
Abstract
Energy homeostasis is coordinated by bidirectional communication pathways between the brain and peripheral organs, including adipose tissue, muscle, the pancreas, liver, and gut. Disruption of the integrated chemical, hormonal, and neuronal signals that constitute the gut–brain axis significantly contributes to disorders of metabolism and body weight. Initial studies of glucagon-like peptide-1 (GLP-1), a gut hormone released in response to the ingestion of nutrients, focused on its incretin actions to improve postprandial glucose homeostasis by enhancing meal-induced insulin secretion. However, GLP-1 is also a key player in the gut–brain regulatory axis with multiple effects on appetite and energy metabolism outside of its peripheral glucoregulatory actions. In this review, we explore the function of GLP-1 as a component of the gut–brain axis in the regulation of energy homeostasis, and consider the implications of this role for the development of therapeutic treatment options for obesity.
Introduction
Obesity, a complex multifactorial disease, has emerged as the leading global health care issue. The etiological complexity and its association with numerous comorbidities create unique management challenges when treating the patient with obesity. As a result, a clear knowledge of the neurohormonal regulation of appetite and metabolism is vital for an understanding of the mechanisms behind current and future medications for weight management.
Dysregulation of the homeostatic systems governing energy metabolism leads to unwanted weight gain and defense of a new, higher body weight in patients with overweight or obesity. 1 –3 Fundamentally, any weight-loss intervention must act either directly or indirectly to induce a state of negative energy balance. 4 However, for this weight loss to then be sustained, successful treatment must interfere with the homeostatic metabolic, endocrine, and behavioral mechanisms intent on returning an individual with overweight or obesity to their former weight. 5
Energy homeostasis is coordinated by the brain and involves the integration of chemical, hormonal, and neuronal signals from peripheral organs such as adipose tissue, pancreas, liver, and the gut, constituting the gut–brain axis. 6,7 Peripheral signals, such as leptin and insulin, reflect the body's level of energy stores and various gut factors convey the types and availability of nutrients signifying the timing, frequency, caloric amount, and macronutrient content of ingested meals. Disruption of the signals constituting this axis is thought to be a common pathophysiological pathway that leads not only to unwanted weight gain and the development of obesity, but is now also recognized to be a key mediator in the development of metabolic disorders such as dyslipidemia, prediabetes, and type 2 diabetes. 8
A key player in the gut–brain axis is glucagon-like peptide-1 (GLP-1), a hormone released in response to meal ingestion that was originally described for its incretin properties because of its capacity to enhance meal-related insulin secretion, suppress glucagon, and thus improve postprandial glucose homeostasis. However, outside of their glucoregulatory role, GLP-1 receptor agonists (GLP-1RAs) have also demonstrated effects on appetite and energy metabolism, likely by acting directly on specific areas of the brainstem and hypothalamus. 7
In this review, we explore the role of GLP-1 as a component of the gut–brain axis involved in the regulation of energy homeostasis, and the implications of this role for the development of therapeutic treatment options for obesity.
