
Abstract
Adjuvants are important components of vaccine formulations. Effective adjuvants line innate and adaptive immunity by signaling through pathogen recognition receptors. Synthetic cytosine-phosphate-guanine (CpG) oligodeoxynucleotides (ODNs) have been shown to have potentials as adjuvants for vaccines. However, the immunostimulatory effect of CpG is species-specific and depends on the sequence of CpG motifs. A CpG ODN (2135), containing 3 identical copies of GTCGTT motif, was previously reported to have the strongest effects on bovine peripheral blood mononuclear cells (PBMC). Based on the sequence of 2135, we replaced the GTCGTT motif with 11 other sequences containing CG and investigated their effects on bovine lymphocyte proliferation. Results showed that the CpG ODNs containing 3 copies of GACGTT motif had the highest lymphocyte stimulation index (7.91±1.18), which was significantly (P<0.05) higher than that of 2135 (4.25±0.56). The CpG ODNs containing 3 copies of GACGTT motif also significantly increased the mRNA expression of interferon (IFN)-α, interleukin (IL)-12, and IL-21 in bovine PBMC. When dairy cows were immunized with the keyhole limpet hemocyanin (KLH) antigen formulated with CpG ODNs containing 3 copies of GACGTT, production of KLH-specific antibodies in serum and in milk whey was significantly (P<0.05) enhanced. IFN-γ in whole blood stimulated by KLH was also significantly (P<0.05) increased in cows immunized with KLH plus CpG ODNs. Our results indicate that CpG ODNs containing 3 copies of the GACGTT motifs is a potential adjuvant for bovine vaccines.
Introduction
The immune responses elicited by CpG ODNs lead to the consideration of using CpG as an adjuvant. Formulation of CpG ODNs in vaccines dramatically increased the production of antigen-specific antibodies and enhanced Th 1 type immunity as characterized by an increased production of IgG2, secretion of IFN-γ, and lytic activity of cytotoxic T lymphocytes in mice (Klinman et al., 1999; Hunolstein et al., 2000; Chen et al., 2004). Other studies also demonstrated that CpG ODNs, when used as an adjuvant, are less toxic and induce less undesired side effects at the site of injection without compromising the production of IgG2 in comparison with other adjuvants (Chu et al., 1997; Weiner et al., 1997; Weeratna et al., 2000).
The response of mammalian cells to CpG ODNs is species-specific, presumably due, as least in part, to the recognition by TLR9 in different species. It is in general agreement that the optimal motif for mice and birds is GACGTT, and the most stimulatory motif for other species, including human, bovine, ovine, and swine, is GTCGTT (Zhang et al., 2001, 2008). These claims remain controversial. The first immunostimulatory CpG motif for bovine species, AACGTT, was identified in 1998 (Brown et al., 1998). A sequence containing AACGTT motifs has been demonstrated to increase the secretion of IL-12, tumor necrosis factor (TNF)-α, and nitric oxide in bovine macrophages (Shoda et al., 2001). After testing a panel of 38 CpG ODNs, the sequence that effectively stimulated human cells (2135=TCGTCGTTTGTCGTTTTGTCGTT) was shown to be the optimal sequence for bovine cells in terms of lymphocyte proliferation response and IFN-γ secretion, a marker of Th1 type responses (Pontarollo et al., 2002). However, these CpG ODNs, with different motifs, were not compared based on the same sequence and copies of motifs, which might lead to the biased conclusion that GTCGTT is the optimal motif for bovine cells. In the present study, 11 different hexamer CpG motifs were used to replace GTCGTT in 2135 without changing the flanking sequences and their effects on proliferation of bovine peripheral blood mononuclear cells (PBMC) were analyzed. The CpG ODNs that had the highest stimulation index (SI) in the proliferation assay was further examined for its ability to upregulate the expression of selected cytokine genes by real-time PCR and for adjuvant activity in cattle vaccinated with a model antigen, keyhole limpet hemocyanin (KLH).
Materials and Methods
In vitro studies
CpG oligodeoxynucleotides
The ODNs were synthesized (Mission Biotech) based on the sequence of ODN 2135 (TC-GTCGTT-T-GTCGTT-TT-GTCGTT), which was previously demonstrated to induce strong immunostimulatory effects on bovine PBMC (Pontarollo et al., 2002). Totally 11 different motifs were randomly selected to replace GTCGTT. All CpG ODNs contained 3 copies of the active motif on the nuclease-resistant phosphorothioate backbone (Table 1). A sequence of ODN containing 3 copies of GTGCTT was used as the control CpG (Ctrl ODN). All ODNs were resolved in sterile nonpyrogenic water at a concentration of 250 μM and stored at −70°C.
CpG, cytokine-phosphate-quanine; ODN, oligodeoxynucleotides.
