
Abstract
Background:
Metabolic syndrome (MetS) is a group of signs and symptoms that are associated with a higher risk of type 2 diabetes mellitus and cardiovascular diseases. The major risk factor for developing MetS is abdominal obesity, which is caused by an increase in adipocyte size or quantity. Increased adipocyte quantity is a result of differentiation of stem cells into adipose tissue. Numerous studies have investigated the expression of key transcription factors, including PPARG and CEBPB during adipocyte differentiation in murine cells such as 3T3-L1 cell lines. To better understand the expression changes during the process of fat accumulation in adipose-derived stem cells (ASCs), we compared the expression of DYRK1B, PPARG, and ẟB in ASCs between the patient (harboring DYRK1B R102C) and control (healthy individuals) groups.
Methods:
Gene expression was evaluated on the eighth day before induction and days 1, 5, and 15 postinduction. The pluripotent capacity of ASCs and the potential for differentiation into adipocytes were confirmed by flow cytometry analysis of surface markers (CD34, CD44, CD105, and CD90), and Oil Red O staining, respectively. The Expression of DYRK1B, PPARG, and CEBPB were assessed by real-time-polymerase chain reaction in patients and normal individuals. The effects of AZ191, a potent small molecule inhibitor on DYRK1B and CEBPB expression in patients' samples were studied.
Result:
The expression of DYRK1B kinase and transcription factors (CEBPB and PPARG) are higher in ASCs harboring DYRK1B R102C compared with noncarriers on days 5 and 15 during adipocyte differentiation. These proteins may be helpful to elucidate the mechanisms underlying obesity and obesity-related disorders like MetS. Furthermore, the new compound AZ191 exhibited inhibitory activity toward DYRK1B and CEBPB. We suggest that AZ191 may be helpful in defining the potential roles of DYRK1B and CEBPB in adipogenesis.
Introduction
Metabolic syndrome (MetS) or X syndrome is a group of signs and symptoms, comprising abdominal obesity, elevated blood pressure, dyslipidemia, and insulin resistance. MetS is roughly related to a twofold increase in the risk of cardiovascular diseases (CVDs) and a fivefold increased risk of type 2 diabetes mellitus. 1 The major triggering risk factor for MetS, in addition to insulin resistance, is central obesity, which is measured as waist circumference (WC). Obese individuals are at a higher risk of unpleasant health problems, such as CVDs, type 2 diabetes, and other adverse disorders. Depending on various criteria, including place of residence, ethnicity, and the definition used for diagnosis, the prevalence of MetS is estimated to be ranging from <10% to as high as 84% around the world. 2,3 The prevalence of MetS is relatively high in Iran, with estimates ranging from 20% to 25%. Its prevalence increases with increasing BMI (body mass index) and age. It has been reported that women are at higher risk for MetS than men. 4
Several variants in DYRK1B have been reported. Among these variants, NM_004714.3:
The DYRKs (dual-specificity tyrosine-phosphorylated kinases) are a highly conserved family of protein kinases. In mammalians, DYRKs are divided into two classes. Class I (DYRK1A and DYRK1B) and class II (DYRK2, DYRK3, and DYRK4). 6 These proteins participate in many vital processes such as metabolism, gene expression, and signaling pathways. 7,8 Dysregulation of these kinases has been linked to multiple conditions, including diabetes and CVDs. As a result, they have been suggested as powerful targets for the treatment of MetS. 7,9,10 DYRK1B participates in differentiation and undergoes different splicing during fat formation. 11 Moreover, DYRK1B is overexpressed and mutated in cancers. 12 –14 DYRK1B is activated by autophosphorylation on tyrosine-273 (Y273) within the catalytic domain.
It has been demonstrated that several small molecules such as INDY and Harmine have inhibitory effects on class I DYRKs (DYRK1A and DYRK1B). 15,16
The new compound AZ191 is a DYRK1B-selective inhibitor. AZ191 like Harmine selectively inhibits Ser/Thr kinase activity at 1 μM concentration. 11
Gain of function mutation, DYRK1B R102C, promotes adipogenesis and causes MetS by increasing obesity. 5,7,17 Accumulation of adipose tissue results in obesity. 18 Adipocyte differentiation is a well-organized multistep process requiring the successive activation of several groups of transcription factors, including CCAAT/enhancer-binding protein (CEBP) family and peroxisome proliferator-activated receptor-γ (PPAR-γ). During adipocyte differentiation, C/EBPD/B is expressed early and induces the expression of C/EBPα and PPARγ, two master transcription factors for terminal adipocyte differentiation. Numerous studies have utilized preadipocyte cell lines like 3T3-L1 or 3T3-442A to evaluate molecular mechanisms affecting adipogenesis.