The Gut–Brain Axis and GLP-1
The gut–brain axis is a bidirectional communication pathway linking the gut with the key centers of the peripheral nervous system (PNS) and central nervous system (CNS), contributing to regulation of energy homeostasis. 9 More than a century ago, Pavlov reported that the sight, smell or taste of food without swallowing stimulates gastric secretions in dogs. 10 Around the same time, Starling and Bayliss demonstrated that the intravenous infusion of scraped mucosa from the duodenum immediately stimulated pancreatic secretions. 11 They postulated that this phenomenon was the result of factors present within the gut lining, referred to as secretin. 11 It is now well recognized that the presence of nutrients detected in the mouth, stomach, and small intestine, often mediated through activation of G protein-coupled receptors (GPCRs), 12 affects the secretion of numerous gut hormones. Among these known appetite-altering gut peptides, ghrelin is recognized as an orexigenic hormone due to its ability in increasing hunger and food intake, and peptides such as cholecystokinin, peptide YY, oxyntomodulin, and GLP-1 are recognized as satiety hormones, with anorexigenic (appetite suppressing) effects. 13
GLP-1 is a 30 amino-acid peptide [GLP-1-(7-36)] derived from the preproglucagon (GCG) gene, 14 which is expressed in the pancreatic α-cells and intestinal L-cells 15 as well as in neurons located in the brainstem and the hypothalamus. 16 Tissue-specific posttranslational processing of proglucagon peptide, as a result of differential activities of prohormone convertase 1/3 and 2 (PC1/3 and PC2), 17 yields peptides specific to the pancreas, intestine, and brain. 14,15 Glucagon is the main proglucagon-derived peptide in the pancreatic α-cell given much higher expression of PC2 compared with PC1/3 in this tissue, whereas PCI 1/3 in the intestinal L-cells and neurons of the nucleus of the solitary tract (NTS) of the medulla generate GLP-1. 14
The majority of circulatory GLP-1 is secreted dose-dependently in response to nutrient ingestion by neuroendocrine L-cells located throughout the gastrointestinal (GI) tract (predominantly in the distal ileum and colon). 14 Observation of a colocalization of PC1/3 and GCG expression in the caudal region of the NTS within the brain has also indicated that that GLP-1 is produced in this region. 18,19 The stimuli responsible for the synthesis and secretion of this brain-derived GLP-1 remain largely unknown.
GLP-1 activity is mediated by the GLP-1R, a member of the B1 family of seven-transmembrane GPCRs. 20 GLP-1Rs are expressed in pancreatic β-cells as well as various other tissues, including neurons in specific central brain regions, the peripheral afferent parasympathetic nervous system, atrial cardiac myocytes, pulmonary endothelial cells, kidney, and the GI tract, 14 reflecting the potential for pleiotropic physiological effects of GLP-1 beyond its insulin secretory effect.
The assumption that physiologic actions of endogenous GLP-1 are entirely mediated by binding to organ-specific GLP-1Rs has been challenged. A significant amount of GLP-1 secreted from the gut is rapidly degraded by the enzyme dipeptidyl peptidase-4 (DPP-4) to an inactive component, GLP-1-(9-36), before reaching the portal vein. 21 Thus, given its extremely short plasma half-life of 1–2 min, only a small portion of secreted GLP-1 is thought to reach target organs. Therefore, in addition to binding to GLP-1R at the target organs, GLP-1 effect is also mediated through activation of afferent neural pathways initiating in the gut or hepatic portal veins. 22,23
Glycemic-reducing effects of GLP-1 in patients with type 2 diabetes led to development of a new class of drugs utilizing GLP-1 signaling to treat diabetes. Using strategies to resist the degrading effects of DPP-4 on GLP-1 has produced two categories of GLP-1 based drugs: DPP-4 inhibitors, resulting in higher concentrations of native GLP-1; and GLP-1RAs, modified so as to have longer half-lives than native GLP-1. In general, compared to DPP-4 inhibitors, GLP-1RAs are more effective in reducing HbA1c (0.4%–1.6% vs. 0.4%–0.8%; Table 1). Findings from the clinical trials have also revealed that administration of GLP-1RAs induces weight loss in addition to glucose improvement (Table 1). While the glycemic-reducing effects of GLP-1RAs are attributed mainly to endocrine actions of these compounds at the level of pancreatic islets, their effect on energy homeostasis likely involves central GLP-1 signaling.
The Effect of Available Dipeptidyl Peptidase-4 Inhibitors and Glucagon-Like Peptide-1 Receptor Agonists on Glycemic Control and Weight Loss
Data are reported as mean. The source of the data is from the product label unless otherwise indicated. Percentage change in body weight is calculated for products reporting a baseline weight measurement in the product label. Product data are for monotherapy.
Data are from the largest clinical trial with the given product.
Approved dose for weight loss management.
DPP-4, dipeptidyl peptidase-4; GLP-1RA, glucagon-like peptide-1 receptor agonist; n/a, data not available.
