
Abstract
Background:
Metabolic syndrome is a complex disorder characterized by the presence of insulin resistance (IR), type 2 diabetes mellitus (T2DM), impaired glucose tolerance (IGT), or impaired fasting glucose (IFG), plus at least two of the following conditions—hypertension, hyperlipidemia, obesity, and microalbuminuria. Metabolic syndrome exposes patients to a greater risk of developing cardiovascular disease (CVD) and is often associated with elevated levels of homocysteine (Hcy). In the current work, we analyzed the expression of nicotinamide N-methyltransferase (NNMT). Because NNMT is involved in Hcy metabolism and participates in the regulation of the cellular and plasma levels of this compound, we explored the role played by the enzyme in metabolic syndrome.
Methods:
Real-time PCR, immunohistochemistry, western blot analysis, and catalytic activity assay were performed to evaluate NNMT expression levels in adipose tissue from 10 Wistar Ottawa Karlsburg W (WOKW) rats, which are an animal model for metabolic syndrome, and from 10 Dark Agouti (DA) rats as the disease-resistant control strain.
Results:
NNMT mRNA, protein, and activity levels were significantly higher in adipose tissue obtained from WOKW rats compared with those observed in adipose tissue of DA rats.
Conclusion:
Data reported in this study represent the first evidence supporting the hypothesis that NNMT could play an important role in the pathogenesis of metabolic syndrome and could have a potential for the development of a targeted therapy.
Introduction
I
The pathogenesis of metabolic syndrome has multiple origins. In particular, obesity and a sedentary lifestyle coupled with unbalanced dietary intake and genetic factors seem to interact and produce the metabolic disorder. Metabolic syndrome represents a cluster of metabolic abnormalities and, due to its multifactorial and complex trait, exposes patients to a greater risk of developing CVD. 3
Several studies reported a correlation between plasma homocysteine (Hcy) levels and CVD. Indeed, hyperhomocysteinemia (HHcy) is associated with an increased risk of ischemic heart disease (IHD) 4 and stroke. 5 Moreover, a recent study showed that elevated Hcy levels are significantly related to abdominal obesity, IGT, hypertriglyceridemia, and elevated blood pressure, which represent the main components of metabolic syndrome. 6
In humans, Hcy is formed by the demethylation of the essential amino acid methionine. First, dietary methionine and adenosine triphosphate (ATP) produce S-adenosylmethionine (SAM) by means of methionine adenosyltransferase. Then, SAM is converted to S-adenosylhomocysteine (SAH) through reactions catalyzed by SAM-dependent methyltransferases. Subsequently, SAH hydrolase catalyzes the breakdown of SAH into adenosine and Hcy. 7
In the present work, we focused on the enzyme nicotinamide N-methyltransferase (NNMT), which catalyzes the N-methylation of nicotinamide, pyridines, and other structural analogs, using SAM as a methyl donor. 8 The reaction catalyzed by NNMT yields N1-methylnicotinamide, which is excreted into the urine, 9 and SAH, which is converted into Hcy by SAH hydrolase. 7 Therefore, NNMT is involved in Hcy metabolism and could contribute to playing an important role in regulating the cellular and plasma levels of this compound. Results reported by Souto et al. demonstrated that one single-nucleotide polymorphism (SNP) within the NNMT gene is associated with high levels of Hcy, suggesting that the enzyme could be a major genetic determinant of plasma Hcy levels. 7
Because HHcy is often associated with metabolic syndrome and NNMT is involved in the metabolism of Hcy, we wanted to explore the involvement of NNMT in metabolic syndrome. In this regard, we evaluated NNMT mRNA, protein, and activity levels by quantitative real-time PCR, immunohistochemistry, western blot analysis, and enzyme assay in adipose tissue from Wistar Ottawa Karlsburg W (WOKW) rats, which represent an animal model for metabolic syndrome, 10 and from Dark Agouti (DA) rats as the disease-resistant control strain.
Materials and Methods
Animals
Eight-week-old male and female DA rats (n=10) and WOKW rats (n=10) were selected for this study. Rats (Prof. Ingrid Klöting, Department of Laboratory Animal Science, University of Greifswald, Karlsburg, Germany) were housed in plastic cages and fed with food pellets and water ad libitum. Animals were maintained at constant temperature (20±1°C) and humidity (50±5%) on a 12-hr light/12-hr dark cycle (7 AM to 7 PM).
