
Abstract
Background:
We aimed to investigate the effects of chronic oral treatment with centrally acting antihypertensive drugs, such as clonidine (CLO), an α2-adrenoceptor agonist, or LNP599, a selective I1 imidazoline receptor agonist, on brain microvascular function in rats with high-fat diet (HFD)-induced metabolic syndrome.
Methods:
Male Wistar Kyoto rats were maintained on a normal diet (CON) or a HFD for 20 weeks. After this period, the HFD group received oral CLO (0.1 mg/kg), LNP599 (20 mg/kg), or vehicle daily for 4 weeks. Systolic blood pressure and heart rate (HR) were evaluated by photoplethysmography. Functional capillary density, endothelial function, and endothelial–leukocyte interactions in the brain were investigated by intravital video microscopy. Cerebral microcirculatory flow was evaluated by laser speckle contrast imaging. Brain tissue endothelial nitric oxide synthase, oxidative enzyme, and inflammatory marker expression levels were analyzed.
Results:
Metabolic syndrome decreased brain functional capillary density and microvascular blood perfusion, changes accompanied by deficient brain microcirculation vasodilatory responses to acetylcholine. Significant numbers of rolling and adherent leukocytes were also observed in the brain venules. Chronic sympathetic inhibition with clonidine and LNP599 reduced blood pressure and HR. These effects were accompanied by reversals of cerebral capillary rarefaction, improvements in cerebral microvascular blood flow and endothelial function, and decreases in endothelial–leukocyte interactions in the cerebral venules.
Conclusions:
Our results suggest that central sympathetic inhibition exerts beneficial effects by increasing perfusion and reducing inflammatory marker expression and oxidative stress in the brains of rats with metabolic syndrome. Centrally acting antihypertensive drugs may be helpful in regulating cerebral microcirculatory function and vascular inflammation in metabolic syndrome.
Introduction
T
Sympathetic hyperactivity and visceral obesity play key roles in the inflammatory process by increasing plasma catecholamine and proinflammatory cytokine concentrations and by attenuating vagal stimulation. 3,4 Previous studies have demonstrated that acetylcholine can suppress proinflammatory cytokine production in vitro and stimulate vagal nerve activity in vivo, thereby preventing inflammation. 5,6 Visceral adipose tissue seems to play a central role in chronic inflammation by releasing proinflammatory factors, such as tumor necrosis factor-α and interleukin-6; chemotactic factors for type 1 macrophages; and vasoconstrictive factors, such as angiotensin II. All of these factors can reach the circumventricular organs (CVOs) and worsen neuroinflammation. 7
The CVOs are a group of small areas surrounding the ventricular system of the central nervous system. A very permeable capillary network supplied by fenestrated capillaries enables the CVOs to remain in close contact with the rest of the organism via the blood circulation. The lack of a blood–brain barrier, as well as a peculiar ultrastructure, enables the CVOs to interact with the rest of the organism via neuroreceptors, facilitating their involvement in neuroendocrine processes. Previous studies have demonstrated that some CVO areas, such as the subfornical organ, are important peripheral inflammation sensors mediating the influence of circulating proinflammatory cytokines on autonomic and cardiovascular function. 8
Endothelial dysfunction is another feature of metabolic syndrome and is related to oxidative stress. Some metabolic syndrome features, such as low high-density lipoprotein levels, high low-density lipoprotein levels, arterial hypertension, and increased free fatty acid concentrations, are independent risk factors for atherosclerosis, which directly injures the endothelium. 9 Obesity is associated with chronic cerebral hypoperfusion, which results from decreased endothelium-derived nitric oxide synthesis and activity and leads to cognitive decline and neurodegenerative effects mediated by oxidative stress. 10
In the sympathetic nervous system, efferent signals traveling from the brain to the peripheral tissues are orchestrated by several forebrain and midbrain nuclei, such as the hypothalamic paraventricular region and the CVOs, the rostral ventrolateral medulla, the anteroventral third ventricle, and the solitary tract nucleus. Variations in blood flow within these areas may contribute to arterial hypertension and increase peripheral catecholamine release. 11 The concept of vascular–neuronal signaling demonstrating the relationship between hypertension and vascular inflammation within the brainstem has been described previously. Leukocyte accumulation within the microvasculature of the solitary tract nucleus in spontaneously hypertensive rats can reduce cerebral blood flow and increase sympathetic activation. 12
It was recently shown that central sympatho-inhibitory drugs, such as clonidine and rilmenidine, improve skeletal and cardiac muscle microcirculation in rats with metabolic syndrome. 13,14 The effects of these centrally acting antihypertensive drugs are mediated by α2-adrenergic and I1 imidazoline receptors. The effects of clonidine have not been studied in patients with metabolic alterations, as clonidine was originally developed as an antihypertensive drug. Other drugs of the same pharmacological class, designated as second- and third-generation agents, were later developed and found to exert positive effects on abnormal metabolic conditions, such as insulin resistance, impaired glucose tolerance, and dyslipidemia, in humans 15 –17 and in experimental models involving rats. 14,18
A newly synthesized compound, LNP599, 18 which has a high affinity for imidazoline receptors and no detectable affinity for α2-adrenoceptors and is mainly indicated for its beneficial metabolic effects, was recently suggested as a new target for hypertension and metabolic syndrome management. 19
Thus, this study aimed to investigate the impact of centrally acting antihypertensive drugs on high-fat diet (HFD)-induced metabolic syndrome-related cerebral microcirculation alterations in rats.
