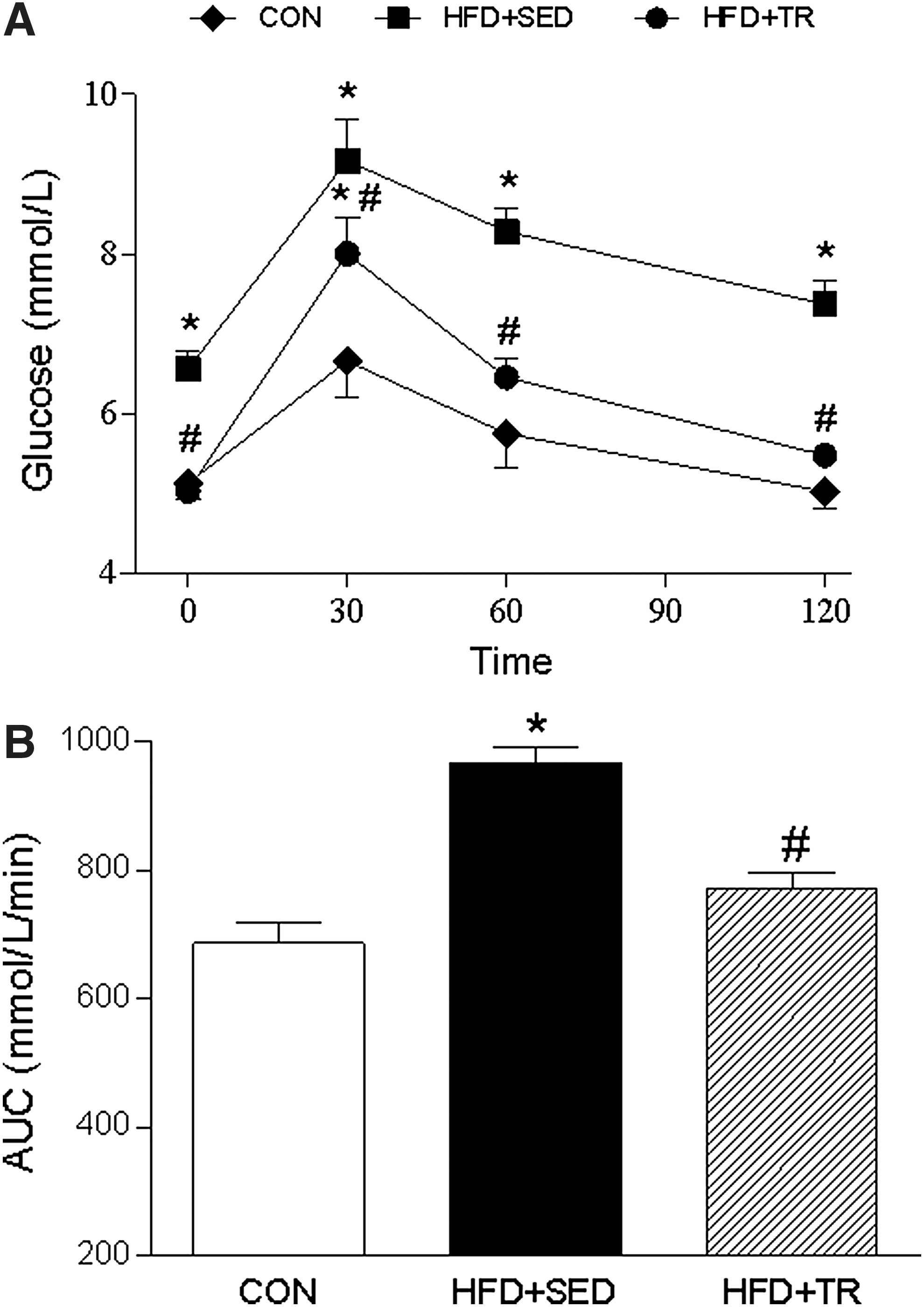
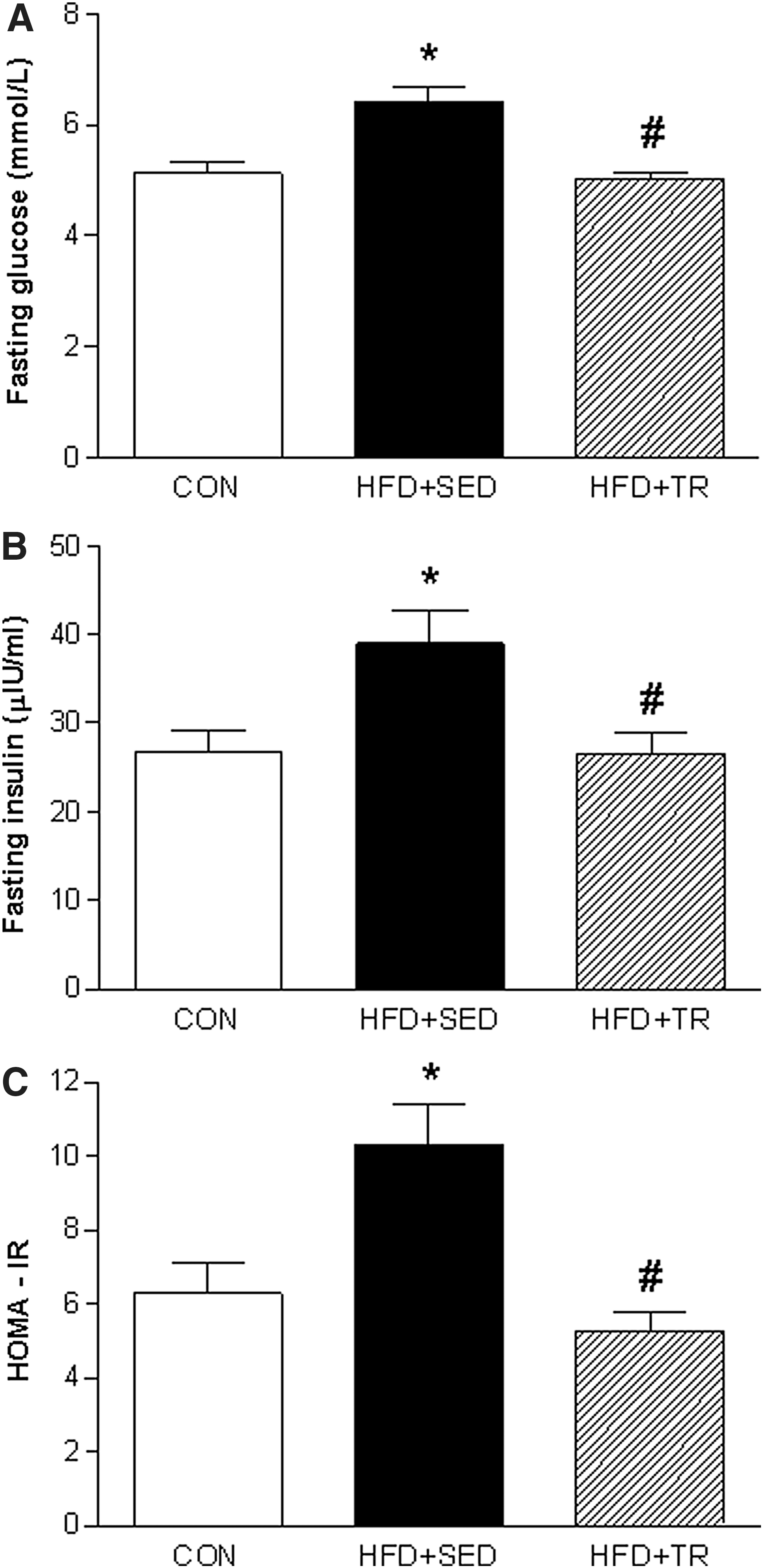
To evaluate the effects of exercise on glucose metabolism, an IGTT was performed at the end of the training period. Figure 3A demonstrates that the HFD significantly increased the blood glucose levels of the sedentary group at all time points of the glycemic curve (P<0.05) compared with the CON and trained (HFD+TR) groups. The AUC values of the HFD+SED group (967±24 mmol/L/min) (Fig. 3B) were significantly higher than those observed in the CON (686±31 mmol/L/min, P<0.001) and HFD+TR (770±25 mmol/L/min, P<0.05) groups. The HFD+SED group also demonstrated significantly higher fasting glucose (6.4±0.2 mmol/L; P<0.0001) and fasting insulin (38.9±3.7 μIU/mL; P<0.05) levels compared with the CON group (5.1±0.2 mmol/L and 26.7±2.5 μIU/mL, respectively). In addition, the animals in HFD+SED group showed insulin resistance, as characterized by HOMA-IR. Exercise significantly reduced the fasting glucose (5.0±0.1 mmol/L; P<0.0001) and fasting insulin (26.5±2.3 μIU/mL; P<0.05) levels and the HOMA-IR values (Fig. 4).