Physiological Effects of GLP-1 Focusing on Energy Balance
Energy homeostasis is governed by a complex interplay between nutrients, neural signaling, adipokines, and gut peptides, whose actions are mediated by binding directly to neuronal receptors in the CNS or PNS. Both enteric and autonomic nervous systems are involved in carrying afferent signals initiated by nutrient or GI hormones from the gut to the CNS via vagal or spinal pathways, as well as efferent signals from the CNS back to the gut. 9
Central nervous system
Rodent studies
One of the key brain regions involved in energy intake and balance is the arcuate nucleus (ARC) of hypothalamus (Fig. 1). 24 The ARC is situated near the median eminence of the hypothalamus, which is thought to lie partially outside of the blood–brain barrier in humans. 25 This positioning is potentially advantageous in that it allows ready access for peripheral hormones that regulate food intake without having to first penetrate the blood–brain barrier. Similarly, the area postrema, another important brain area involved in energy regulation and juxtaposed to the NTS in the brainstem, 7 also possesses an incomplete blood–brain barrier. 26 Hence, meal-related circulating gut hormones can readily cross from the bloodstream and bind directly to their receptors in this region of the brainstem.
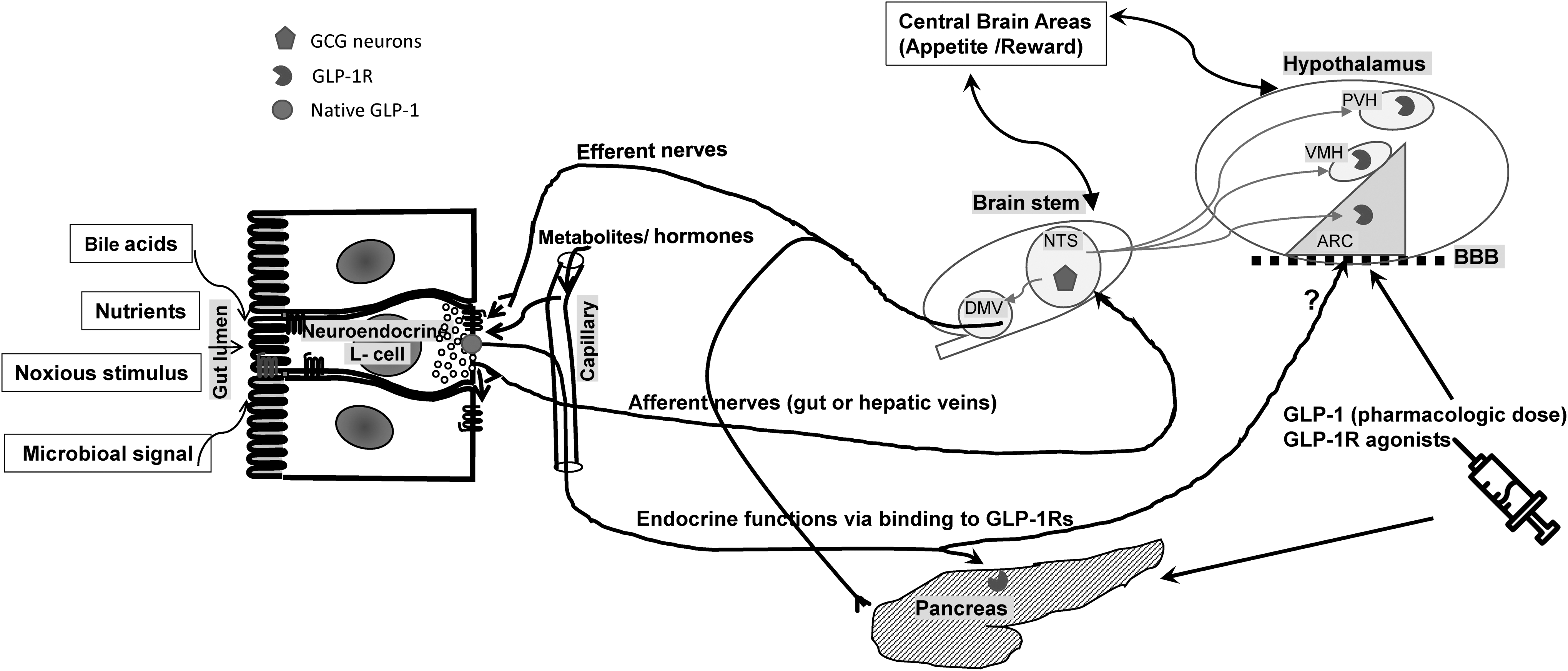
The role of GLP-1 in energy balance within the complex gut–brain interplay. Gut-derived GLP-1 is secreted from intestinal L-cells into circulation as a result of nutrient ingestion as well as other stimuli, such as bile acids, microbiota, and noxious stimuli. Physiologic effects of endogenous gut-derived GLP-1 is mediated by its binding to GLP-1Rs (pancreatic islet cells) or stimulating vagal nervous system, transmitting signals to the CNS (afferent nerves) and from the CNS back to the peripheral organs (efferent nerves). GLP-1Rs are found within some brain regions, particularly