
Abstract
Background:
Metabolic syndrome (MetS) facilitates the development of cardiovascular disease due to atherosclerosis, which is accelerated by defects of the vascular endothelium. Vascular dysfunction in response to nitric oxide (NO) occurs in the mesenteric arteries of an animal model of MetS, SHRSP.Z-Leprfa /IzmDmcr (SHRSP.ZF) rats. Vascular responses to vasodilators are affected by perivascular adipose tissue (PVAT) that surrounds the outsides of arteries. In this study, we assessed the role of PVAT in vascular dysfunction observed in SHRSP.ZF.
Methods:
To determine the effects of PVAT on vasodilators in SHRSP.ZF and control Wistar-Kyoto (WKY) rats, we used organ bath bioassay techniques to assay acetylcholine and nitroprusside-induced relaxations of isolated mesenteric arterial ring preparations with PVAT intact or removed.
Results:
A PVAT-mediated enhancement of relaxations induced by acetylcholine and nitroprusside occurred in SHRSP.ZF at 20 weeks of age, but not at 10 and 30 weeks, and did not occur in WKY. Furthermore, the enhancing effects of PVAT from SHRSP.ZF at 20 weeks could not be substituted by replacement with PVAT from either WKY or 30-week-old SHRSP.ZF, was inhibited by NO synthase inhibitor, and abolished by removal of the arteries' endothelium. Cyclic guanosine monophosphate (cGMP) accumulation elicited by nitroprusside was higher in SHRSP.ZF arteries with PVAT than arteries without PVAT at 20 weeks, but the enhancement of cGMP accumulation did not occur at 30 weeks.
Conclusions:
PVAT may regulate arterial tone by releasing diffusible vasorelaxing factor(s), which, through endothelium-derived NO production, compensates for impaired vasodilations at early stages of MetS.
Introduction
O
MetS is a chronic disease condition. The effects of aging and the duration of exposure to metabolic abnormalities on the function of PVAT and the nature of the mediators/factor(s) involved in PVAT's vasomotor functions are undetermined. We reported that chronic oxidative-nitrative stress is closely linked to vascular dysfunction in animal models of MetS, SHRSP.Z-Leprfa /IzmDmcr rats (SHRSP.ZF). 10,11 The SHRSP.ZF were established by breeding stroke-prone spontaneously hypertensive rats with Zucker obese rats. 12 SHRSP.ZF are used as an animal model of MetS for investigating the mechanisms of hypertrophy of adipocytes, 13 renal injury, 14 and cardiovascular diseases. 15,16 Therefore, in the present study, we assessed by using SHRSP.ZF whether PVAT effects are changed with increasing age and duration of exposure to metabolic abnormalities, and we investigated the regulatory mechanisms in vascular tone by PVAT.
Materials and Methods
Experimental animals
Male SHRSP.ZF and lean normotensive controls, Wistar-Kyoto (WKY) rats, were used in four studies. SHRSP.ZF and WKY were established by the Disease Model Cooperative Research Association (Hamamatsu, Japan). First, to assess the effects of age on PVAT regulation of vascular tone with increasing age, SHRSP.ZF were assigned to one of three groups according to their age (weeks) at the time of in vitro vasodilation studies: 10, 20, and 30 weeks (n = 10 each). Equal numbers of age-matched WKY were used. Next, to assess the mechanisms underlying PVAT's effects, a second set of SHRSP.ZF at 20 weeks (n = 10) was used. A third set of SHRSP.ZF at 20 and 30 weeks and WKY at 20 weeks was used to assess differences in effects of PVAT between strains (n = 10 each). A fourth set of SHRSP.ZF at 20 and 30 weeks was used to assess differences in effects of PVAT on Cyclic guanosine monophosphate (cGMP) accumulation elicited by nitroprusside (n = 5 each). The rats received regular feed (CE-2; Clea Japan, Inc., Tokyo, Japan) and were provided water ad libitum while they were housed in the animal care room. Rats were anesthetized with sodium pentobarbital (65 mg/kg, intraperitoneal) to obtain arteries and blood. All protocols were approved by the Animal Care Committee of Mukogawa Women's University.