Intraperitoneal glucose tolerance test
Fasting plasma glucose levels
Functional capillary density assessment
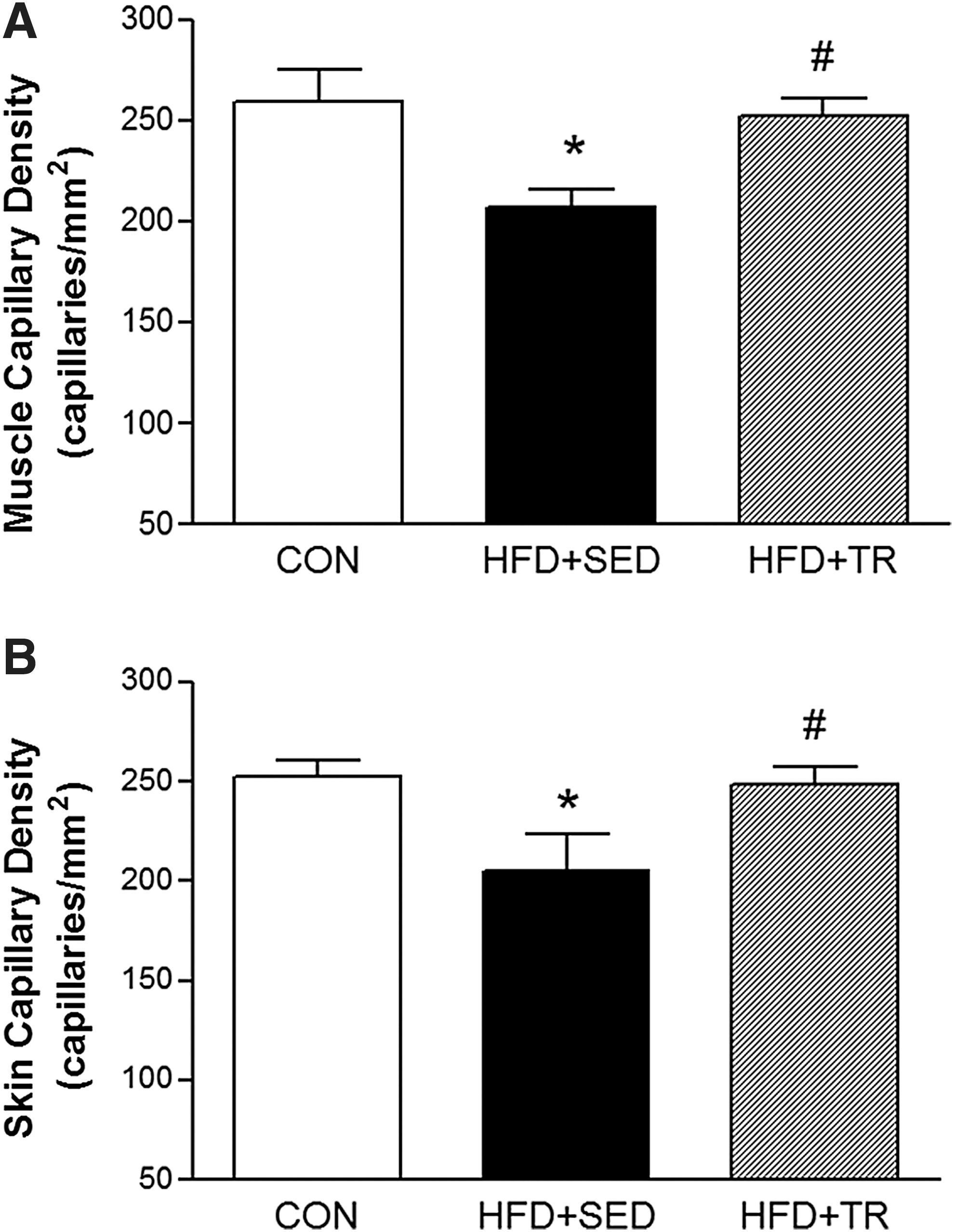
As demonstrated in Fig. 5, A and B, the HFD significantly reduced the number of spontaneously perfused capillaries (functional capillary density) in skeletal muscle (207±9 capillaries/mm2; P<0.05) and skin (205±19 capillaries/mm2; P<0.05) compared with the control group (260±15 and 252±8 capillaries/mm2, respectively). Moreover, exercise training completely reversed the HFD-induced microvascular rarefaction in both locations (skeletal muscle, 252±9 and skin, 248±9 capillaries/mm2; P<0.05). No differences were observed between the CON and HFD+TR groups.
Capillary density of the skeletal muscle
Discussion
The main findings of this study are as follows: (1) The administration of a long-term HFD to rats induces metabolic, hemodynamic, and microvascular alterations representative of metabolic syndrome and obesity; (2) this diet also results in marked central body fat distribution, which is a hallmark of metabolic syndrome; and (3) these alterations are reversed by chronic aerobic exercise training.
The increase in the visceral fat depots can account for the release of various endogenous substances, such as proinflammatory cytokines, many of which have immunomodulatory actions 3 and are associated with inducing a chronic low-grade systemic inflammatory response. 31,32 The inflammatory processes characteristic of obesity and metabolic syndrome are related to visceral adipose tissue and central obesity rather than to total body fat. 33 In summary, there is a close relationship between increased adipose tissue inflammation and altered immune response in obesity, and both factors contribute to the development of related metabolic complications. It is also important to note that the HFD used in the present study undoubtedly induced a significant increase in plasma polyunsaturated fatty acids (PUFAs) concentrations, thus contributing to the inflammatory process that characterizes the experimental model used in the present study.