Fasting blood was collected from the tails of animals and kept in a heparinized vial. Plasma was obtained by centrifugation at 1000×g for 20 min and then stored at −80°C until assayed. Plasma total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), triglycerides (TGs), and glucose (GL) concentrations were determined using CT F400 CH, HD F080 CH, TR F400 CH, and GL 100F CH kits (Chema Diagnostica, Monsano, Italy), respectively, according to the manufacturer's instructions. The atherogenic index of plasma (AIP) was calculated by log(TG/HDL-C). 11 TC, HDL-C, TG, and GL plasma levels and the AIP of DA and WOKW rats are reported in Table 1.
Values are reported as mean±standard deviation (* P<0.001).
AIP=log(TG/HDL-C)
DA, Dark Agouti; WOKW, Wistar Ottawa Karlsburg W.
Rats were sacrificed, and abdominal fat depots were excised. After removal, adipose tissue samples were partly snap frozen in liquid nitrogen and stored at −80°C, and partly formalin-fixed and paraffin-embedded (FFPE).
All experiments were performed according to the Principles of Laboratory Animal Care. All efforts were made to minimize animal suffering and to reduce the number of animals used.
RNA extraction and cDNA synthesis
An aliquot of frozen tissue (100–150 mg) was homogenized in a lysis buffer, and total RNA was then extracted through the Aurum Total RNA Fatty and Fibrous Tissue Kit (Bio-Rad Laboratories, Hercules, CA). RNA samples were tested by ultraviolet absorption at 260 nm to determine their concentration. The quality of the RNA samples was further confirmed by electrophoresis on denaturated 1% agarose gels. Total RNA (2 μg) was reverse transcribed in a total volume of 25 μL for 60 min at 37°C with Moloney murine leukemia virus (Mo-MLV) Reverse Transcriptase (Promega, Madison, WI,) using random primers.
Real-time quantitative PCR
The cDNA, generated as previously described, was used as template for real-time quantitative PCR to examine NNMT mRNA expression levels quantitatively. The real-time PCR assay was performed using a CFX96 Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA). To avoid false-positive results resulting from amplification of contaminating genomic DNA in the cDNA preparation, all primers were selected to flank an intron. Moreover, PCR efficiency was tested for both primer pairs and found to be close to 1. The primers used were 5′-GAATCAGGCTTCACCTCCAA-3′ (forward) and 5′-TCACACCGTCTAGGCAGAAT-3′ (reverse) for NNMT, and 5′-TCCTTCCTGGGCATGGAGT-3′ and 5′-AGCACTGTGTTGGCGTACAG-3′ for β-actin.
The two genes were run in duplicate for 40 cycles at 94°C for 30 sec and 58°C for 30 sec, using SsoFast EvaGreen Supermix (Bio-Rad Laboratories, Hercules, CA). All samples were tested in triplicate with the reference gene β-actin for data normalization to correct for variations in RNA quality and quantity. Direct detection of PCR products was monitored by measuring the fluorescence produced by EvaGreen dye binding to double-stranded DNA during the amplification process. These measurements were then plotted against cycle numbers. The parameter threshold cycle (Ct) was defined as the cycle number at which the first detectable increase above the threshold in fluorescence was observed. Fold changes in relative gene expression were calculated by 2−Δ(ΔCt) where ΔCt=Ct (NNMT) − Ct (β-actin) and Δ(ΔCt)=mean-ΔCt (adipose tissue from WOKW rats)−mean-ΔCt (adipose tissue from DA rats).
To compare the expression levels of NNMT in adipose tissue from DA rats and from WOKW rats, the ΔCt value was calculated. A small ΔCt represents a high NNMT expression level, whereas a large ΔCt value is attributable to a low expression level.