The culture medium used in this investigation contains the enhancers of adipogenesis such as isobutyl-methylxanthine (IBMX) and dexamethasone (DXM), which regulate PPARG. Furthermore, IBMX and DXM are stimulators of transcription factors C/EBPβ and C/EBPẟ, which are essential for differentiation.
19
In the present study, we focus on DYRK1B, PPARG, and CEBPB expression alterations in adipose tissue-derived stem cells harboring
Materials and Methods
Adipose-derived stem cell culture, immunophenotypic characterization, and induction of adipogenic differentiation
Abdominal subcutaneous adipose tissue was obtained from three adult patients harboring the variant (labeled as A, B, and C) and three adult normal individuals (labeled as D, E, and F) under local anesthesia by a board-certified surgeon.
Cells from the affected and unaffected participants were cultured in a complete culture medium encompassing Dulbecco's modified Eagle's medium (DMEM) (GIBCO; Life Technologies™, New York) complemented with 10% fetal bovine serum (FBS; Sigma-Aldrich, St. Louis, MO), 1% (v/v) penicillin (10,000 U/mL)–streptomycin (10,000 μg/mL) (p/s; GIBCO; Life Technologies), and 1% nonessential amino acids in 75-cm2culture flasks (NUNC™, Roskilde Site, Kamstrupvej, Denmark) under humidified standard condition at 37°C in a 5% CO2 incubator. When the cells were 80% confluent, they were washed twice with 3 mL phosphate-buffered saline (PBS) containing 2% (v/v) p/s and separated from the bottom of the culture flask by adding 1 mL trypsin (GIBCO; Life Technologies). The cells were incubated at 37°C in 5% CO2 for about 7–10 min to ensure that the cells were separated. Trypsin was deactivated by adding the same amount of complete medium. The suspension was centrifuged at 1200 rpm for 5 min to pellet the cells.
Surface marker characterization
Mesenchymal stem cells (MSCs) were characterized by the analysis of expression of cell surface markers with flow cytometry at the third passage. The cell suspension (1 × 106 cells/mL) was washed in a blocking solution, cold PBS containing 10% FBS, for 20 min. Subsequently, the cells were labeled with Fluorescein isothiocyanate-conjugated anti-CD44, anti-CD90, Phycoerythrin-conjugated anti CD34, and PerCP (Peridinin Chlorophyll Protein complex)-conjugated anti-CD105 antibodies (Abcam, Cambridge, UK). They were then washed with cold PBS, resuspended in PBS containing 10% FBS, and analyzed by flow cytometry (FACSCalibur™; BD Biosciences, San Jose, CA). The data were analyzed utilizing WinMDI 2.9.
Adipogenic differentiation
For adipogenic differentiation, cells were seeded at a density of 5000 cells/cm2 in a complete culture medium and grown until they were 80% confluent. The cell cultures were washed twice with 3 mL PBS containing 2% (v/v) p/s and differentiated by adding an adipogenic induction medium to the cell cultures. The adipogenic medium contained DMEM (GIBCO; Life Technologies) completed with 10% FBS, 0.5 Mm IBMX, 1 μM DXM, 10 μM insulin, and 200 μM indomethacin (Sigma-Aldrich). Cells from passages 3 to 5 were used. The process of differentiation was completed during a 15-day period. The culture medium was replaced every 3 days. Cells were harvested at three different time points, days 1, 5, and 15, and then were used for RNA isolation.
Oil Red O staining
The completion of adipogenesis was confirmed by Oil Red O, which stains intracellular triglyceride droplets. After 10 days of differentiation start, the cells were fixed with 10% formalin and then incubated for 15 min with Oil Red O solution. Thereafter, the cells were washed three times with distilled water. The dye was eluted from the cells using isopropanol. The nuclear staining was carried out for 5 min using a stock solution of Hematoxylin–Eosin in water (10 mg/mL) diluted1:20 in PBS, with an aliquot added to a final concentration of 5 μg/mL Hematoxylin–Eosin. The cells were then washed three times with PBS.
RNA isolation and real-time quantitative polymerase chain reaction
Total cellular RNA was isolated from the control and patient cells by the RNA Extraction Kit (Cinnagen, Inc., Tehran, Iran). The quantity and quality of obtained RNA were checked using a Nanodrop™ spectrophotometer (3411 Silverside Rd, Bancroft Building, Wilmington DE, 19810) and agarose gel electrophoresis, respectively. Samples were stored at −80°C until cDNA synthesis.