Materials and Methods
HFD-induced metabolic syndrome in rats
All experiments were conducted in accordance with internationally accepted principles for the care and use of laboratory animals and were approved by the Oswaldo Cruz Foundation Animal Welfare Committee (protocol No. P 0034-08). We used an experimental metabolic syndrome model involving Wistar rats fed a HFD supplemented with salt, as previously described. 20 This standardized model induces marked increases in visceral adipose tissue, serum cholesterol, and triglyceride levels and insulin resistance.
The experiments were performed on male Wistar Kyoto rats (Oswaldo Cruz Foundation Animal Facilities) housed under controlled light (12:12 hrs of light–dark cycles) and temperature (22°C ± 1°C) conditions and allowed access to food and water ad libitum. The animals were studied in the fed state.
At 4 weeks of age, the rats were randomly divided into two groups that were fed standard chow (Nuvilab-CR1; Nuvital Nutrients Ltd., Colombo, Brazil) (CON, n = 10) or a HFD (n = 30) for 20 weeks. Then, the HFD group received clonidine (CLO; 0.1 mg/kg), LNP599 (20 mg/kg), or vehicle (distilled water, VEH) by gavage once daily for 4 weeks, a period of time equivalent to ∼2 years in the life of a human, according to the literature. 21 To exclude the direct effects of calorie restriction, the CON and HFD were maintained during pharmacological treatment. The composition of the commercial standard chow (Nuvilab-CR1; Nuvital Nutrients Ltd.) used in the present study was consistent with the American Institute of Nutrition guidelines, revised in 1993, for rats of this age.
The HFD comprised standard chow supplemented with corn starch, condensed milk, saturated animal fat (lard), and 0.5% NaCl. Regarding micronutrient composition, the CON diet contained 23% protein, 71% carbohydrate, 6% lipid, and 1.3% NaCl (4300 kcal/kg), and the HFD contained 14% protein, 56% carbohydrate, 30% lipid, and 0.5% NaCl (5443 kcal/kg). In this experimental model of metabolic syndrome, the HFD animals had a larger intake of calories than animals fed standard rat chow. 20 Blood samples were collected by cardiac puncture at the end of the experimental protocol. The plasma was separated via centrifugation at 3000 rpm for 15 min at 4°C, and the aliquots were stored at −80°C until measurement.
Hemodynamic measurements
Systolic arterial pressure and heart rate (HR) were assessed in each animal via weekly noninvasive tail-cuff plethysmography measurements (BP-2000, Visitech Blood Pressure Analysis System; Visitech Systems, Inc., Apex, NC), the last of which were performed on the last day of treatment.
Intravital video microscopy
The rats were anesthetized with a mixture of xylazine and ketamine hydrochloride (200 and 10 mg/kg, intraperitoneal, respectively). The jugular vein was cannulated to inject fluorescent tracers and additional anesthesia. Core body temperature was monitored with a rectal probe and maintained at 37°C with a homeothermic blanket system (Harvard Apparatus, Cambridge, United Kingdom).
Intravital microscopy was performed as previously described. 22 Briefly, the animals were fixed in a stereotaxic frame, the left parietal bone was exposed by a midline skin incision, a cranial window overlying the right parietal bone (1–5 mm lateral, between the coronal and lambdoid sutures) was created with a high-speed drill, and the dura mater and arachnoid membranes were excised and withdrawn to expose the cerebral microcirculation. The cranial window was suffused with artificial cerebrospinal fluid (in mmol: NaCl, 132; KCl, 2.95; CaCl2, 1.71; MgCl2, 0.64; NaHCO3, 24.6; dextrose, 3.71; and urea, 6.7; at 37°C, pH 7.4). This procedure does not cause changes in permeability under baseline conditions.
The animals were then placed under an upright fixed-stage intravital microscope equipped with a mercury lamp (Olympus BX51/WI) coupled to a charge-coupled device (CCD) digital video camera system (Optronics, Tokyo, Japan). Olympus 10× and 20× objectives were used in the experiments and produced total magnifications of 100× and 200× , respectively.