the ARC, PVH, and the VMH. Endogenous GLP-1 produced by GCG neurons of NTS of the medulla affects food intake by its binding to the hypothalamic GLP-1Rs. The GCG neurons receive information from peripheral organ through afferent nerves as well as areas in the brain and project back to the hypothalamus as well as the vagal efferent neurons in the DMV and the intermediolateral cell column of spinal cord. Anorexic effects of GLP-1 (above physiologic concentrations) or GLP-1R agonists are likely mediated through binding directly to hypothalamic GLP-1Rs after crossing the blood–brain barrier (BBB). ARC, arcuate nucleus; CNS, central nervous system; DMV, dorsal motor vagal nucleus; GCG, preproglucagon; GLP-1R, glucagon-like peptide-1 receptor; NTS, nucleus of the solitary tract; PVH, paraventricular nuclei of the hypothalamus; VMH, ventromedial nuclei of the hypothalamus.
GLP-1Rs are found within many of these brain regions, including the arcuate, paraventricular, and the ventromedial nuclei of the hypothalamus. 27 –30 In addition to direct receptor binding and neuronal activation at these sites, peripherally secreted GLP-1 also targets GPCRs on the vagus nerve, which transmit signals to the NTS via vagal afferents. 31 Centrally, GLP-1 is secreted by the GCG neurons of NTS and directly activates GLP-1Rs found throughout the brain. 32 Absence of GLP-1R on GCG neurons suggests that the function of these neurons is not solely regulated by GLP-1 itself. 33 From their location in the hindbrain, GCG neurons receive information via afferent neurons from the periphery as well as the CNS (including the hypothalamus), and, in turn, project reciprocally up to the hypothalamus and amygdala as well as to vagal efferent neurons in the dorsal motor nucleus of the vagus and to the intermediolateral cell column of spinal cord. 34 Both nutrient and noxious stimuli, 35,36 as well as gastric distension, 37 have been shown to activate these neurons.
