
Abstract
Background:
Exposure to intermittent hypoxia (IH) may play a role in the development of metabolic impairments in the context of obstructive sleep apnea syndrome, probably by elevated plasma levels of free fatty acids. Employing gas-permeable cultureware to grow differentiated human and mouse adipocytes in vitro, we directly studied the effects of pericellular oxygen fluctuations on key adipocyte metabolic functions—spontaneous lipolytic rates, triglyceride accumulation, de novo lipogenesis, and expression of adipocyte-specific marker genes.
Materials and Methods:
3T3-L1 fibroblasts and human subcutaneous preadipocytes were differentiated under conditions that induced repetitive pericellular-oxygen cycles IH between 1% O2 (5 min) and 16% O2 (5 min), continuously for 14 days or under control conditions. Chemicals were used to inhibit the flux of acetyl-CoA from glycolysis (alfa-cyano-4-hydroxy cinnamate) or the tricarboxylic acid cycle (SB204990), or to stimulate the flux of acetyl-CoA from pyruvate to the lipogenic pool. Lipolytic rate, intracellular lipids, and expression of adipocyte differentiation markers were assessed and t-test or ANOVA were used to find significant differences.
Results:
The rate of lipolysis increased by 211% in 3T3-L1 cells and by 39% in obese human adipocytes. Exposure to IH reduced intracellular lipid stores by 37% and reduced the expression of adipocyte differentiation markers. Pharmacological stimulation or inhibition of de novo lipogenesis did not modify the intracellular lipid content under IH.
Conclusions:
Pericellular oxygen fluctuations directly stimulated lipolysis, but did not increase de novo lipogenesis from endogenous substrates. Similarly, IH hampered adipocyte differentiation from precursors.
Introduction
Obstructive sleep apnea (OSA) syndrome affects 5%–15% of adults 1 and is characterized by sleep-associated repetitive narrowing or occlusion of the upper airway resulting in periodic reductions in blood and tissue oxygenation (hypoxemia) as well as sleep fragmentation. OSA has been associated with hypertension, atherosclerosis, and all-cause mortality. 2,3 Furthermore, OSA was linked to glucose intolerance, insulin resistance, and development of type 2 diabetes mellitus (T2DM), irrespective of traditional risk factors. 4
Although epidemiological evidence linking OSA with metabolic impairments is convincing, mechanisms mediating such associations remain only partially understood. Human and animal studies showed that insulin resistance, hyperglycemia, glucose intolerance, and impaired insulin secretion are consequences of exposure to intermittent hypoxia (IH), 5 –8 which are likely mediated by multiple mechanisms, including oxidative stress, accelerated sympathetic tone, increased plasma cortisol levels, endocrine changes, and induction of a proinflammatory state. 4 Furthermore, adipose tissue functions were reported to be modified by sustained hypoxia in obese subjects 9,10 or in chronic obstructive-pulmonary disease. 4,11 In particular, elevated levels of plasma free fatty acids (FFAs), released during lipolysis, promote insulin resistance in muscles and the liver, and inhibit pancreatic insulin secretion, thereby causally contributing to the development of T2DM. 12,13 Indeed, inhibiting lipolysis improved some of the glucose-homeostasis impairments induced by IH in a mouse model of OSA syndrome. 14
Sustained hypoxia modified gene expression and secretion of adipokines, and impacted the viability, growth, and metabolism of adipose tissue in vitro. 15,16 Importantly, significant differences in cellular responses to sustained versus IH were observed, including differences in transcriptional regulation, receptor expression, intracellular signaling, and metabolic effects. 17,18 Previously, we demonstrated that sustained mild hypoxia (4% O2) increased adipose tissue lipolysis and adipogenesis, and promoted triglyceride storage and differentiation of adipocytes in vitro 19 ; however, the impact of IH (OSA model) remains unclear.
Unfortunately, the physical laws of gas diffusion in liquids together with O2 consumption by adherent cells lead to variable and unpredictable pericellular O2 concentrations differing from that in the incubator atmosphere, substantially hampering prolonged exposures to sustained or IH. 20 –23 As exposure to IH in the context of OSA lasts for several hours per day, prolonged exposures to predictable pericellular O2 levels are warranted to investigate cellular responses and to identify potential pharmacological targets.