The cDNA was synthesized using 500 ng of total RNA in a first-strand complementary DNA synthesis reaction with the help of RevertAid™ First-Strand cDNA Synthesis kit (Fermentas, Inc., Schwerte, Germany). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using the 7500 Real-Time PCR System and the RealQ Plus 2 × Master Mix Green (Ampliqon Co., Denmark). Specific primers targeting DYRK1B, PPARG, and CEBPB were designed using AlleleID (version 7.5) (Table 1). Two hundred nanomolar of each primer (Table 1) was added to each reaction to target the specific sequence. The glyceraldehyde-3-phosphate dehydrogenase housekeeping gene was also used as an internal control of qPCR reactions. The qPCR was set at 15 min at 94°C followed by 45 cycles of 30 sec at 94°C, and 1 min at 60°C (Table 1).
Real-Time Polymerase Chain Reaction Primers Used in This Study
Statistical analysis
We performed all in vitro experiments in two groups (including three normal and three affected individuals). Data are expressed as mean and standard errors (Table 2).
The Demographic and Clinical Characteristics of the Patients
BMI, body mass index; WC, waist circumference.
All individuals participating in this study are from a family living in southwest of Iran. Written informed consent for participation and publication was obtained from all of the individuals participating in this study, individually. The Ethics Committee of Shiraz University of Medical Sciences, Iran approved the present study (Ethics Code: IR. SUMS. REC. 1395.5604). This study was conducted according to the Declaration of Helsinki.