Capillary density and leukocyte–endothelial interaction evaluation
Brain microvessel visualization was facilitated by intravenous (IV) administration of 0.1 mL of 2% fluorescein isothiocyanate (FITC)-labeled dextran (molecular weight 150,000) and by epi-illumination at 460–490 nm using a 520-nm emission filter.
To analyze the interactions between leukocytes and the endothelium, leukocytes were labeled via IV injections of rhodamine 6G (0.3 mg/kg IV), and leukocyte fluorescence was visualized on postcapillary venules (different regions of capillary analysis) via epi-illumination at 510–550 nm using a 590-nm emission filter. Five randomly selected venular segments (30–100 μm in diameter) in each preparation were observed for 30 sec to assess leukocyte recruitment. Leukocyte–endothelial interactions were evaluated by determining the number of leukocytes that adhered to 100 μm of the venular wall for a period of 30 sec. Leukocytes crossing the venular segment at a speed below that of circulating red blood cells were defined as rolling leukocytes; this rate was expressed as cells/min.
Cerebral blood flow analysis
We used laser speckle contrast imaging (LSCI; Perimed, Järfälla, Sweden) for cerebral blood flow analysis, as this recently developed technique provides microvascular perfusion indices proportional to the concentrations and mean velocities of red blood cells. LSCI is a noninvasive blood flow imaging technique and has been used mostly in clinical studies to assess microcirculation reactivity and to accurately quantify relative changes in skin perfusion. 23 This methodology allows measurements of cerebral blood flow with high temporal and spatial resolution and facilitates selected-field analyses and determinations of relative changes in cerebral blood flow.
After intravital microscopy, the open cranial windows of the anesthetized rats were positioned under a laser light (785 nm wavelength) at a fixed working distance of 10 cm. All animals were subjected to the cranial window procedure under intravital microscopy, followed by laser speckle analysis under appropriate conditions, to exclude the effects of time in the experimental protocol.
To analyze changes in cerebral blood flow, we defined regions of interest over the brain surface using images projected on a monitor. Care was taken not to disturb the preparations during image acquisition. Analyses of 16 laser speckle images per second and relative cerebral blood flow were performed for all animal groups over a period of 20 min using Perisoft software (Perimed), and the results were expressed as arbitrary perfusion units.
Cerebral microvascular reactivity to acetylcholine
Microvascular endothelial function was evaluated based on endothelium responses to topical application of the endothelium-dependent vasodilator acetylcholine (Ach, 10−6 M). The cranial window was suffused with Ach for 5 min, and cortical blood flow variations were measured via LSCI before and after vasoactive substance exposure. Microvascular blood flow responses were expressed as percentages (%) of variation from baseline. After intravital microscopy and laser speckle analysis, the animals were killed with pentobarbital overdoses, in accordance with ethics committee specifications, and brain and blood samples were collected for subsequent molecular and biochemical analyses.
Reverse transcription-polymerase chain reaction analysis of endothelial nitric oxide synthase and NADPH oxidase expression
Total RNA was prepared from rat right brain hemisphere samples using an RNeasy Mini Kit (Qiagen, Hilden, Germany), according to the manufacturer's instructions. Complementary DNA (cDNA) was synthesized from 1 μg of total RNA in a final volume of 20 μL using a High-capacity cDNA Reverse Transcription Kit (Applied Biosystems). The following primer sequences were used for polymerase chain reaction (PCR) amplification: endothelial nitric oxide synthase (eNOS), forward: 5′-GTATTTGATGCTCGGGACTG-3′ and reverse: 3′-AGATTGCCTCGGTTTGTTG-5′; NADPH oxidase p47 subunit (phox), forward: 5′-GTGAAGCCATCGAGGTCATTC-3′ and reverse: 3′-CCCGCGGCTTCTAATCTGT-5′; and beta-actin (β-a): forward: 5′-CCACCCGCGAGTACAACCTTCTT-3′ and reverse: 3′-GAAGCCGGCCTTGCACATGCC-5′.
DNA sequences were chosen using the NCBI/Nucleotide database and then converted using the Primer 3 program (Rozen and Skaletsky, 2000). PCR amplification of the resulting cDNA template was conducted for 35 cycles under the following conditions. After an initial denaturation step at 95°C for 15 min, temperature cycling was initiated. Each cycle consisted of denaturation at 94°C for 30 sec, annealing at 60°C for 30 sec, and elongation at 72°C for 30 sec. The PCR products were analyzed on 6% polyacrylamide gel and stained via silver dying. Images were captured and analyzed using ImageJ software (National Institute of Health).
Western blotting
Cerebral proteins were isolated from the left hemisphere of the brain (100 mg/mL) homogenates and suspended in phosphate-buffered saline containing 0.4 M sodium chloride, 0.05% Tween 20, and protease inhibitors (0.1 mM phenylmethylsulfonyl fluoride and 1/100 protease inhibitors cocktail; Sigma).