The aim of this study was to investigate the effects of prolonged IH exposure on key adipocyte functions, that is, spontaneous-lipolytic rate, intracellular triglyceride stores, de novo lipogenesis, and adipocyte differentiation. In addition, pathways regulating de novo lipogenesis were explored using pharmacological compounds.
Materials and Methods
3T3-L1 and human preadipocyte cell culture, differentiation, and hypoxic exposure
3T3-L1 fibroblasts (for subsequent use in experiments investigating intracellular lipid stores, spontaneous lipolytic rate, expression of adipogenesis markers, and the contribution of glycolysis/tricarboxylic acid cycle to the acetyl-CoA pool) and human subcutaneous preadipocytes derived from obese or nonobese donors for spontaneous lipolysis investigation (SP-F-1 and SP-F-3, Zen-Bio, Inc.) were cultured according to manufacturer`s instructions in a CO2 incubator at 37°C and 5% CO2 on fluorocarbon-bottom culture plates (Lumox; Sarstedt, Nümbrecht, Germany). At passage number 11 (3T3-L1 cells) or 3 (human cells), the cells were expanded 4–5 days in T75 flasks, harvested, and plated (5000 cells/cm2) in fluorocarbon-bottom plates.
After reaching confluence, the dishes were placed in system enabling exposure to IH, as previously described. 24 In brief, the system induced exposure to IH by delivering 5-min cycles of exposure to a gas mixture containing 16% O2 and 5% CO2, followed by a 5-min exposure to 1% O2 and 5% CO2 (Linde Gas a.s., Prague, Czech Republic). Control exposures were performed using the identical system where both gas mixtures contained 16% O2 and 5% CO2. Cells were fed every other day. Control as well as IH exposures lasted for 14 days as recommended by the supplier of the cells—after 14 days of incubation/differentiation, cells reach maximal differentiation (later, cells start to detach from the plastic cultureware bottom and die due to excessive lipid accumulation).
Pharmacological compounds were tested only in 3T3-L1 cells by adding individual drugs to culture media throughout the entire 14-day IH- or control-exposure period, as follows: (1) pyruvate dehydrogenase activator, 5 mM dichloroacetate (DCA, D54702; Sigma-Aldrich, St. Louis, Mo), (2) ATP-dependent citrate lyase (ACLY) inhibitor, 40 μM SB20499, that is, (3R,5S)-rel-5-[6-(2,4-dichlorophenyl) hexyl] tetrahydro-3-hydroxy-2-oxo-3-furan acetic acid (product no. 4962; Tocris Bioscience, Bristol, United Kingdom), and (3) inhibitor of pyruvate transport to mitochondria, 150 μM α-cyano-4-hydroxy cinnamate (CHCA, C2020; Sigma-Aldrich). Six independent replicates were performed for all experiments investigating the effect of pharmacological compounds on lipogenesis except for vehicle for CHCA treatment, where five independent replicates were performed.
Spontaneous lipolysis determination
Differentiated cells were starved for 24 hr in serum-free medium (Zen-Bio, Inc.), allowed to recover for 1 hr in Krebs Ringer Bicarbonate buffer (10 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid [HEPES], 2% fatty acid-free bovine serum albumin, and 6 mM glucose at pH 7.4), and subsequently incubated for 3 hr to assess the spontaneous-lipolysis rate. Glycerol released into the medium was measured colorimetrically (F6428; Sigma-Aldrich) and normalized to the total lipid content, as previously described. 25 Six independent replicates were performed for 14-day IH experiments (human obese/nonobese adipocytes and 3T3-L1 cells), whereas three independent replicates were performed for 24-hr IH exposures in 3T3-L1 cells.