The role of GCG neuronal regulation in energy homeostasis circuitry remains largely unclear. Reduction in GCG expression in the NTS using viral knockdown of GCG expression 38 has been shown to induce weight gain and increase adiposity, mirroring the effects of chronic blockade of CNS GLP-1R using intracerebroventricular infusion of exendin-9-39. 39 In a recent study, selective activation of GCG neurons using designer receptors exclusively activated by designer drugs acutely reduced food intake in rats on a standard or high-fat diet. 40 However, chronic activation of GCG neurons using this method led to weight loss only in mice on high-fat diet with no effect on animals on standard diet. 40 Activation of GCG neurons in this study had no influence on the known GLP-1 actions in regulating glucose metabolism or stress response (malaise, hypothalamic-pituitary-adrenal axis activation, or sympathetic nerve activation). 40
Although both peripherally and centrally administered GLP-1 affects appetite, the differential effects of endogenous GLP-1 secreted from the gut versus CNS in regulation of energy intake are poorly understood. Given the rapid degradation of GLP-1 by DPP-4, peripheral vagal activation rather than direct CNS effect likely mediates the feeding effect of postprandial gut-derived GLP. 34 Moreover, the effects of endogenous gut- and brain-derived GLP-1 on energy balance seem to be independent of each other since CNS administration of physiologic concentrations of GLP-1 in rodent models has been shown to lower calorie consumption and induce weight loss, whereas peripheral GLP-1 infusion has no effect on energy food intake. 41
In contrast to endogenous GLP-1 at physiologic concentrations, peripherally administered GLP-1RAs that avoid DPP-4 degradation have been shown to lower food intake mainly through direct action on CNS GLP-1R, as concurrent administration of a central GLP-1R antagonist eliminates the anorectic effect of the peripheral GLP-1RA. 28 Using a labeled GLP-1RA (liraglutide), it was also demonstrated that this compound binds exclusively to neurons within the ARC and other areas in the hypothalamus rather than to peripheral locations such as the vagus nerve. 30 Furthermore, selective deletion of neuronal GLP-1Rs eliminates both the uptake of liraglutide, a GLP-1RA, into the brain 30 and its ability to affect appetite and weight loss, 42 indicative of its central effect.
Human studies
The effect of exogenous infusion of GLP-1-(7-36) on energy consumption in humans is consistent with those in animal model studies, in which ad libitum food intake in both lean and obese individuals is decreased dose-dependently 43 when circulatory GLP-1 is increased to levels above postprandial physiological concentrations. 44 In an early GLP-1RA study in humans, 5 weeks of once-daily treatment with liraglutide 1.8 or 3.0 mg in 49 nondiabetic individuals with obesity resulted in ∼15% lower energy intake accompanied by a ∼2 kg weight loss. 45
Over the last decade, functional magnetic resonance imaging (fMRI) has been used to measure neural activity in various brain areas for the study of mediators of food responses in humans. In lean healthy subjects, blocking GLP-1R alters neural activities in central brain regions responsive to oral food stimuli. 46 In individuals with obesity, with or without type 2 diabetes, appetite- and reward-related areas of the brain that have shown increased brain response to food pictures compared to lean controls 47,48 are reduced during infusion of GLP-1RA. 47 The specificity of these effects was demonstrated by prevention of these neural responses to visual and oral food cues when a GLP-1RA was administered during coinfusion of a GLP-1R blocker. 47,48
Weight loss accompanying GLP-1 infusion in humans has also been attributed to drug-induced malaise or food aversion. However, clinical trials using GLP-1RAs have shown that nausea, if experienced at all, is generally short-lived and typically resolves within the first few weeks of treatment, yet, the weight loss from these compounds is durable, and the frequency and duration of nausea is not related to the amount of weight loss achieved. 49 Furthermore, in SCALE Obesity and Prediabetes, a large clinical trial (n = 3731) in which liraglutide 3.0 mg was administered daily, there were no significant differences in weight loss between individuals experiencing at least one episode of nausea or vomiting (n = 301) and those reporting no episodes of nausea or vomiting during the 56 weeks on treatment. 50 These findings indicate that the weight loss observed during GLP-1RA treatment cannot be fully accounted for by the occurrence of GI adverse events.
Altogether, studies in animal models and humans confirm an important role for central GLP-1 signaling in the regulation of appetite control and energy balance.