Immunohistochemistry
Immunohistochemical analyses were performed on 4-μm sections obtained from FFPE adipose tissue blocks and mounted on poly-
Western blot analysis
Aliquots of frozen adipose tissue (250–350 mg), obtained from DA and WOKW rats, were suspended in 500 μL of lysis buffer [phosphate-buffered saline (PBS) containing 1% Nonidet P-40, 0.1% sodium dodecyl sulfate (SDS), 1 mM phenylmethylsulfonyl fluoride (PMSF), and 2 μg/mL aprotinin] and homogenized on ice using an Ultra-Turrax homogenizer (IKA, Staufen, Germany). After centrifugation at 16,000×g for 10 min at 4°C, the supernatant represented the protein extract. Samples containing 50 μg of protein were subjected to 15% SDS polyacrylamide gel electrophoresis (PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes. After regular blocking and washing, the membranes were incubated with rabbit polyclonal antibody (1:1000 dilution; Sigma-Aldrich) against NNMT for 1 hr, followed by incubation with horseradish peroxidase (HRP)-conjugated goat anti-rabbit immunoglobulin G (IgG; Pierce, Rockford, IL) (1:2000 dilution) for 1 hr. NNMT protein was detected using enhanced SuperSignal West Femto Maximum Sensitivity chemiluminescent substrate (Pierce, Rockford, IL). The chemiluminescent signal of NNMT protein detected in blots was acquired using ChemiDoc XRS+ System (Bio-Rad Laboratories, Hercules, CA).
Catalytic activity assay
Aliquots of frozen adipose tissue (250–350 mg), obtained from DA and WOKW rats, were suspended in 500 μL of cold lysis buffer (50 mM Tris-HCl, pH 8.6, 2 μg/mL: aprotinin, 1 mM PMSF, 1 mM dithiothreitol) and homogenized on ice using an Ultra-Turrax homogenizer (IKA, Staufen, Germany). The homogenate was centrifuged at 16,000×g for 20 min at 4°C, and the supernatant was kept at 4°C until assayed. The standard assay mixture contained 50 mM Tris-HCl (pH 8.6), 1 mM dithiothreitol, 5 mM nicotinamide, 1 mM S-adenosyl-
Protein assay
Protein concentration was measured by the Bradford method using bovine serum albumin as the standard. 13
Statistical analysis
Data were analyzed using GraphPad Prism software version 6.00 for Windows (GraphPad Software, San Diego, CA). Differences between groups were determined using the Mann–Whitney U-test, and Student t-test. A P value<0.05 was accepted as statistically significant.
Results
Quantitative real-time PCR
We examined NNMT mRNA expression levels in adipose tissue specimens from 10 DA rats and from 10 WOKW rats by real-time PCR. Differential gene expression measurements (WOKW versus DA) showed a significant (P=0.0026) upregulation of NNMT (two-fold increase) in the adipose tissue of WOKW rats, displaying an almost complete panel of traits underlying metabolic syndrome, compared with that of the DA rats that were chosen as control strain (Fig. 1).
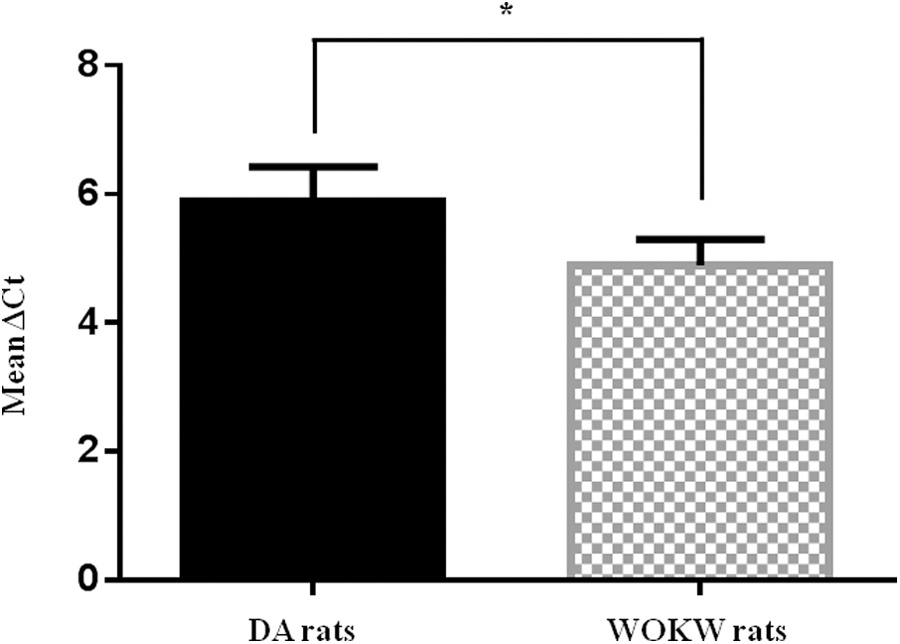
Quantitative real-time PCR analysis of nicotinamide N-methyltransferase (NNMT). In adipose tissue samples from Dark Agouti (DA) rats (n=10) and Wistar Ottawa Karlsburg W (WOKW) rats (n=10), NNMT expression levels were determined as described under Materials and Methods and expressed as ΔCt (mean±standard deviation). Mean ΔCt was calculated as the average of all ΔCt values within each group (*P<0.05).