Intracellular lipid quantification
Cells were fixed for 1 hr in 10% formalin, washed twice with 1 mL phosphate-buffered saline, and stained with a working solution of Oil Red O (O0625; Sigma Aldrich) for 1.5 hr. Subsequently, the cells were washed, air dried, and the Oil Red O stain was extracted with 0.5 mL isopropanol. Total lipid contents were determined quantitatively by measuring the absorbance of Oil Red o at 510 nm and comparing the signal to a standard curve constructed from triolein (Y0001113; Sigma Aldrich) dissolved in hexane (25 mg/mL), sorbed to a polystyrene well, dried, stained with Oil Red O and subsequently extracted with isopropanol. 26
Gene-expression analysis
Total RNA was extracted using TriPure reagent (11667157001; Sigma Aldrich), treated with DNase (Roche Diagnostics, Mannheim, Germany) and reverse transcribed using the SuperScript III system (Invitrogen, Carlsbad, CA). Gene expression levels of diacylglycerol O-acyltransferase 2 (DGAT), fatty acid binding protein 4 (FABP4), adipocyte fatty acid synthase (FASN), hormone sensitive lipase (LIPE), perilipin 1 (PLIN1), ACLY, and the reference genes TATA box-binding protein (TBP) and beta-glucuronidase (GUSB) were assessed by quantitative PCR (ABI 7500; Applied Biosystems, Carlsbad, CA) using TaqMan probes (product nos. Mm00499536_m1, Mm00445878_m1, Mm00662319_m1, Mm01197698_m1, Mm00495359_m1, Mm00558672_m1, Mm01302282_m1, Mm00446971_m1, and Mm01197698_m1, respectively). The geometric means for TBP and GUSB were used as endogenous controls. Subsequently, ΔC t values (C t gene of interest— C t housekeeping gene) were used for statistical analysis (two-tailed t-test). Results are presented as fold-change values where control conditions were considered as 100%.
Statistical analysis
The effects of IH exposure on outcome variables (compared with the control exposure) were analyzed using an unpaired t-test. The effects of pharmacological substances on outcome variables were analyzed using two-way analysis of variance (ANOVA) with Fisher's least significant difference post hoc tests, and interactions between pharmacological treatments and IH exposure were determined. Kolmogorov–Smirnov test was performed on all variables confirming normal distribution. GraphPad Prism 7 software (GraphPad Software, San Diego, CA) was used for analysis and graphing of data. Data are presented as the mean ± standard error of the mean. A value of P < 0.05 was considered to reflect a statistically significant difference in all tests. Complete data set will be available as a Supplementary Data with this article.
Results
Intracellular lipid stores and spontaneous lipolysis
Exposure to IH for 14 days reduced total triglyceride accumulation by 37% when compared with the control exposure (1.37 ± 0.14 vs. 0.86 ± 0.04 mg lipids/well; P < 0.05), as shown in Fig. 1A. Identical IH exposure increased spontaneous lipolysis by 211% (glycerol in medium from 3T3-L1 cells; 0.52 ± 0.34 vs. 1.65 ± 0.42 μmol/mg lipids after 180 min; P < 0.05); however, a shorter exposure (24 hr) did not affect lipolysis (glycerol in medium: 0.30 ± 0.02 vs. 0.25 ± 0.04 μmol/mg lipids after 180 min; P > 0.05), as shown in Fig. 1B. To replicate these observations, human preadipocytes derived from obese or nonobese donors were also exposed to IH. The results shown in Fig. 1C revealed that IH increased lipolysis by 39% in adipocytes from obese individuals, but not in those from nonobese individuals (glycerol in medium: 1.75 ± 0.18 vs. 2.44 ± 0.25 μmol/mg lipids after 180 min for obese individuals; 1.14 ± 0.09 vs. 1.07 ± 0.09 μmol/mg lipids after 180 min for nonobese individuals; P > 0.05 as determined by ANOVA).

The effects of IH on adipocyte metabolism and differentiation. Exposure to IH for 14 days (5-min cycles, 16% O2 to 1% O2) reduced intracellular lipid stores
Adipocyte-differentiation markers
The gene-expression levels of key adipocyte-differentiation markers following exposures are summarized in Fig. 1D. IH reduced differentiation/adipogenesis and FABP4, DGAT, LIPE, and perilipin expression to 0.45 ± 0.01, 0.38 ± 0.02, 0.41 ± 0.02, and 0.43 ± 0.02 versus the control conditions (P < 0.05 in each case). Expression of a key enzyme involved in de novo lipogenesis (FASN) was also reduced by IH to 0.30 ± 0.01 of control conditions (P < 0.05), whereas the expression of ACLY, which provides acetyl-CoA for de novo lipogenesis, was unaffected by IH (1.0 ± 0.03 vs. 1.06 ± 0.04; P > 0.05).