Gut
The bidirectional communication of the gut–brain axis involves the modulation of gut functions that may affect energy intake and expenditure through changes in gut motility or gut microbiota. 9
Gut motility
While endogenous GLP-1 has been shown to have trivial effects on gastric emptying (GE), 51 –53 pharmacological doses of GLP-1 or GLP-1RAs have shown notable reductions in the rate of food emptying from the stomach into the gut. 54,55 The mechanisms underpinning this reduction or delay in GE are complex. It is likely that the effects on gastric motility are mediated by GLP-1-related activation of the vagal nerve as well as direct actions 56 through GLP-1Rs found along the GI tract, such as enteric neurons of the small and large intestine. 14,57
Interestingly, short-acting GLP-1RAs appear to reduce GE to a greater extent than long-acting GLP-1RAs. 58 Although underlying mechanisms for differential effects on GE are yet to be fully elucidated, it is plausible that GE effect is subject to tachyphylaxis/desensitization during steady, elevated concentrations of long-acting GLP-1RAs compared with peaks and troughs of drug concentrations of short-acting GLP-RAs. 58
Whether the delay in GE comprises part of the weight-loss mechanism of action of GLP-1RAs is debated. Using scintigraphy 54 (the gold standard technique for measuring GE), liraglutide has been shown to be associated with significantly delayed GE of solids after 5 and 16 weeks of treatment compared with placebo. GE at week 5 was significantly associated with the degree of weight loss at 16 weeks. However, despite evidence of tachyphylaxis to drug-induced deceleration of GE after 16 weeks, weight loss effects of treatment remained unchanged. Therefore, the early changes in GE may also contribute to liraglutide-induced weight loss.
Notably, delayed GE does not seem to be linked to one of the most common side effects of GLP-1RAs, nausea. 59 GE measured using scintigraphy did not differ among those individuals taking liraglutide who reported nausea and those who did not. 54 Similarly, in a study of 16 individuals treated with exenatide, no significant differences in the rates of GE, determined using scintigraphy, were observed between individuals who experienced GI adverse events (e.g., nausea) and those who did not. 60
Intestinal flora
Long thought to be inactive commensal organisms, microbiota has been implicated in alteration in glucose metabolism and energy homeostasis within their host based on studies in animal. Enteric microbiota are observed throughout the GI tract, and although the profile differs between individuals, the relative abundance of differing species is similar in healthy subjects. 9 Given the large diversity displayed in gut flora, defining what is “normal composition” for human gut microbiota is difficult. 61 However, individuals with obesity display less microbial diversity than those of normal weight. 62
The differences in bacterial diversity between individuals who are lean and those with obesity are thought to result in alterations to microbiota-generated metabolites that, in turn, affect energy homeostasis through the regulation of peptide secretion, including GLP-1. 62,63 Mice bred to exhibit an absence of gut flora demonstrate a reduced expression of a number of GPCRs thought to mediate gut peptide release from enteroendocrine cells, which is thought to promote increased energy intake in these animals. 64 Furthermore, in mice, the imbalance of gut microbiota that occurs in obesity reduces the expression of GLP-1Rs. 65 In humans, evidence suggests that prebiotic treatment increases microbiome diversity, which in turn may modulate levels of GLP-1 66 and promote the feeling of satiety. 67 Given the close anatomical proximity, gut microbiota could potentially alter the nutrient-sensing capacity of the enteroendocrine cells and subsequent gut peptide release. 68 However, 7-day treatment with antibiotics had no effect on glucose tolerance despite increasing bacterial diversity in nondiabetic obese individuals diagnosed with either impaired fasting glucose or impaired glucose tolerance. 69
Interestingly, and with implications for the development of future obesity treatments, the gut microbiota profile in rats appears to change following treatment with liraglutide and after bariatric surgery to reflect the profile of lean subjects. 70,71 It has been suggested that the GLP-1RAs that influence gut transit time and the rate of GE could modify microbiota diversity due to alterations nutrient composition. 71 Taking these data into consideration, while it is possible that microbiota may play a role in the gut–brain axis promoting improved energy balance, well-designed studies in human are lacking.
GLP-1-Based Therapeutic Intervention for Treatment of Obesity
Although GLP-1RAs were originally developed to treat diabetes, the additional role of GLP-1 in energy balance broadens the spectrum of uses for its beneficial effects, including for weight management in people with obesity. This is evidenced by the fact that all GLP-1RAs approved to treat diabetes have been shown to result in some degree of weight loss (Table 1), with one currently approved for chronic weight management in both the United States and Europe. 72
The weight-loss efficacy of these compounds differs, likely due to variations in chemical modifications that alter their pharmacokinetic parameters or pharmacodynamic properties to increase their half-lives compared with native GLP-1. 73 For example, the relatively large molecule albiglutide 74 may activate GLP-1R in the hypothalamus to a lesser extent than smaller molecules such as liraglutide and exenatide. This may be the result of a hindered passage through fenestrated capillaries as the compounds enter the brain through circumventricular organs, 75 leading to a smaller weight loss response when compared with smaller molecules that are able to activate central GLP-1R to a greater extent.