The samples were sonicated twice and centrifuged for 10 min at 3000g. The supernatant was subsequently frozen at −80°C. The proteins in the lysates were separated by sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS/PAGE; 20 μg/lane) on 10% gels. Immunoblotting was performed with the following specific primary antibodies: rabbit anti-eNOS (130 kDa), rabbit anti-vascular cell adhesion molecule-1 (anti-VCAM-1; 100 kDa), and rabbit anti-intercellular adhesion molecule-1 (anti-ICAM-1; 52 kDa; Abcam). Mouse anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH, 36 kDa) monoclonal antibodies were used as loading controls. The membranes were incubated overnight at 4°C with the indicated specific primary antibodies before being incubated with the secondary goat anti-rabbit IgG or secondary goat anti-mouse IgG horseradish peroxidase (HRP)-labeled antibodies for 1 hr at 25°C.
The blots were ultimately incubated with Super Signal Chemiluminescence substrates (Thermo Scientific) or Westar Supernova substrates (Cyanagen, Bologna, Italy).
Drugs
FITC-labeled dextran, rhodamine 6G, clonidine, and Ach were obtained from Sigma Chemical Co. (St. Louis, MO); ketamine was obtained from Cristália; and xylazine was obtained from Syntec. LNP599 was synthetized and provided by the Laboratory of Neurobiology and Cardiovascular Pharmacology, Faculty of Medicine, University of Strasbourg (Strasbourg, France).
Statistical analysis
All data for each group are expressed as the mean ± standard error of the mean, and comparisons among the groups were performed with one-way analysis of variance (ANOVA). When an overall difference was detected by ANOVA, the Bonferroni test was used to localize statistically significant differences. Differences with P values <0.05 were considered significant. All calculations were performed using computer-based statistical software (GraphPad 5.0; GraphPad Software, Inc., La Jolla, CA).
Results
Hemodynamic and metabolic effects of central sympathetic inhibition on HFD-induced metabolic syndrome in rats
The HFD+VEH group exhibited increased systolic blood pressure levels compared to the CON+VEH group after the HFD period and 4 weeks of vehicle treatment. The blood pressure increases observed in the HFD+VEH group were reversed by the centrally acting antihypertensive drugs LNP599 and CLO. Similarly, the HFD+VEH group exhibited an increased HR compared with the CON+VEH group, and this increase was also reversed by LNP599 and CLO (Table 1).
Body weight, SAP, HR, fasting glucose and insulin levels of rats with metabolic syndrome. Values represent the mean ± standard error, n = at least 8 per group.
p < 0.05, b p < 0.01, c p < 0.001 versus CON VEH; d p < 0.05, e p < 0.01 versus HFD VEH.
CLO, clonidine; HR, heart rate; HFD, high-fat diet; SAP, systolic arterial pressure.
The HFD+VEH group exhibited significantly increased fasting blood glucose levels compared with the CON+VEH group. However, only the HFD+LNP599 group exhibited significantly reduced glucose levels compared with the HFD+VEH group. The HFD+CON group exhibited reduced glucose levels compared with the HFD+VEH group, although glucose levels remained elevated in this group compared to the control group (Table 1).
Insulin levels were highly increased in the HFD+VEH group and were normalized in the HFD+LNP599 and HFD+CLO groups (Table 1). No effects on body weight were observed in any of the groups after treatment (Table 1).