Mechanisms mediating the effects of hypoxia
To further elucidate the interplay between intracellular lipid synthesis and lipolysis, we used pharmacological inhibitors/activators of key metabolic pathways that generate acetyl-CoA, the key substrate for lipid synthesis (Fig. 2). When the flux of pyruvate (generated during glycolysis) to acetyl-CoA was inhibited by CHCA, the intracellular lipid stores decreased by 29% under control conditions (1.0 ± 0.08 vs. 0.71 ± 0.02; P < 0.05), but not under IH exposure (1.0 ± 0.02 vs. 0.89 ± 0.03; P > 0.05), as shown in Fig. 3A. Similarly, stimulation of pyruvate flux with DCA contributed to the intracellular acetyl-CoA pool by stimulating lipid stores under control conditions (1.0 ± 0.10 vs. 1.31 ± 0.08; P < 0.05), but not under IH exposure (1.0 ± 0.03 vs. 0.86 ± 0.03; P > 0.05), as shown in Fig. 3B. Inhibiting acetyl-CoA generation from citrate (Krebs cycle) with SB204990 did not modify the lipid stores under IH conditions (1.0 ± 0.03 vs. 1.10 ± 0.05; P > 0.05), but increased intracellular lipids by 28% under the control exposure (1.0 ± 0.12 vs. 1.28 ± 0.07; P < 0.05), as shown in Fig. 3C.

Metabolic pathways supporting de novo lipogenesis. Acetyl-CoA (a key metabolite used for fatty acid synthesis) can be derived from glycolysis (pyruvate) or the Krebs cycle (through the reverse-tricarboxylic acid cycle), utilizing glutamine as the source of carbon. ACO, aconitase; GDH, glutamine dehydrogenase; IDH2, isocitrate dehydrogenase 2; MPC, mitochondrial pyruvate carrier; pharmacological compounds used: CHCA, α-cyano-4-hydroxy cinnamate (inhibitor of the mitochondrial pyruvate carrier); DCA, dichloroacetate (activator of pyruvate dehydrogenase); SB204990, inhibitor of the ATP-dependent citrate lyase.
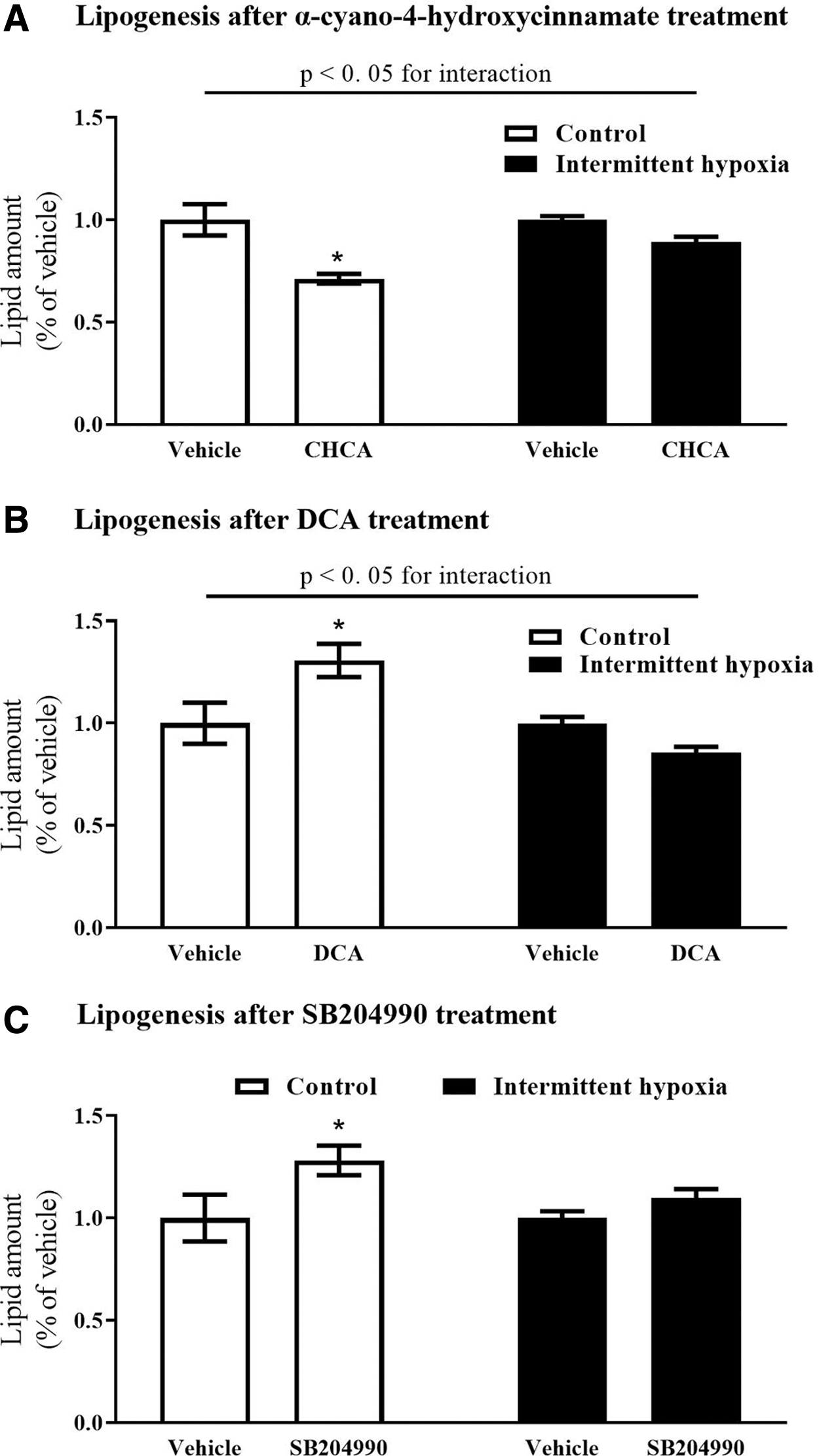
The impact of pharmacological substances on de novo lipid synthesis. Inhibition of pyruvate (an end product of glycolysis) contributed to the lipogenic pool by antagonizing pyruvate mitochondrial transporter with α-cyano-4-hydroxycinnamate inhibited lipogenesis under control conditions, but did not modify lipogenesis under IH exposure
Discussion