Determination of metabolic parameters
Systolic blood pressure was measured by a tail-cuff method by using a blood pressure monitor (Model MK-2000; Muromachi Kikai Co., Tokyo, Japan) 1 week before the experimental date of vasodilation studies, as previously described. 17 On the day of the vasodilation studies, blood was drawn from the abdominal aortas of non-fasting rats, and the serum was separated by centrifugation at 1000g for 10 min at 4°C. Serum levels of triglyceride, glucose, insulin, and thiobarbituric acid reactive substances (TBARS) were determined by using commercial kits: the triglyceride E-test, glucose CII-test (Wako Pure Chemical Ind. Ltd., Osaka, Japan), the rat insulin detection kit (Morinaga Biochemistry Lab., Tokyo, Japan), and TBARS Assay Kit (Cayman Chemical Co., Ann Arbor, MI).
Vasodilation studies
Our experimental procedures for recording isometric tension with isolated blood vessels are described elsewhere in detail. 10 The superior mesenteric arteries were cleaned or not cleaned of surrounding PVAT, cut into ∼3-mm-wide rings, and mounted isometrically at an optimal resting tension of 0.3 g in 10 mL organ baths that were filled with a Krebs–Henseleit solution (pH 7.4) (in mM: NaCl 118.4, KCl 4.7, MgSO4 1.2, CaCl2 2.5, NaHCO3 25, KH2PO4 1.2, and glucose 11.1) and bubbled with 95% O2−5% CO2 at 37°C. For in vitro PVAT transplant experiments, cleaned arterial rings were mounted in organ baths and incubated with mesenteric arterial PVAT that was isolated from the same or comparative strain. This PVAT was cut into 50–70 mg pieces, cleaned by removing vascular tissue, tied to a stainless steel bar, and placed in close proximity to the arterial ring.
For each ring preparation, 1–3 μM phenylephrine was added to organ baths to produce a stable level of tone; then, relaxation responses were determined by addition of cumulative concentrations of acetylcholine or nitroprusside. In a separate series of experiments, the relaxation responses were determined under conditions of arterial pretreatments with a nitric oxide (NO) synthase inhibitor,
Determination of cGMP levels
Mesenteric arterial rings cleaned or not cleaned of surrounding PVAT of SHRSP.ZF were mounted under isometric conditions as described earlier for vasodilation studies. Each ring preparation was exposed to phenylephrine (1 μM) for 3 min and then treated with nitroprusside (0.01 μM) for 3 min. Then, after removal of any PVAT, the tissues were immediately frozen in liquid nitrogen and homogenized in a glass/glass homogenized in ice-cold 6% trichloroacetic acid. cGMP accumulations in arteries were measured by using an enzyme-linked immunoassay (Amersham Biotrak System; GE Healthcare, Buckinghamshire, United Kingdom) as previously described. 18 Protein concentration in each sample was determined by using a BCA protein assay kit (Pierce Biotechnology, Inc., Rockford, IL). cGMP data are reported as fmol cGMP per mg protein.
Data analysis
Data are expressed as mean ± standard error of the mean. n = number of animals. Statistical analysis was performed with one-way analysis of variance (ANOVA) or two-way ANOVA followed by Bonferroni post hoc test using GraphPad Prism. “Enhancing effect” represents the effect of PVAT on vasodilator responses, and it was calculated as differences in the relaxation (%) in response to 0.1 μM nitroprusside in mesenteric arteries between with PVAT and without PVAT in each animal. Linear regression analysis was performed by using GraphPad Prism. Differences between groups were considered significant if P < 0.05.