Results
Identification of cell surface markers
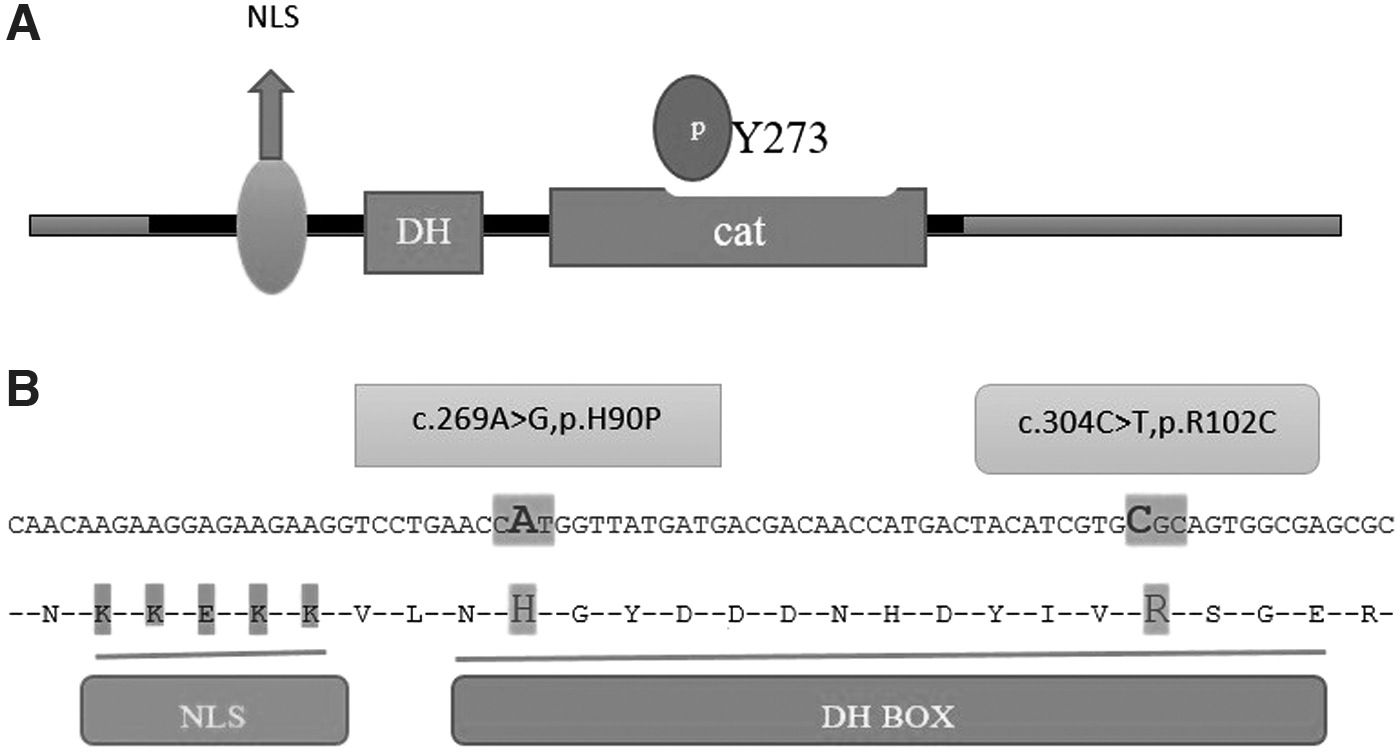
According to the International Society for Cellular Therapy, MSCs are stem cells with properties, including plastic adherence, they must express markers, including CD105, CD73, CD90, CD44, and lack expression of hematopoietic markers such as CD34, CD38, CD43, CD48, CD117, and CD150. These cells must have the capacity to differentiate into adipocytes, osteocytes, and chondrocytes. 20 Adipose-derived stem cells (ASCs) were isolated from six individual donors. The identification was based on the presence of CD44, CD105, and CD90 and the absence of markers specific to hematopoietic cell lines such as CD34. The expression of CD34 was as low as 9.65% and CD44, CD105, and CD90 were detected on the surface of 92.2%, 55.9%, and 97.0% of cells. These results are comparable with other studies and confirm the existence of MSCs. As shown in Fig. 2 blue is for positive expression of CD marker and red for negative control. 21 –23
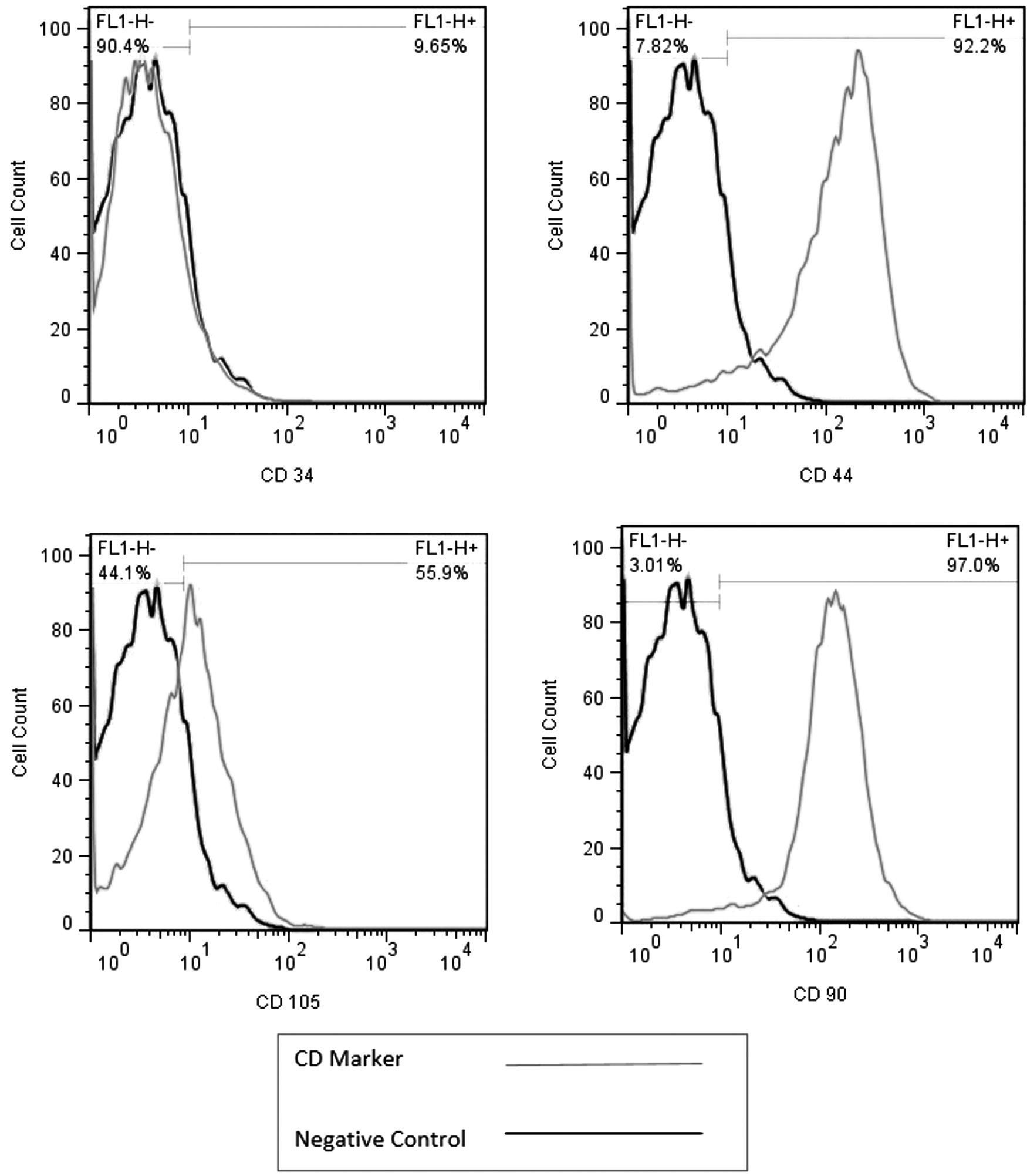
Flow cytometry results for adipose tissue-derived MSC, expressing CD44 (92.2%), CD90 (97%), and CD105 (55.9%) but low level of CD34 (9.65%). Black histogram is the negative control. Left side of the gray histogram (in the negative control part) represents the negative expression of CD marker; right side represents the positive expression of CD marker. MSC, mesenchymal stem cell.