Immunohistochemistry
Immunohistochemical analyses confirmed results obtained by real-time PCR and revealed an upregulation of NNMT in adipose tissue of WOKW rats. Indeed, adipose tissue of WOKW rats showed a high cytoplasmic positivity for NNMT, whereas adipose tissue of DA rats displayed weak cytoplasmic NNMT immunoreactivity (Fig. 2). In particular, WOKW rats (median, 75%; interquartile range, 62.5%–80%) exhibited significantly (P=0.0286) higher NNMT levels compared with those of DA rats (median, 17.5%; interquartile range, 15%–20%).
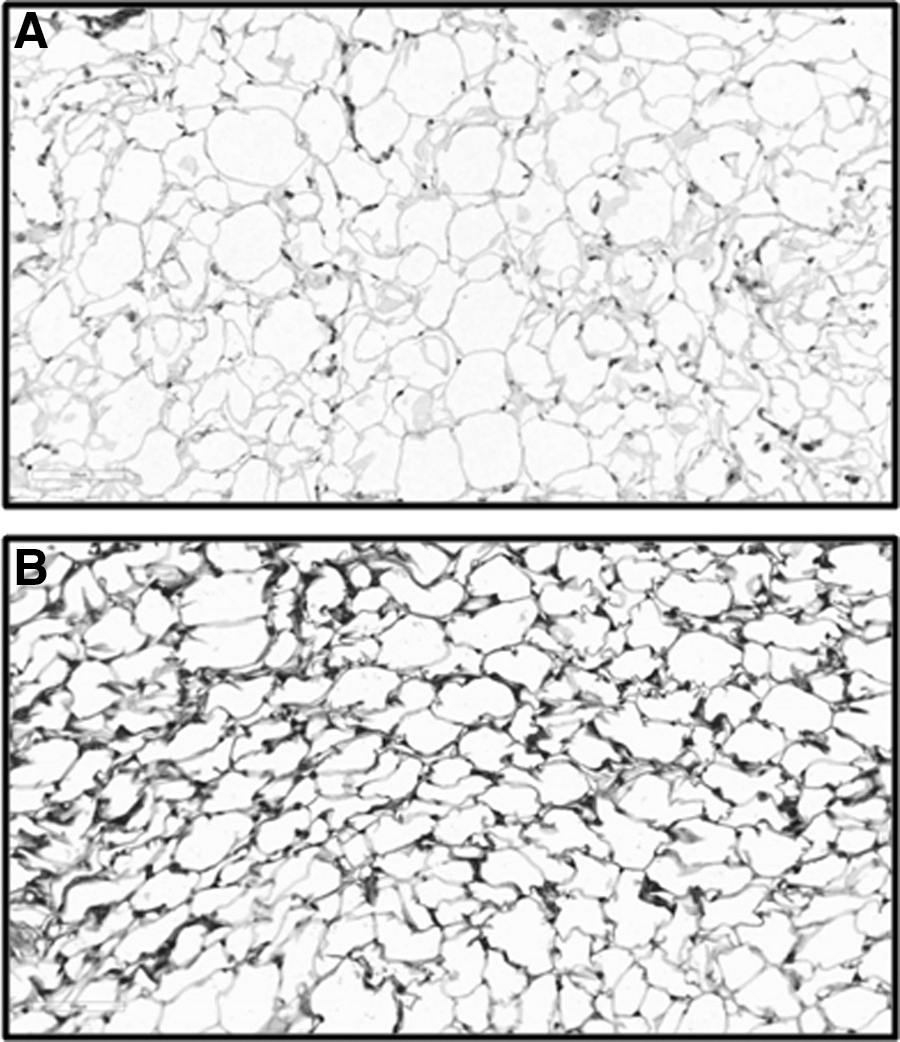
Immunohistochemistry of nicotinamide N-methyltransferase (NNMT). Sections of formalin-fixed and paraffin-embedded (FFPE) adipose tissue samples from Dark Agouti (DA) rats (
Western blot
In adipose tissue samples obtained from DA and WOKW rats, NNMT protein levels were also evaluated by western blot analysis. Results obtained showed a single immunoreactive band at approximately 30 kDa, which corresponds to the known molecular mass of NNMT. Consistent with previous data, lanes loaded with equal protein amounts showed a markedly increased NNMT expression levels in samples obtained from WOKW rats compared with those detected in specimens from DA rats (Fig. 3).