Results
Metabolic abnormalities in SHRSP.ZF
We assessed the metabolic dysfunction in each group of animals and compared the metabolic parameters of SHRSP.ZF and WKY used in the assessment of effects of PVAT on vascular tone with increasing age (n = 10 each at 10, 20, and 30 weeks). At 10, 20, and 30 weeks, SHRSP.ZF showed signs of abdominal obesity as evidenced by heavier body weight and larger waist length–body length ratio compared with age-matched WKY (Table 1). SHRSP.ZF had higher blood pressure, higher serum triglyceride, glucose, and insulin than those in WKY. The serum TBARS level, an index of oxidative stress, was higher in SHRSP.ZF than that in WKY at 30 weeks.
*P < 0.05, as compared with age-matched WKY
SHRSP.ZF, SHRSP.Z-Leprfa /IzmDmcr rats; WKY, Wistar-Kyoto rats; ANOVA, analysis of variance; TBARS, thiobarbituric acid-reactive substances; ns, not significant, n = 10 each strain at each age.
Effects of PVAT on relaxations in SHRSP.ZF and WKY
Figure 1 shows the effects of PVAT on endothelium-dependent relaxations induced by acetylcholine and endothelium-independent NO-mediated relaxations induced by nitroprusside in mesenteric arteries of SHRSP.ZF and WKY (n = 10 each strain). The responses to agonists in the presence or absence of PVAT were equivalent in mesenteric arteries of WKY and SHRSP.ZF at 10 weeks. In contrast, at 20 weeks, vascular dysfunction was present in mesenteric arteries without PVAT of SHRSP.ZF as compared with WKY, and the presence of PVAT in SHRSP.ZF enhanced relaxation responses by both agonists. This enhancement effect was lost in SHRSP.ZF at 30 weeks.
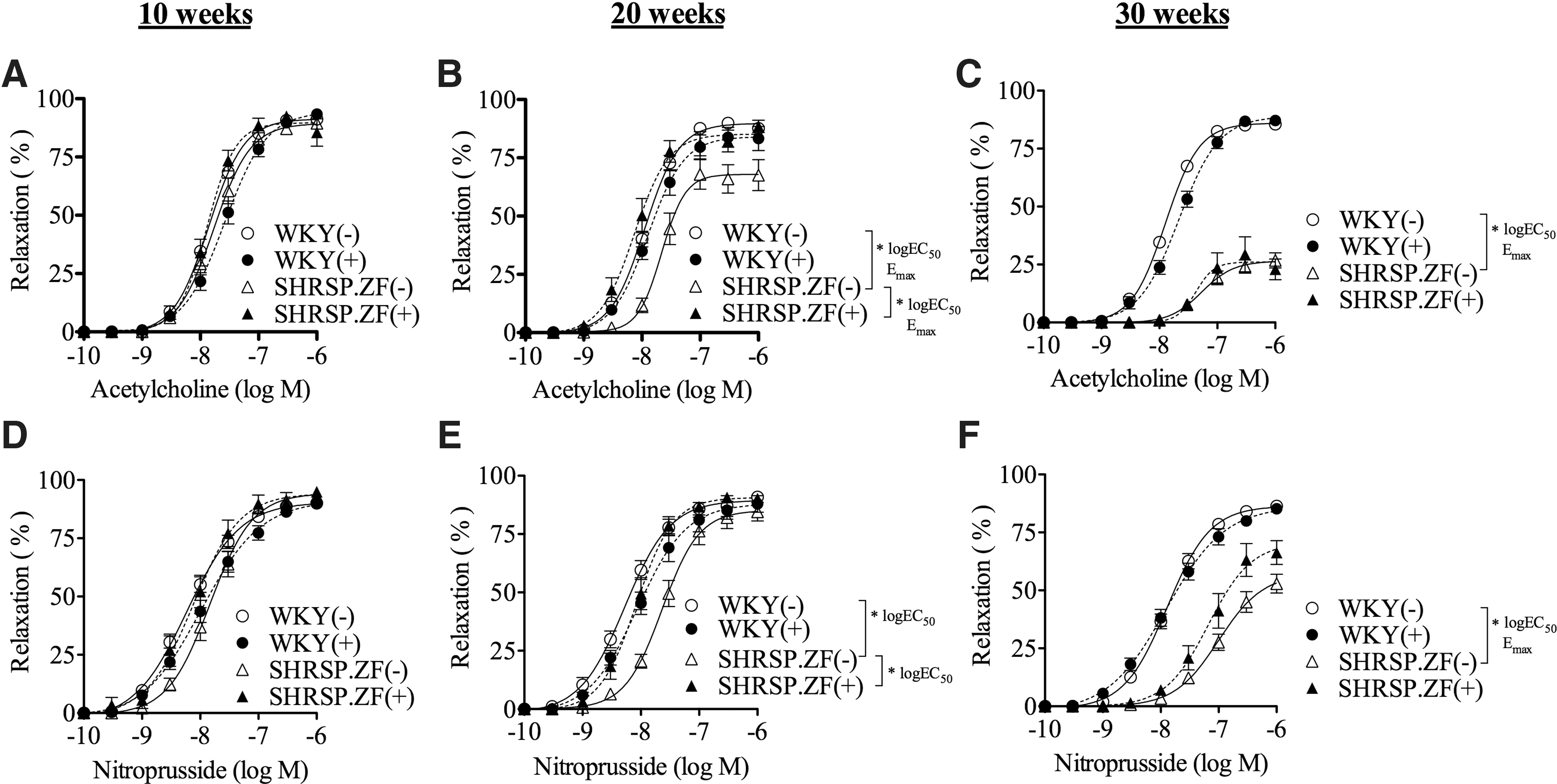
Vasodilation in response to acetylcholine