Employing cultureware with a gas-permeable well bottom, enabling rapid and reproducible changes in pericellular O2 concentrations, 24,27 this study demonstrated that adipocyte metabolism and differentiation were directly modified by exposure to IH. Mice and human adipocytes from obese individuals exposed to IH showed increased spontaneous-lipolysis rates, resulting in decreased intracellular lipid stores. Furthermore, the reduction of intracellular lipids was aggravated by reduced differentiation of adipocytes from immature precursors and by the lack of de novo lipogenesis stimulation under IH conditions.
Previous studies investigating adipocyte responses to sustained hypoxic exposures of various severities and durations demonstrated that key transcriptional regulators of metabolic and stress responses, hypoxia-inducible factor-1 (HIF-1) and/or nuclear factor kappa B (NFκB), were activated together with the production of inflammatory cytokines. 28 –30 Sustained hypoxia induced insulin resistance, 31 stimulated adipogenesis, 19,28 increased lipid synthesis, and stimulated spontaneous lipolysis in vitro 19 and in humans with chronic hypoxemia. 11 However, IH represents a more appropriate model of OSA, as periodic drops in hemoglobin saturation induce IH in various tissues, including adipose tissue. 32
Furthermore, accumulating evidence suggests that important differences in the cellular responses exist between sustained versus IH exposure, with preferential activation of NFκB, inflammatory pathways, and reactive oxygen species generation found with IH. 17,33 –37 In this study, cultured cells were exposed to shifts in pericellular O2 (5-min cycles, 16% to 1%) for 14 days, approximating adipocyte responses in the context of OSA. Prolonged exposure plays an important role as multiple cellular responses, including expression of glucose transporter type 4 and HIF-1 activity, exhibited significant temporal dependences with often biphasic patterns. 38 –40
Furthermore, previous studies showed that long-term, but not short-term, hypoxia stimulated lipolysis in 3T3-L1 adipocytes, 41 which was confirmed and extended in this study by showing that IH (lasting 24 hr) did not modify lipolysis, whereas a 14-day exposure promoted lipolysis. In addition, the effect of IH exposure on spontaneous lipolysis was only observed in human adipocytes derived from obese, but not nonobese, donors, suggesting that other mechanisms, for example, droplet size or genetic background are probably involved in the dysmetabolic effects of IH exposure in humans and that adipocytes of obese subjects are more susceptible to the effects of IH, although precise mechanisms remain to be investigated in future studies.