Effects of sympathetic inhibition on the cerebral microcirculation of rats with HFD-induced metabolic syndrome
To investigate the protective effects of sympathetic inhibition in an experimental model of metabolic syndrome, we performed intravital microscopic examinations at the end of the experimental protocol. Figure 1 and Supplementary Figure S1 (Supplementary Data available online at
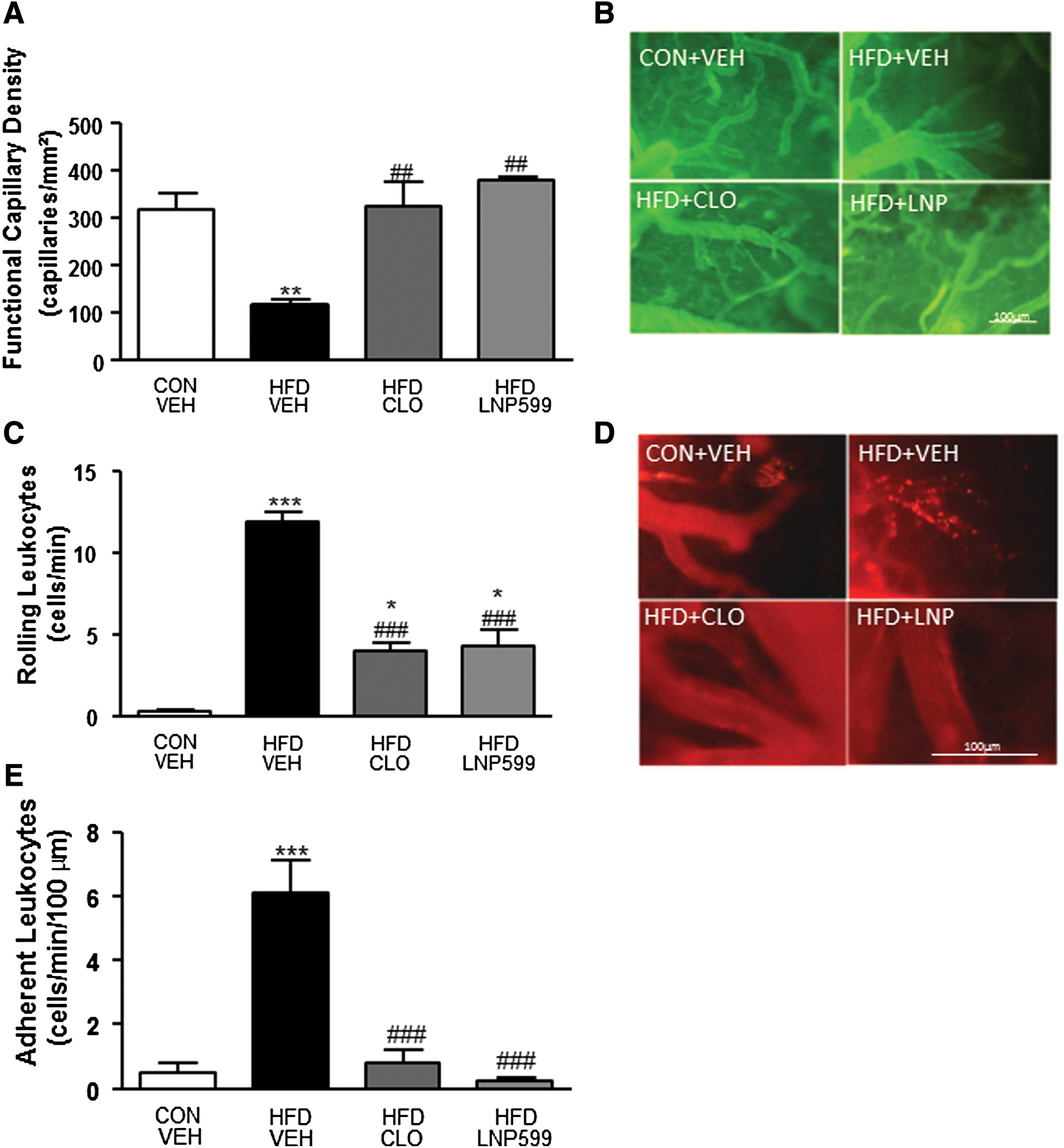
Functional capillary density and leukocyte-endothelium interactions in the brain microcirculation of rats with metabolic syndrome treated with centrally acting antihypertensive drugs or vehicle. Brain functional capillary density
Intravital fluorescence microscopic examinations of the cerebral venules of HFD+VEH rats revealed a significant increase in the numbers of rolling and firmly adherent leukocytes in these rats compared with CON+VEH rats. The HFD+CLO and HFD+LNP599 groups exhibited lower numbers of rolling leukocytes than the HFD+VEH group, but higher numbers of rolling leukocytes than the CON+VEH group. CLO and LNP599 treatment significantly blunted the recruitment of adherent leukocytes into the cerebral microvasculature (Figs. 1 C, D and E).
Laser speckle imaging evaluation of brain perfusion in rats with HFD-induced metabolic syndrome
Microvascular brain perfusion was significantly reduced in the brains of rats in the HFD+VEH group compared with those in the CON+VEH group, whereas there was no difference in microvascular brain perfusion between the treated groups and the control group (Fig. 2A and Supplementary Figure S1).

Brain microvascular blood flow and endothelial function in rats with metabolic syndrome treated with centrally acting antihypertensive drugs or vehicle. Brain microvascular blood flow
LSCI assessments of cerebrovascular function revealed that microvascular vasodilator responses to ACh were blunted in the HFD+VEH group (Fig. 2B, C) compared with the CON+VEH group, whereas vasodilator responses to Ach were restored or even potentiated in the HFD+CLO and HFD+LNP599 groups (Fig. 2B).
Effects of sympathetic activity on cerebral NADPH oxidase and eNOS gene expression in metabolic syndrome rats
Real-time PCR was used to assess NADPH oxidase p47 subunit and eNOS enzyme gene expression (Fig. 3).