The enhancing effects of PVAT in SHRSP.ZF at 20 weeks were abolished by removing the arteries' endothelium (Fig. 2A), and they were completely inhibited by pretreating arteries with an NO synthase inhibitor (Fig. 2B) (n = 10). In addition, the PVAT-enhancing effect was negatively correlated to the relaxation in arteries without PVAT at 20 weeks (r = −0.929, P < 0.001).
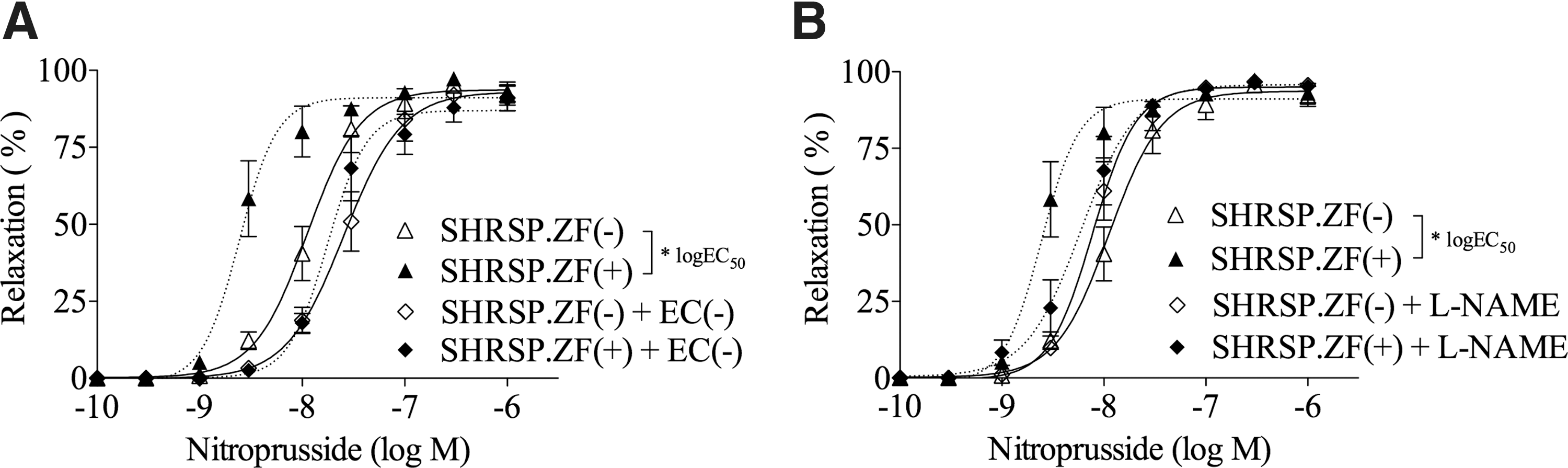
Effects of removing the endothelium [EC,
Using mesenteric arterial PVAT and arteries of WKY and SHRSP.ZF at 20 weeks (n = 10 each strain), we compared the effects of in vitro transplanting PVAT between SHRSP.ZF and WKY on the relaxations (Fig. 3). The relaxations induced by acetylcholine and nitroprusside in mesenteric arteries from WKY were not affected by the presence of PVAT from either strain (Fig. 3A, C). In contrast, the relaxations in SHRSP.ZF arteries were enhanced by the presence of PVAT from the same strain, and the relaxation percentages were at similar levels as those in the arteries enveloped by PVAT (Fig. 3B, D). However, the relaxations in SHRSP.ZF arteries were not affected by the presence of PVAT from WKY (Fig. 3B, D). Furthermore, when the effects of in vitro transplanting PVAT between 20-week age and 30-week age of SHRSP.ZF on the relaxation were compared (Fig. 4, n = 10 each age), the relaxations induced by nitroprusside in SHRSP.ZF arteries at 20 (Fig. 4A) and 30 weeks (Fig. 4B) were not affected by the presence of PVAT from 30-week age of SHRSP.ZF.

Effects of in vitro transplant of PVAT on vasodilations in response to acetylcholine
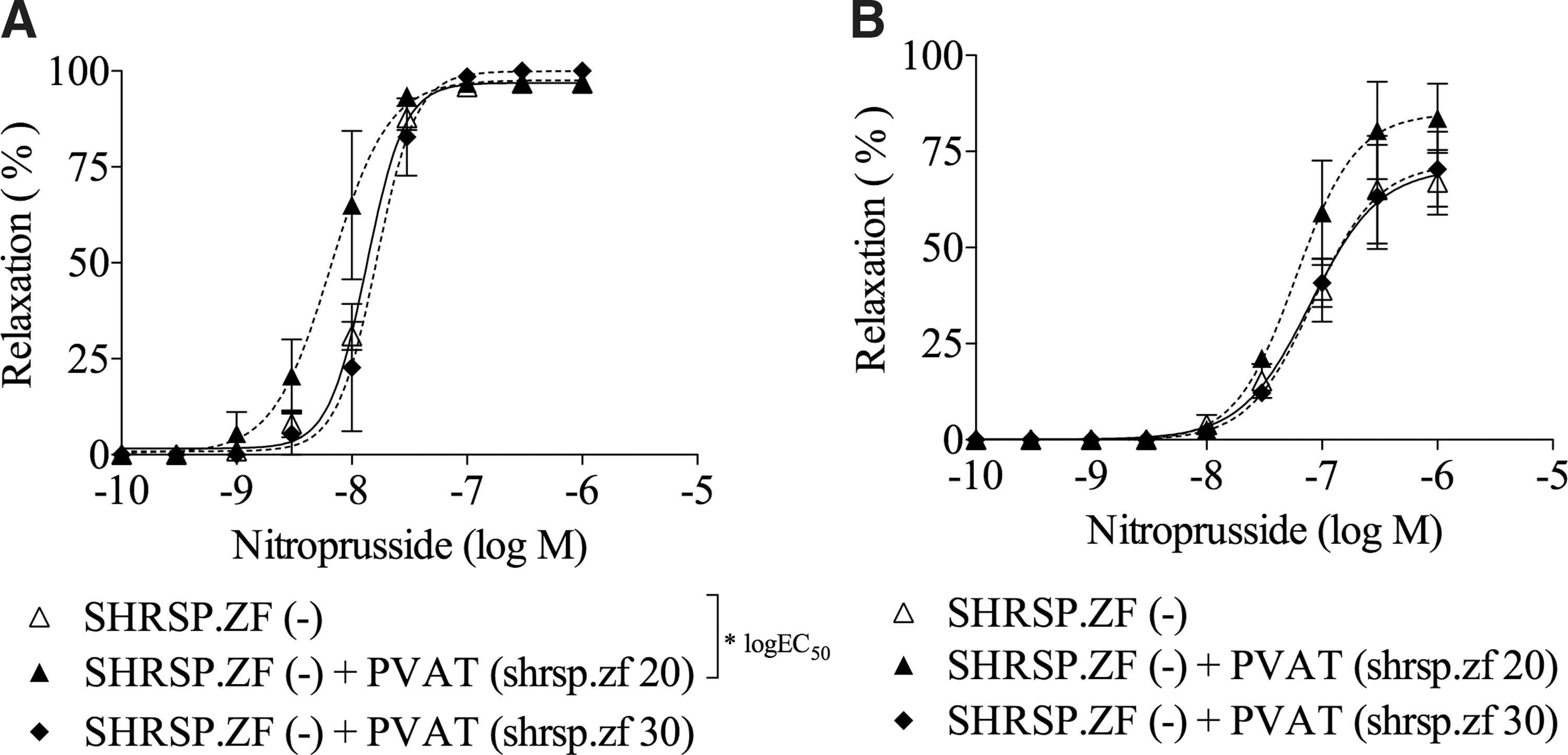
Effects of in vitro transplant of PVAT on vasodilations in response to nitroprusside in mesenteric arteries from SHRSP.ZF at 20