Differentiation of AMSCs
As seen under Oil Red O and Hematoxylin–Eosin staining, there was an increased accumulation of lipid droplets in cells (red stain), with a remarkable increase in quantity and size of the droplets (Fig. 3). In the presence of adipogenic media, MSCs could store lipid droplets. These results confirmed the pluripotent capacity of MSCs. 24
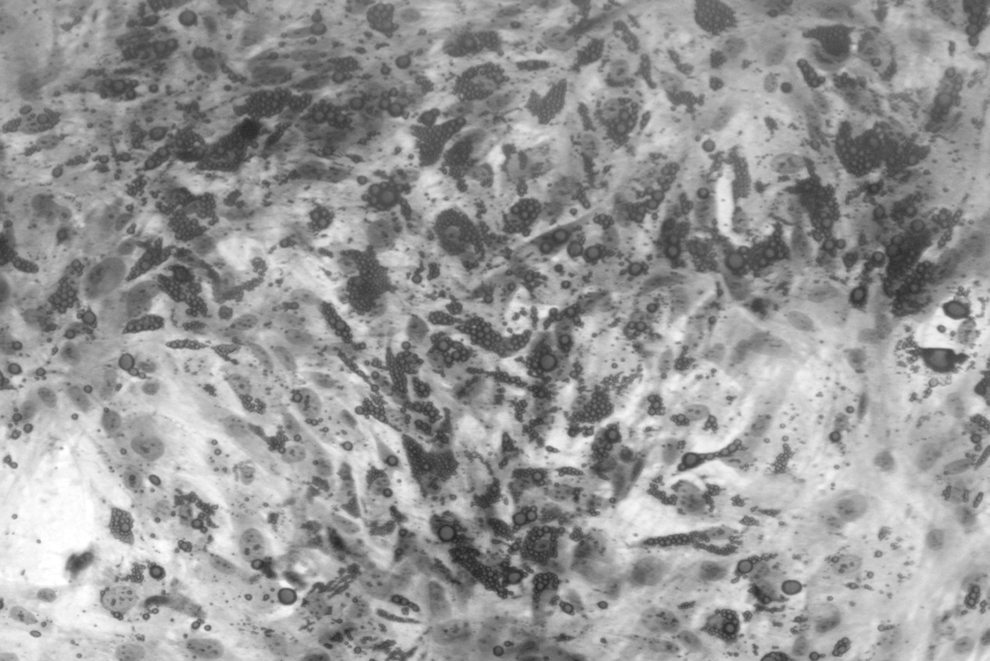
Microscopy image depicting adipocytes after differentiation. The lipid droplets were stained with Oil Red O (dark gray stain). Nuclei were stained with Hematoxylin–Eosin (light gray stain).