Weight-loss procedures such as Roux-en-Y gastric bypass (RYGB) and sleeve gastrectomy (SG) have gained popularity in recent years, given their effectiveness in the treatment of obesity and its comorbidities. 76 –78 Patients following RYGB, and to a lesser extent following SG, experience changes to a number of appetite hormones, including an 10-fold increase in meal-induced GLP-1 secretion, 79 –81 likely due to rapid nutrient emptying from the stomach pouch to the distal small intestine. Given the known anorectic effect of GLP-1, the secretion of this peptide in response to nutrient ingestion was one of the first gut-hormones proposed to explain the robust and sustainable weight loss following RYGB.
However, the benefit of these procedures on weight regulation is unlikely to be explained by alteration in just one factor, as rerouted GI surgeries leads to a myriad of alterations in gut function, meal-induced hormone secretion, and changes in microbiome. For example, GLP-1R knockout mice do not appear to show any difference in weight loss after gastric bypass or SG compared to wild animals. 82 –84 On the contrary, a recent study in humans using fMRI to measure CNS activation in response to food consumption in women with obesity showed significant reductions in activity of insular cortex (key structure controlling emotion involved with the pleasure experienced from palatable food) from presurgery to 4 weeks after RYGB, corresponding with large increases in meal-stimulated GLP-1 levels. 85 Importantly, they demonstrated a reversal of these changes by coadministration of a GLP-1R antagonist, hinting toward a beneficial contribution of this peptide in energy regulation after RYGB. 85 While these data are only suggestive of the role GLP-1 in mediating weight loss after RYGB, the discrepancy between animal model and human responses points out the necessity of well-designed studies in humans to address this question. Interestingly, the broad changes in different gut-hormonal profiles following bariatric surgeries that favor sustained weight loss 81,86,87 have encouraged strategies to replicate these effects through production of hybrid peptides and coformulations. Examples of these hybrid peptides are GLP-1/glucagon 88 or GLP-1/GIP 89 coagonists, which have shown promise in their effectiveness to improve appetite and diabetes control.
Conclusion
Energy homeostasis is regulated by a complex interaction of multiple organ systems, including the gut–brain axis. Disruption of this complex system is associated with the development of obesity and diabetes. GLP-1, a 30-amino acid peptide predominantly secreted by neuroendocrine L-cells in the gut, is thought to be a key regulator of this homeostatic axis. Although the effect of gut-derived GLP-1 at physiologic concentrations on energy balance seems to be mediated by action on peripheral vagal afferents, recently developed GLP-1RAs appear to have significant weight-loss effects predominantly through direct central activation of GLP-1Rs.
The role that GLP-1 plays in the gut–brain axis offers rich clinical potential not only for improved glucose metabolism but also to improve appetite control and adiposity. Weight-loss efficacy differs among GLP-1RAs, in part, due to differing pharmacokinetic and pharmacodynamic properties. Although GLP-1RAs are mainly approved for the management of type 2 diabetes, liraglutide (3 mg) is recently approved for weight management in patients with obesity, and several are in early clinical trials.
Footnotes
Acknowledgments
The authors are grateful to Jamie Cozens, MSc, of Watermeadow Medical, UK (an Ashfield company), for writing assistance in the development of this article. This assistance was funded by Novo Nordisk, which also had a role in the review of the article for scientific accuracy.
Authors' Contributions
Dr. M.S. and J.Q.P. planned out the first draft in detail, both have commented extensively on subsequent drafts, and both have edited the final draft for submission.
Author Disclosure Statement
M.S. receives grant funding from NIH (DK083554 and DK105379); J.Q.P. receives compensation as an advisor for Novo Nordisk and receives grant funding from the NIH.