Obesity and chronic inflammation have also been associated with changes in the microvascular structure and function and have a negative impact on tissue perfusion. 34 –36 Moreover, obesity and metabolic syndrome are characterized by a progressive inability of the cardiovascular system to adequately perfuse tissues and organs during elevated metabolic demand. 37 Such changes are directly related to increased cardiovascular risk 38 because impaired capillary recruitment and endothelium-dependent vasodilation are directly involved in the pathophysiology of hypertension, insulin resistance, and T2DM. 39,40 In a previous study, we had already demonstrated that metabolic syndrome induced by HFD over a long time period (20 weeks) resulted in increased visceral fat, glucose intolerance, and sympathetic nervous system hyperactivity and that these metabolic and neuroendocrine dysfunctions significantly reduced the functional skeletal muscle capillary density, 41 thus participating in the hemodynamic changes that culminate in arterial hypertension. 5,37
Similarly, in the present study, obesity induced by prolonged HFD intake resulted in insulin resistance/glucose intolerance, increased SBP and HR values, and a marked reduction in spontaneously perfused capillaries (functional capillary rarefaction) in the skin and skeletal muscle. Remarkably, aerobic exercise training over the course of 12 weeks reversed all these changes. It is important to note that cutaneous capillary density and reactivity can reflect systemic microcirculation alterations in cardiovascular and metabolic disorders. For example, we previously demonstrated that evaluating skin capillary density could be useful as a marker of the systemic microcirculatory effects of antihypertensive treatments in experimental models of spontaneous arterial hypertension 42 and in hypertensive patients. 43 Skin microcirculation has also been used to evaluate systemic microvascular reactivity in patients with arterial hypertension, dyslipidemia, obesity/metabolic syndrome, 44 and T1DM. 45
Experimental evidence indicates that physical exercise alters blood flow and increases shear stress on the vascular endothelium, thereby stimulating NO release. 46 NO has a potent vasodilator effect by reducing vascular resistance 47 –49 and consequently reducing high blood pressure. 50 Additionally, NO stimulates growth factors, such as vascular endothelial growth factor (VEGF), thus acting as a stimulus for angiogenesis. 51,52 Several studies also demonstrated that exercise training is responsible for the negative modulation of sympathetic activity, thus reducing peripheral vascular resistance. 24,53 The mechanisms involved in blood pressure reduction, such as increased NO synthesis and release, the improvement in autonomic control (enhanced vagal activity), and structural adaptations of the heart, can also modulate HR and promote the consistent reduction in HR observed in the trained animals in our study. 54
In the present study, we also demonstrated that exercise training for 12 weeks reversed insulin resistance and glucose intolerance. However, we did not investigate the molecular mechanisms involved in the beneficial effects of exercise training in glucose metabolism. Several molecular mechanisms have already been shown to be involved in the improvement of glycemic control during exercise training. For example, experimental evidence has demonstrated that muscle contraction increases glucose uptake via adenosine monophosphate (AMP)-activated protein kinase (AMPK) and promotes glucose transporter 4 (GLUT4) translocation to the plasma membrane independent of insulin. 55,56 Exercise also increases GLUT4 expression 15 and reduces oxidative stress, thereby improving insulin sensitivity. 57
The limitations of this study must be considered. First, we did not address the muscle fiber types that were involved in the microvascular alterations observed in our experimental HFD-induced obesity model, which are known to affect glucose and insulin availability differentially. Second, even if functional capillary density is a microcirculatory parameter that is classically considered to reflect tissue perfusion, it was not directly matched to the microvascular blood flow. Third, we used an indirect measurement of insulin sensitivity, which is commonly used in clinical practice. In fact, the oral or intraperitoneal glucose tolerance tests reflect the efficiency of the body to dispose of glucose after a glucose overload. However, it is important to note that glucose tolerance and insulin sensitivity are not equivalent concepts. In addition to metabolic actions of insulin, insulin secretion, incretin effects, and other factors contribute importantly to glucose tolerance. Thus, the glucose tolerance tests provide useful information about glucose tolerance but not insulin sensitivity/resistance per se. Nevertheless, a number of surrogate indexes of insulin sensitivity/resistance incorporate the results of the glucose tolerance tests.
In summary, we have demonstrated that HFD-induced microvascular, hemodynamic and metabolic alterations can be reversed by moderate aerobic exercise training.
Footnotes
Acknowledgments
This investigation was supported by grants from the CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico), FAPERJ (Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro), and Oswaldo Cruz Foundation (Fiocruz), Rio de Janeiro, Brazil.
Author Disclosure Statement
No competing financial interests exist.