Quantitative real-time polymerase chain reaction
DYRK1B is a member of the DYRKs family, comprising evolutionarily conserved protein kinases. 6 DYRK1B is involved in several processes, including myogenesis, adipogenesis, carcinogenesis, and cell survival. 11 Antiadipogenic characteristics of Hedgehog signaling could increase the expression of their antiadipogenic target genes like Wnt. Activation of Wnt protein family hinders adipocyte differentiation. 18 Wnt signaling restrains adipocyte differentiation by inhibiting the expression of key adipogenic transcription factors, namely PPARG and CEBPA. The expression of these transcription factors is induced by CEBPB and CEBPD. 25 DYRK1B stimulates adipogenesis by inhibiting Sonic Hedgehog and Wnt pathways and inducing the expression of C/EBPα and PPARγ. 26
Hedgehog pathway activation includes the binding of extracellular Hh ligand to the complex made up of receptor Patched (Ptc) and Smoothened (Smo) protein. This leads Smo to release from Ptc. As a result, the Hedgehog pathway inhibits adipocyte differentiation by increasing the expression of antiadipogenic Wnt protein and reducing the expression of CEBP/α and PPARγ (Fig. 4). 18,27
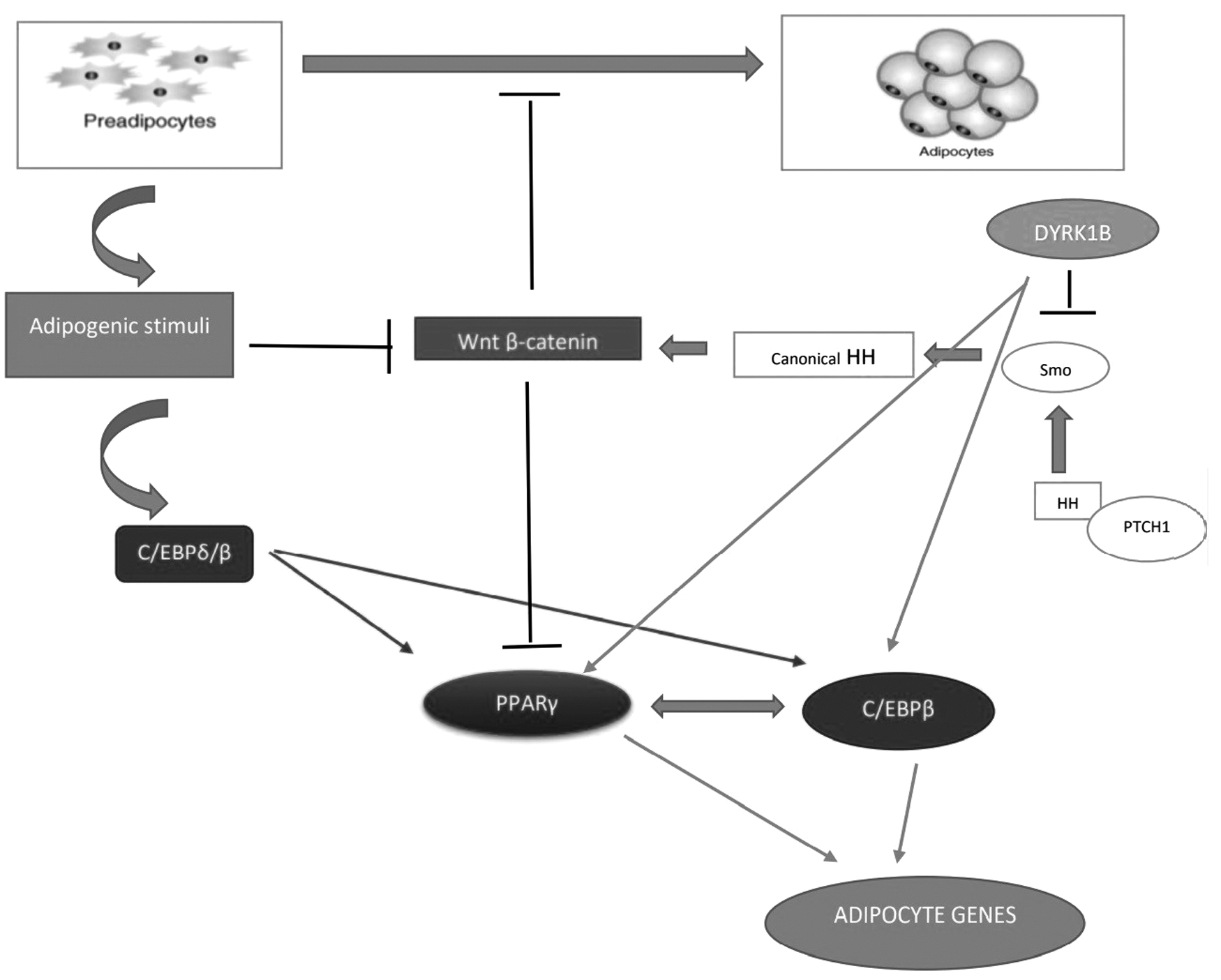
Summary of the function of DYRK1B, CEBPDB, CEBPB, and PPARG during adipogenesis.