Western blot analysis of nicotinamide N-methyltransferase (NNMT). A 50-μg amount of adipose tissue lysates, obtained from Dark Agouti (DA) and Wistar Ottawa Karlsburg W (WOKW) rats, was subjected to 15% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes. Blots were probed with anti-NNMT antibody and detected with enhanced chemiluminescence.
Enzyme activity
A catalytic activity assay was performed in 10 adipose tissue samples from DA rats and 10 specimens obtained from WOKW rats. The levels of NNMT activity in adipose tissue, expressed in U/mg protein, were statistically significantly higher (P=0.0286) in WOKW (5.630±0.533) than in DA rats (3.030±0.752) (Fig. 4).
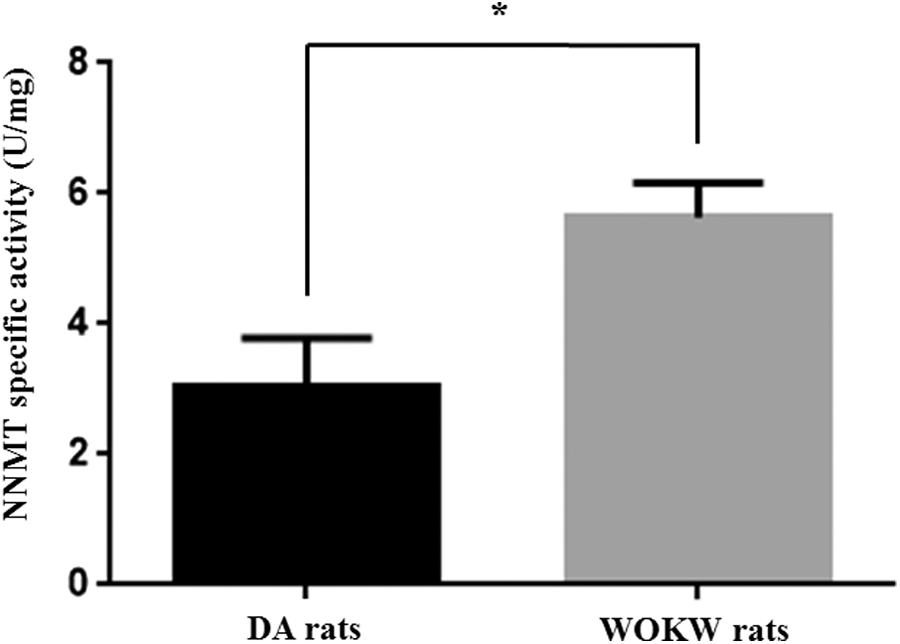
Nicotinamide N-methyltransferase (NNMT) enzyme assay in adipose tissue. NNMT specific activity was analyzed in adipose tissue samples obtained from Dark Agouti (DA) and Wistar Ottawa Karlsburg W (WOKW) rats, by measuring the amount of N1-methylnicotinamide produced. Values are expressed as mean±standard deviation (*P<0.05).
Discussion
Metabolic syndrome is a major clinical challenge, and its escalation has been fuelled by urbanization, surplus energy intake, increasing obesity, and sedentary life habits. Metabolic syndrome is a pathologic condition defined by interconnected biochemical, metabolic, and clinical factors that increase the risk of CVD and T2DM and cause mortality. Clinical identification and management of metabolic syndrome patients is fundamental to adequately reduce the risk of subsequent diseases. 14 Given the rising incidence, the elevated health costs, and the mortality associated with this disorder, the molecular and cellular pathogenesis of metabolic syndrome is of great interest. 15 Molecular mechanisms underlying metabolic syndrome pathogenesis are not clearly disclosed and many experimental animal models have been used to elucidate them. 16
Both metabolic syndrome and HHcy represent independent risk factors for the development of CVD. Moreover, metabolic syndrome and HHcy are often associated and seem to interact in increasing CVD risk, more than the two disorders can do alone. 17 Because NNMT plays a crucial role in Hcy metabolism, the aim of the present investigation was the analysis of the potential involvement of this enzyme in metabolic syndrome.