Evaluation of oxidative stress and e-NOS in rats with metabolic syndrome treated with centrally acting antihypertensive drugs or vehicle. Real-time PCR analyses of the mRNA transcript levels of genes coding for the NADPH oxidase p47 subunit
We observed a significant increase in the expression of the NADPH oxidase subunit p47phox in the brains of HFD+VEH rats; however, we observed no difference in the expression of p47phox between the HFD+CLO and HFD+LNP599 groups and the control group (Fig. 3A).
As shown in Fig. 3B, the HFD exerted no significant effects on brain eNOS expression in either the HFD+CLO group or the HFD+LNP599 group.
Western blot analysis of eNOS, VCAM-1, and ICAM-1 protein expression in the brains of rats with metabolic syndrome
A marked decrease in eNOS expression was observed in the HFD+VEH group compared with the CON+VEH group. This effect was reversed in the HFD+CLO group (Fig. 4). LNP599 treatment did not affect the abovementioned HFD-induced reductions in eNOS expression.

eNOS content in the brains of rats with metabolic syndrome treated with centrally acting antihypertensive drugs or vehicle. Western blot analysis of eNOS protein expression in the brains of rats fed a normal (CON) or HFD after long-term oral treatment with the centrally acting drugs clonidine (HFD+CLO, 0.1 mg/kg) or LNP599 (HFD+LNP599, 20 mg/kg) or vehicle (HFD+VEH). Values represent the mean ± SEM, n = 5–8 for each group. *p < 0.05 versus the CON+VEH group; # p < 0.05 versus HFD+CLO; § p < 0.05 versus HFDCLO. eNOS, endothelial nitric oxide synthase.
The HFD induced an increase in VCAM-1 protein expression, and sympathetic nervous system inhibitor treatment was not able to reverse this increase (Fig. 5A). We also observed a significant increase in ICAM-1 expression in the HFD+VEH group compared with the CON+VEH group. There was no difference in ICAM-1 expression between the HFD+CLO and HFD+LNP groups and the CON+VEH group.
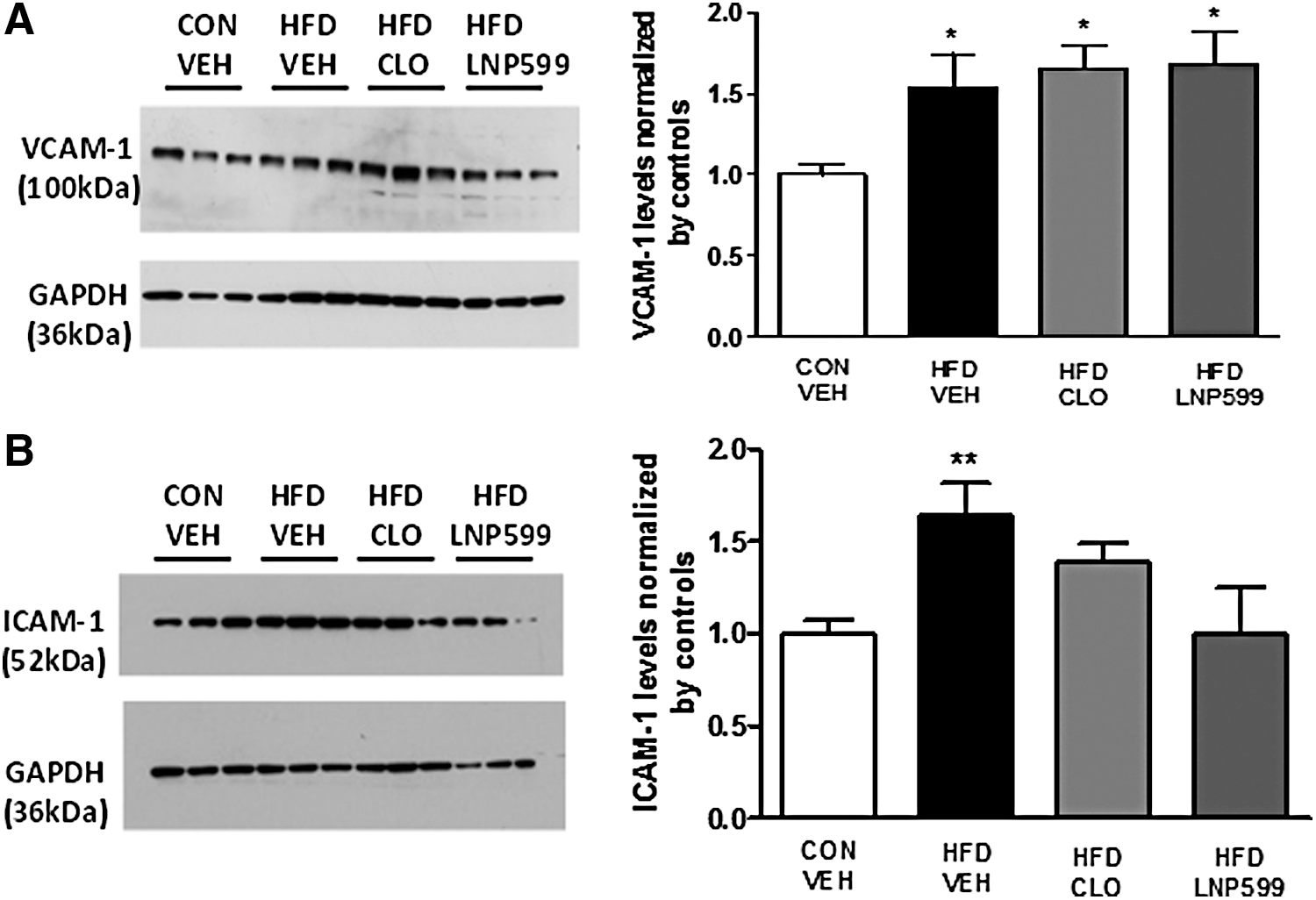
Evaluation of the expression of the vascular inflammation markers VCAM-1 and ICAM-1 in rats with metabolic syndrome treated with centrally acting antihypertensive drugs or vehicle. Expression levels of VCAM-1 (100 kDa)
Discussion
The major findings of the present study were as follows: (1) metabolic syndrome induces brain microvasculature alterations, such as capillary rarefaction, and also induces microvascular endothelial dysfunction, increases in oxidative stress, vascular inflammation, and reductions in microvascular perfusion; and (2) centrally acting sympathetic inhibitors are able to improve metabolic syndrome-induced brain microcirculation alterations.
Capillary integrity is an important determinant of brain function. 24 The vasculature of the pia mater membrane forms a complex vascular network on the brain surface that nourishes the surface layers of the cerebral cortex. 25 Pathological changes in the vascular wall and alterations in metabolic factor release can profoundly influence cerebral vascular autoregulation responses. Atherosclerosis and/or reductions in vasodilation in the arterioles and small vessels of the microcirculation lead to brain hypoperfusion, which causes subsequent reductions in capillary flow accompanied by leukocyte interactions with postcapillary venule endothelial walls. This process compromises vascular endothelial cell integrity, allowing macromolecule extravasation into the extravascular space. 26