To define the role of DYRK1B, PPARG, and CEBPB in adipocyte differentiation of ASCs, the expression profile was determined. Expression of CEBPB and PPARG in ASCs could be detected at the start of induction by real-time PCR. Expression of DYRK1B increased 5 days after induction and reached its maximum level after 15 days (Fig. 5).
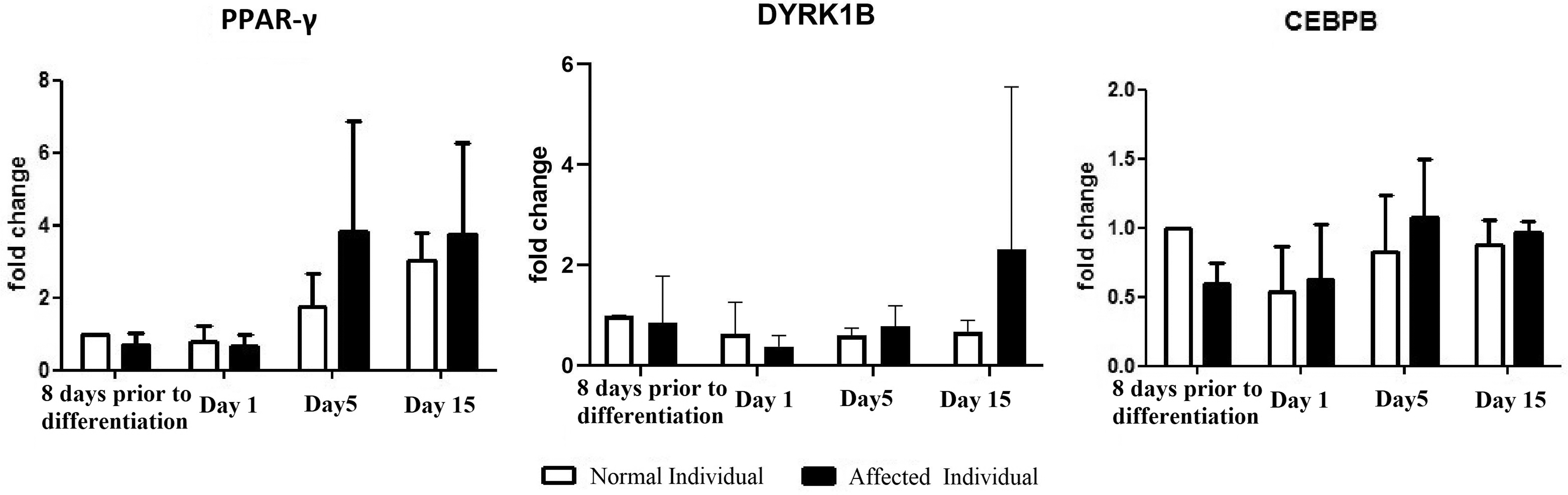
Relative expression levels of DYRK1B, PPARG, and CEBPB in samples obtained from affected and normal individuals. Expression values plotted are mean and standard error of the mean. DYRK1B, PPARG, and CEBPB expression were upregulated on days 5 and 15. The expression of DYRK1B, PPARG, and CEBPB were increased on the 5th and 15th days in comparison with the first day since the initiation of differentiation.
The expressions of DYRK1B, PPARG, and CEBPB were upregulated on days 5 and 15, indicating the maturation of the adipocytes. The expressions of DYRK1B, PPARG, and CEBPB were increased on the 5th and 15th days in comparison with the first day since the initiation of differentiation. A range of obesity-related diseases arise in the late stages of obesity, and most adipocytes in this stage have already been differentiated and maturated. Thus, the genes that were upregulated at the late stage of adipocyte differentiation are hypothesized to be related to lipid droplet accumulation and obesity-related diseases.
The expression of DYRK1B, PPARG, and CEBPB in the patients harboring the variant were higher in comparison to the controls. The expression patterns of DYRK1B, PPARG, and CEBPB in control and the patient's population are summarized in Fig. 5.
Effect of AZ191 treatment on the expression of DYRK1B and CEBPB
AZ191 inhibits the serine/threonine kinase activity selectively, but with no effect on tyrosine kinase autophosphorylation at 1 μM concentration. 28
In the present study, the effect of AZ191 on the expression of DYRK1B and CEBPB was assessed in the complete culture medium (control) and adipogenic medium with and without AZ191 at 3 μM concentration. Interestingly AZ191 treatment reduced the expression of DYRK1B and CEBPB after differentiation was induced (as shown in Fig. 6). Regarding the expression difference between control and adipogenic induction, DYRK1B and CEBPB expression were higher in the adipogenic medium (Fig. 6). Taken together, these results indicated that AZ191 is a potent inhibitor of DYRK1B and CEBPB.