NNMT expression levels were evaluated by real-time PCR, immunohistochemistry, western blot analysis, and catalytic activity assay in adipose tissue from WOKW rats, which develop a complete metabolic syndrome closely resembling the human disease, and from control DA rats. Results obtained showed that NNMT mRNA and protein levels were statistically significantly higher in adipose tissue from WOKW rats compared with those observed in adipose tissue of DA rats. Moreover, a marked increase in NNMT enzymatic activity was detected in tissue specimens from WOKW rats than those obtained from DA rats.
NNMT is a SAM-dependent cytosolic enzyme that catalyzes the N-methylation of nicotinamide. 18 NNMT is highly expressed in the liver, 8 and its upregulation has been reported in several malignancies, including clear cell renal cell carcinoma (ccRCC), 19 oral squamous cell carcinoma (OSCC), 20 –24 bladder urothelial carcinoma (BUC), 25 and non-small cell lung cancer (NSCLC). 26,27 Moreover, NNMT was found to be overexpressed in Parkinson disease, 28 atherosclerosis, 29 and chronic obstructive pulmonary disease (COPD). 30
Regarding the expression of NNMT in adipose tissue and the enzyme contribution to Hcy production, Riederer et al. reported very interesting results. They showed that murine and human adipose tissue displayed elevated NNMT expression levels, even higher than those observed in the liver, which was previously considered the main NNMT-containing organ. NNMT mRNA and catalytic activity levels were also significantly high during the differentiation of 3T3-L1 fibroblasts into adipocytes. Moreover, both progressive differentiation of these cells and incubation of murine adipose tissue with nicotinamide led to increased Hcy release. 31 All of these data demonstrated that adipose tissue may be an important source of both NNMT and Hcy. In the light of these considerations, our results are consistent with the hypothesis that NNMT could make a major contribution to the determination of the elevated plasma Hcy levels associated with metabolic syndrome. It would be interesting to evaluate plasma Hcy levels in WOKW and DA rats to explore whether or not there is a statistically significant correlation with NNMT expression levels in adipose tissue.
Although NNMT overexpression has been observed in several pathologic conditions, a few studies investigated the functional significance of this alteration. Kim et al. explored the biological role of the enzyme in myoblasts and found that NNMT upregulation triggered a significant increase in cell proliferation and migration, and reduced cell death induced by reactive oxygen species (ROS) exposure. 30 Because metabolic syndrome is often characterized by oxidative stress associated with ROS production, 32 the NNMT overexpression observed in adipose tissue from WOKW rats could represent a protective mechanism sustaining the pathogenesis of this metabolic disorder, counteracting the oxidative damage related to ROS production. A very recent paper showed that NNMT knockdown in adipose tissue and liver of high-fat diet (HFD)-fed mice led to a reduction of fat mass, an increase of insulin sensitivity, and a decrease of serum TGs and free fatty acid levels. Leanness associated with NNMT downregulation is mainly due to an enhancement of energy expenditure, which was demonstrated by increased oxygen consumption during a comprehensive laboratory animal monitoring system (CLAMS). Taken together, these findings strongly indicate that NNMT knockdown has a protective effect against the deleterious metabolic consequences of diet-induced obesity. 33 It is our future aim to evaluate the effect of NNMT knockdown in metabolic syndrome to clarify the role played by the enzyme in the etiology of the disease and to raise the possibility of using NNMT as an attractive target for the treatment of the disorders associated with metabolic syndrome.
To our knowledge, this is the first study to evaluate NNMT expression levels in metabolic syndrome. Although preliminary, results revealed NNMT overexpression in adipose tissue of a rat model of metabolic syndrome. These results seem to suggest that NNMT may play an important role in the pathogenesis of metabolic syndrome and could have a great potential for the development of a targeted therapy.
Footnotes
Acknowledgment
We are indebted to Dr. Luca Tiano (Polytechnic University of Marche) for his assistance in collecting rat tissues.
Author Disclosure statement
No competing financial interests exist.