Independently of the vascular endothelium, exogenous NO donors can increase cGMP accumulation in vascular smooth muscle cells. We, therefore, compared cGMP accumulation elicited by nitroprusside (0.01 μM, 3 min) in SHRSP.ZF mesenteric arteries having PVAT with the arteries not having PVAT. At 20 weeks, cGMP accumulation in the arteries enveloped by PVAT was ∼2.6-fold higher than that in the arteries without PVAT (Fig. 5A, n = 5). In contrast, the incremental increase by PVAT at 30 weeks was less than that at 20 weeks (Fig. 5A, n = 5). The changes of cGMP levels were occurred in parallel with the relaxations at 3 min after treatment with nitroprusside (Fig. 5B, n = 5 each age).

Cyclic guanosine monophosphate accumulation
Discussion
This study demonstrates that PVAT augments vasorelaxations via activating endothelial NO synthase to compensate for vascular dysfunction present in arteries during continual exposure in vivo to metabolic abnormalities. Long-term exposure to metabolic abnormalities causes a breakdown of this compensatory PVAT-mediated mechanism, leading to further progression of cardiovascular diseases in MetS. This is the first article indicating that the duration of exposure to metabolic abnormalities affects vasomotor function of PVAT in MetS.
Structural and functional regional heterogeneity among PVAT is associated with specific vascular complications in patients. 19 Pairing of the arteries of SHRSP.ZF and PVAT of SHRSP.ZF was essential to transferring the PVAT enhancing effect, and interestingly, the enhancing effects of PVAT correlated to the degree of reduced vasodilation in SHRSP.ZF. From these findings, we proposed that PVAT has supplementary