Effect of AZ191 treatment on the expression of DYRK1B and CEBPB in various media in the cells harboring the variant (DYRK1B R102C) in the 10th day of differentiation. The expression of DYRK1B and CEBPB decreased with AZ191 treatment in comparison with adipogenic medium without AZ191.
Discussion
Despite the worldwide increasing prevalence of MetS, a suitable therapy has not been founded, yet. 29 In comparison to murine cell lines, human MSCs are more troublesome to manipulate, nevertheless, their usefulness and unique curative potential make them necessary to study. Moreover, differences in basic structure and regulation between human and rodent cells may affect gene expression. As a result, murine cells would be less reliable to model human diseases like obesity. 30
Presently, the literature regarding the role of CEBPB, PPARG, and DYRK1B in adipogenesis is focused on investigations on murine cells, and in vitro investigations, making the evidence from studies in ASCs quite rare. 31
Mutated DYRK1B (
To the best of our knowledge, this is the first study evaluating the possible role of CEBPB, PPARG, and DYR1B in the development and progression of the MetS. The genes, which are expressed during the late phase of adipocyte differentiation, may play a major role in promoting excessive proliferation and accumulation of lipid droplets, which contribute to the development of obesity and obesity-related disorders.
Previous studies showed that gain-of-function and loss-of-function mutations in PPARG lead to obesity and decreased body weight, respectively. 34 Besides PPARG, there is strong evidence that CEBPB plays an important role in adipocyte differentiation, associating it with obesity. 35 Wnt and Hedgehog signaling inhibit adipocyte differentiation through affecting PPARG and CEBPB, key regulators of adipogenesis. 18 Previous studies confirmed that the increased level of CEBPB results in inhibiting Wnt signaling. Moreover, CEBPB is capable of activating adipocyte maturation by inducing PPARG which in turn, leads to activating CEBPA. 17 In agreement with these studies, PPARG and CEBPB are significantly highly expressed during adipogenesis, defining their prominent roles in adipogenesis.
In this study, we have shown the importance of p.R102C in altering the expression levels of DYRK1B, PPARG, and CEBPB, which underlies the obesity and subsequent MetS in the patients harboring this variant. Our study, in addition to shedding light on the effect of the variant on the expression of DYRK1B, PPAR-γ, and CEBPB, shows the in vitro efficacy of AZ191 in downregulating the expression of DYRK1B and CEBPB, possibly counteracting the effects of the variant and the subsequent obesity it causes. Moreover, new biological treatments, including cell therapy, gene therapy, iRNA, etc. can be utilized to treat those suffering from MetS caused by this variant. 36
The present study is mainly limited by the use of a small sample size. Due to the invasive nature of obtaining the biopsies, the samples size was limited to six individuals from a family. Furthermore, it should be noted that there was a disparity in the number of the women recruited in this study. Another limitation of the present study was not accounting for the possible effects of cell passage repetitions on the differentiation potential of ASCs. As a result, we suggest that the results of the present study need to be validated in future investigations.
Conclusions
In conclusion, the increased expression of CEBPB, PPARG, and DYRK1B in patients during adipogenesis and their contribution to MetS make them effective targets for new drug development efforts targeting obesity and obesity-related disorders. It is not clear whether DYRK1B variants cause MetS by inducing adipogenesis or through glucose production. Thus, clarifying the molecular mechanism of DYRK1B mutations and functional studies of these transcriptional factors is essential.
Footnotes
Authors' Contributions
This study was designed and conceptualized by A.A. and M.D. A.A., H.J.K., and S.Z. executed the study. M.D. supervised the execution of the study. A.A. drafted the article and S.Z. revised the final version. All authors approved the final version of the article.
Declarations
All of the listed authors confirm that this research is supported by Shiraz University of Medical Sciences, an institution primarily involved in education, research, and health care.
Ethics Approval and Consent to Participants
Written informed consent was obtained from each patient or, in the case of minors, from their parents. This study was approved by The Ethics Committee of Shiraz University of Medical Sciences.
Consent to Publish
Consent for participation and publication was obtained from the patients or, in the case of minors, from their parents.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was funded by National Institute for Medical Research Development (NIMAD; grant no. 964030).