Intravital microscopy has been extensively used for studying dynamic processes in cardiovascular diseases. This technique allows analysis of microcirculatory function in the brain and quantification of the inflammatory processes of leukocyte rolling and adhesion to venular walls.
In recent years, microvascular reactivity assessments have proven to be essential for investigating the pathophysiology of cardiovascular diseases. LSCI identifies endothelium-dependent and endothelium-independent microvascular dysfunction in patients and in experimental animal models and has thus served as a valuable tool for studying atherothrombotic disease. 27,28
We confirmed that arterial blood pressure and HR are increased in rats with HFD-induced metabolic syndrome. These hemodynamic effects are accompanied by increases in fasting blood glucose and insulin levels. Enhanced sympathetic nervous system activity is a common feature of metabolic syndrome. 29 Long-term oral treatment with the sympatho-inhibitory compounds clonidine and LNP599 was able to reverse these hemodynamic and metabolic outcomes. The pronounced bradycardic effects of clonidine-like drugs are mediated exclusively by α2-adrenergic receptors located in the nucleus tractus solitarius and nucleus ambiguus 30,31 and have also been attributed to reductions in catecholamine release in noradrenergic nerve terminals. 32 It was proposed previously that insulin resistance and weight gain are the first consequences of increased sympathetic activity in obese adults. 33 Prolonged adrenergic stimulation can lead to beta-adrenergic receptor desensitization, preventing energy dissipation following food consumption and potentiating obesity by facilitating increases in visceral adipose tissue and insulin resistance. 34
HFD consumption induces functional capillary rarefaction in the brain accompanied by increased vascular inflammation characterized by significant amounts of leukocyte rolling and adhesion to venular walls. Overexpression of ICAM-1, a vascular biomarker of endothelial dysfunction, 35 was reduced in the brains of rats with metabolic syndrome that were treated with clonidine and LNP599. Therefore, our results imply that the vascular anti-inflammatory effects of central sympathetic inhibition are accomplished, in part, by reducing ICAM-1 expression, but not VCAM-1 expression. ICAM is mainly involved in firm leukocyte adhesion to small vessels and may be associated with microvasculature plugging by inflammatory cells, as observed in some venular segments of the cerebral microcirculation, leading to impaired microvascular drainage. Thus, based on these observations, we believe that all of the microcirculatory alterations in the brain cortices of animals with metabolic syndrome are related to the brain hypoperfusion observed via LSCI.