IH throughout a 14-day period increased spontaneous lipolysis and simultaneously reduced intracellular triglyceride stores. The demonstration of increased lipolysis is of particular interest, as augmented adipose tissue lipolysis contributes to the plasma pool of FFA, representing a causal factor in the development of muscle insulin resistance, pancreatic dysfunction, and the eventual development of type 2 diabetes. 42 Replicating identical exposure in differentiated preadipocytes derived from obese or nonobese human subjects, we found that only adipocytes from obese individuals showed increased lipolysis after IH exposure, suggesting an important interaction between increased triglyceride stores and the ability to increase lipolysis after hypoxic stimulus, particularly when most OSA patients are obese. 43
Importantly, pharmacological inhibition of lipolysis efficiently mitigated metabolic impairments induced by IH in a mouse model of OSA syndrome. 14 Among the mechanisms that possibly stimulate lipolysis under IH conditions, increased paracrine secretion of lipolysis-stimulating cytokines, for example, tumor necrosis factor-α and interleukin-6, 15 could be suggested, together with lipid oxidation downregulation, thus rendering more FFA available for extracellular transport and release. 44
It should be noted, that spontaneous lipolysis was unaffected by shorter 24-hr IH exposure, expanding previous observations, that short term effects of IH exposures are likely to be mediated by direct sympathetic activation, 45 whereas chronic IH exposure triggers different mechanisms, for example, changes in adipose tissue gene expression. 40 However, it should be noted that, in this study, effects of 24-hr IH exposure were not investigated in human obese and nonobese differentiated preadipocytes, preventing thus firm conclusions and projection of rodent data to human pathophysiology.
In contrast to sustained hypoxia, which was reported to increase intracellular lipid stores and lipid droplet sizes, 19,46 IH exposure in this study reduced intracellular lipids. This observation suggests an impairment in lipid synthesis/storage or reduced secretion into the extracellular space (culture media) and underscores fundamental differences between sustained and IH exposures. Although we did not quantify plasma FFA transport into the cells, reduced FFA uptake was previously documented in adipose tissue as a consequence of sustained hypoxia 47 or reduced expression of CD36, a major FFA transporter protein. 48 Diminished FFA transport into adipocytes is also supported by our current finding of reduced expression of the intracellular FFA-carrier protein, FABP, after IH exposure.
As opposed to sustained hypoxia, 19 de novo lipogenesis (replenishing intracellular lipids) was unaffected by IH as documented by the lack of responses to established inhibitors/activators of acetyl-CoA production from glycolysis or the Krebs cycle. Furthermore, the expression of genes required for triglyceride synthesis (FASN and DGAT) and markers of adipocyte differentiation was reduced by IH in this study, probably through peroxisome proliferator-activated receptor γ (PPARγ) suppression. 49,50 The overall balance of intracellular lipids was thus shifted negatively: diminished by ongoing lipolysis, but not replenished by extracellular transport and/or de novo lipogenesis.
In conclusion, prolonged exposure of adipocytes to IH increased lipolysis in 3T3-L1 as well as human adipocytes differentiated from obese, but not from nonobese, subjects. However, in contrast to sustained hypoxia, IH reduced intracellular triglyceride stores presumably through mechanisms involving reduced de novo lipid synthesis and reduced extracellular FFA transport. These findings demonstrated direct effects of IH in adipocytes and identified lipolysis as a possible pharmacological target for interventions aimed at improving metabolic impairments associated with OSA.
Footnotes
Acknowledgment
The authors thank Sarka Fleischerova for technical assistance.
Authors' Contributions
M.M. and M.W. cultured cells and conducted exposures to intermittent hypoxia and pharmacological exposures, M.K. performed gene expression analysis, J.P. developed the project, supervised experiments, and performed statistical analysis. All authors participated in article preparation and editing.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was supported by grants from Charles University, Prague (GAUK no. 18217) and the Czech Science Foundation (GACR no. 18-10144S).
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.