effects under conditions of impaired vasodilation in arteries. Our interpretation is supported by a study that provided evidence of a vasoprotective role for PVAT in arteries of animal models of hypertension, Dahl salt-sensitive rats, 20 and angiotensin II-induced hypertension. 21 Furthermore, the PVAT-enhancing effects were expressed when the adipose isolated from SHRSP.ZF was placed in close proximity to the PVAT-cleaned arterial ring of SHRSP.ZF. This observation means that transferable factor(s) is/are involved in the enhancing effects of PVAT in SHRSP.ZF artery. Using bioassay experiments involving the transfer of donor bath solution aliquots to acceptor preparations, PVAT was shown to exert an anti-contractile effect through the release of a transferable relaxing factor in aortas of Sprague-Dawley rats 22 and Wistar rats. 23 Furthermore, since the enhancing effects of PVAT were inhibited by removing the endothelium and treating with LNAME, the PVAT-derived factor(s) activates endothelial NO synthase, leading to NO production. This idea was supported by evidence that the accumulation of cGMP elicited by nitroprusside was higher in the presence of PVAT than that in the absence of PVAT in SHRSP.ZF mesenteric artery. From these findings, we concluded that PVAT regulates endothelial NO production in mesenteric artery of SHRSP.ZF with MetS. However, it is possible that PVAT also has a direct effect on smooth muscle cell responsiveness to NO. Similar to our findings, PVAT buffered vasoconstriction by activating endothelial NO synthase in aortas of Wistar and Dahl salt-sensitive rats. 20,23,24 Further studies are needed to understand the trigger to produce the PVAT-derived factor(s).