NADPH oxidases are important sources of reactive oxygen species (ROS) and have been implicated in numerous neurodegenerative processes. 36 Previous studies have shown that HFD feeding is a well-established model of obesity that reproduces key features of obesity in humans, including marked increases in visceral adipose tissue and oxidative stress. 20,37 The superoxide-producing NADPH oxidase, which is similar to the enzyme complex found in phagocytic leukocytes, is present in vascular tissues. This phagocytic enzyme complex has at least five components as follows: two cytosolic subunits, p47phox and p67phox, and a cell membrane-bound protein, cytochrome b558, which consists of gp91phox (renamed Nox2), p22phox, and a small G protein, Rac. 38
Nox2-derived superoxide is involved in brain endothelial dysfunction, as demonstrated previously in a model of diet-induced obesity, and seems to appear in cerebral arterioles earlier than in carotid arteries, 39 contributing to alterations in cerebral blood flow. The most important event in NADPH oxidase activation is p47phox (NOXO2, the organizer subunit of Nox2) activation. The catalytic activity of this enzyme depends on the phosphorylation (serine) of its cytoplasmic p47phox subunit, which justifies the investigations of gene expression in our work. We noted increased NADPH oxidase expression in the brains of animals treated with HFDs, suggesting that the oxidative stress present in metabolic syndrome is related to the long-term effects of high-fat consumption.
It has also been shown that prolonged exposure to ROS can damage insulin receptors and decrease glucose transporter expression, interfering with glucose uptake 40 and decreasing glucose transporter expression. 41 In our study, NADPH oxidase gene expression was significantly reduced in the brains of animals treated with centrally acting sympathetic inhibitors, suggesting that central sympathetic hyperactivity is involved in the oxidative stress associated with metabolic syndrome.
Increases in oxidative stress, particularly increases associated with superoxide anions, limit nitric oxide bioavailability. NO produced by eNOS reacts with superoxide to produce the reactive species peroxynitrite and suppress NADPH oxidase activation. Reduced NO bioavailability favors increased endothelin-1 or endothelium-derived contracting factor activity and promotes endothelial cell growth and vasoconstriction and thus participates in the pathogenesis of oxidative stress in arterial hypertension. 42
In the present study, eNOS protein levels were reduced in the brains of metabolic syndrome rats, findings consistent with those of studies involving animal models of obesity. 43 eNOS expression was downregulated in nontreated HFD animals; however, it was enhanced in clonidine-treated animals because eNOS protein levels in these animals were equal to those of control animals, corroborating the findings of studies involving cardiac myocytes 44 and endothelial cells 45 and showing that endothelial α2-adrenoceptors can mediate nitric oxide synthesis via the α2-adrenoceptor activating pathway.
Taken together, these findings indicate that the brain microvascular effects evoked by clonidine are also related to increases in eNOS production mediated by α2-adrenoceptors, corroborating the findings of studies demonstrating the beneficial effects of clonidine in inflammation. 46 However, LNP599 treatment does not increase eNOS production. According to the literature, NO synthase pathway activation is not mediated by I1 imidazoline receptors. 44,47 Therefore, the improvements in the cerebral microvasculature and the reductions in inflammation and oxidative stress induced by LNP599 treatment are probably facilitated by sympatho-inhibition mediated by I1 imidazoline receptors.
We believe that the beneficial effects exerted by central sympatho-inhibitory drugs on the brain microcirculation may also be due to reductions in insulin resistance and improvements in glycemic regulation, which in turn reduce oxidative stress and inflammation. Insulin exerts potent neuroprotective effects against apoptosis, beta-amyloid toxicity, and oxidative stress. In addition, hyperglycemic states render the brain susceptible to microvascular disorders, as oxidative stress affects vascular pericytes, pluripotent cells that preserve blood–brain barrier, and capillary bed integrity. 48 Therefore, insulin resistance is involved in the pathogenesis of neurodegenerative diseases, as individuals with diabetes mellitus have an increased risk of developing Alzheimer disease and some types of dementia. 49,50
The limitations of this study must be considered. We performed intravital microscopy and microvascular blood flow analysis on rat cerebral cortices, but all of these molecular analyses were completed using whole-brain samples. Although it is likely that alterations in the microcirculation of the cerebral cortex may correspond to changes in the brain parenchyma, additional studies should be performed to confirm this hypothesis.
The strengths of the present study include its promising results regarding the protective effects exerted by centrally acting sympatho-inhibitory drugs, namely, the attenuation of oxidative stress and endothelial dysfunction facilitated by these drugs.
In conclusion, our results suggest that HFD-induced metabolic syndrome induces cerebral microcirculation hypoperfusion associated with microvascular dysfunction and that targeting the central sympathetic activity may represent a valuable therapeutic strategy for treating metabolic syndrome.
Footnotes
Acknowledgments
This investigation was supported by grants from the CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico), the FAPERJ (Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro), and the Oswaldo Cruz Foundation (FIOCRUZ), Rio de Janeiro, Brazil.
Author Disclosure Statement
No conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.