The enhancing effects of PVAT on agonist-induced vasodilations were diminished in SHRSP.ZF at 30 weeks. An anti-contractile effect, that is, versus serotonin, of PVAT is also decreased with age (20, 25, and 35 weeks) in mesenteric arteries of New Zealand obese mice.
25
These findings suggest that a decrease in the compensatory system of PVAT occurs during long-term exposure to metabolic abnormalities. The deterioration of the PVAT's effects, that is, at a later stage of MetS, may lead to development of cardiovascular pathology, resulting from enhancement of vasoconstriction. In a recent clinical study of 63 patients, researchers reported that the volume of PVAT for the left anterior descending coronary artery was higher in the group having vasospastic angina (n = 45) compared with their control group (n = 18).
26
In addition, in a report on an ongoing study of human autopsy samples, the authors reported preliminary evidence of histological differences in the PVAT in vessels with acute versus chronic coronary arterial disease.
27
The results of this study may provide some insight into the time-dependent differences in PVAT. We found that the enhancing effects of PVAT from SHRSP.ZF could not be substituted by replacement with PVAT from 30-week age of SHRSP.ZF. We propose that a decrease in production/release of a transferable relaxing factor may explain these age-dependent changes in the effects of PVAT on vasomotor functions. In mice with diet-induced obesity, an increase in NO production in mesenteric PVAT occurred during the initial 8-week period of dietary treatment
9
and preserved vascular function. This adaptive function of PVAT deteriorated as a result of “endothelial NO synthase uncoupling” and “
Structural and regional heterogeneity among different PVAT varies according to the blood vessel. 19 In our study, we only tested the PVAT surrounding the superior mesenteric artery of male animals with MetS to investigate the effects of PVAT on vascular functions. In rodents, this artery is surrounded by white adipose tissue whereas the thoracic aorta is surrounded by brown adipose tissue. 30 Interestingly, human adult epicardial fat exhibits features referred to as beige, 31 which has characteristics of both white and brown adipose tissues. Moreover, a recent study showed that there were sex- and blood vessel-related differences in PVAT adipocyte morphology. 32 To our knowledge, sex-related differences in the SHRSP.ZF model have not been studied. Further investigations of the interactions between age-related changes in PVAT and blood vessel- and/or sex-related differences in vascular functions are warranted for developing a full understanding of the pathophysiological significance of our findings. Although the focus of our study is an animal model of MetS, our data are also consistent with accumulating evidence from clinical studies that report that increased PVAT volumes, as measured by using echocardiography, computed tomography, magnetic resonance imaging, and ultrasound scan, are associated with cardiovascular diseases 33,34 and modulate the progression of atherosclerosis 35 and calcification 36 in humans. Dysfunction of PVAT causing vascular disease in humans requires further study. 33,37
In conclusion, PVAT can exert a regulatory influence on vasodilation via an NO-dependent pathway in MetS. With aging, and longer times of in vivo exposure to MetS disorder, the deterioration of PVAT enhancement of vasodilation may contribute to chronic vascular dysfunction in cardiovascular diseases and thus, it warrants further study as a therapeutic target.
Footnotes
Acknowledgment
This work was partly supported by MEXT KAKENHI (grant number 16K08563 to S.K.).
Author Disclosure Statement
No conflicting financial interests exist.