Isolation of PBMC
Bovine PBMC were isolated according to the method previously described (Mehrzad et al., 2008) with modifications. Briefly, Heparin-anticoagulated blood was mixed with an equal volume of Ca2+-free Hank's balanced salt solution (HBSS; Gibco/BRL), carefully layered onto Ficoll-Paque Plus (GE Healthcare), and centrifuged at 1500 g for 40 minutes at 4°C. The buffy coat was washed with HBSS and suspended in RPMI (Roswell Park Memorial Institute) 1640 (Gibco/BRL) supplemented with 10% heat inactivated fetal bovine serum (FBS; Gibco/BRL), 1% antibiotic–antimycotic solution (Sigma-Aldrich Co.), and 1%
Lymphocyte proliferation assay
Lymphocyte proliferation assay was carried out as described (Mehrzad and Zhao, 2008) with modifications. Freshly isolated PBMC were suspended in RPMI 1640 supplemented with 10% heat inactivated FBS), 1% antibiotic–antimycotic solution, 1%
In vitro stimulation, RNA extraction, and reverse transcription
Bovine PBMC were added to 24-well plates (3×106 cells/well) with or without selected CpG ODNs (3 μM) and incubated at 37°C, 5% CO2, for 6 hours. PBMC were centrifuged and total RNA was extracted using Trizol (Invitrogen) according to the manufacturer's instructions. The reverse transcription (RT) reaction was started by adding 2 μg of total RNA and 0.5 μg of oligo(dT12-18) to 12 μL of water, then heating at 70°C for 10 minutes. After the samples were cooled on ice, 10 mM dithiothreitol, 0.5 mM of each deoxynucleotide triphosphate (dNTP), 5× first strand buffer, and 200 U Superscript II RNase H− Reverse Transcriptase (Gibco/BRL) were added. The mixture was stabilized at 25°C for 10 minutes and subsequently incubated at 42°C for 50 minutes for the RT reaction to proceed. Thereafter, the temperature was raised to 70°C for 15 minutes to inactivate the reverse transcriptase. Synthesized cDNA was examined for its quality and stored at −20°C until being used for real-time PCR.
Quantitative real-time PCR
Amplification by LightCycler real-time PCR was carried out as described (Lee et al., 2006). Primers for specific bovine genes are listed in Table 2. The reaction conditions for each gene were optimized using a QuantiTect SYBR Green PCR kit (Qiagen) in a LightCycler system (Roche) and applied to the following protocol. The cDNA was analyzed in 20 μL PCR mixture containing a final concentration of 0.5 μM primer, 1 μL of cDNA, and 2× QuantiTect SYBR green PCR master mix. The PCR master mix contained HotStartaq DNA polymerase, SYBR green PCR buffer, and dNTP mix including dUTP, SYBR green I, ROX (passive reference dye), and MgCl2 (2.5 mM). The PCR mixture was added into a cold PCR capillary, centrifuged, and placed in the LightCycler system. The LightCycler was programmed in 3 steps: (1) denaturation at 95°C for 15 minutes; (2) amplification for 50 cycles of denaturation at 94°C for 15 seconds, annealing at 60°C for 30 seconds, and extension at 72°C for a duration depending on the product length (5 seconds per 100 bp); and (3) melting curve analysis at 95°C for 5 seconds, 65°C for 15 seconds and 95°C for 40 seconds, followed by cooling at 40°C. In each reaction, the cycle number at which the fluorescence rose appreciably above background fluorescence was determined as the crossing point (CP).
IFN, interferon; IL, interleukin; TLR9, Toll-like receptor 9.
Relative quantification
The expression of each gene was analyzed using a relative quantification method. A slope was determined from the exponential phase, under the optimized real-time PCR amplification conditions, of each target gene or the reference gene (bovine β-actin). The amplification efficiency (E) was calculated based on the slope, where E=10[−1/slope]. The expression of selected genes was calibrated to that of β-actin, for each treatment and converted to the relative gene expression ratio, where
In vivo studies
Animals
Sixteen 2–3-year-old, pregnant, and clinically healthy Holstein cows in their late stage of lactation were selected based on milk somatic cell counts (<500,000 cells/mL) and the absence of bacteria in aseptically collected milk samples. All cows were housed in a free-stall enclosed barn (located at Pingtung, Taiwan) and milked twice daily. The use and care of all animals in this study were approved by the Animal Care and Use Committee of National Pingtung University of Science and Technology.
Immunization protocol
Vaccines were prepared by emulsifying Freund's incomplete adjuvant (FICA; Sigma) with an equal volume of KLH (Sigma) with or without CpG ODNs in nonpyrogenic water (Table 3). The total volume for each dose of immunization was 2 mL. Animals were randomly assigned into 4 groups (n=4 each): (1) FICA (the control); (2) KLH+FICA; (3) KLH+FICA+CpG; (4) KLH+FICA+cCpG. Cows received the primary immunization (D0) subcutaneously at the right side of the neck and were boosted with the same formulation 2 weeks later (D14) at the left side of the neck. Serum samples were prepared from blood collected on D0, D7, D14, D21, D28, D35, D70, and D140 for analysis of KLH-specific antibody. Heparin-anticoagulated whole blood was collected from the tail vein on D0, D14, and D28 for PBMC isolation and IFN-γ assay. Milk samples (20 mL) were collected on D0, D7, D14, D21, D28, and D35 before the cows moved to the dry period. Milk samples were poured into 40 mL round-bottom centrifuge tubes and centrifuged at 44,000 g for 30 minutes at 4°C. After centrifugation, the fat layer was removed and the skimmed milk was carefully decanted into new centrifuge tubes followed by centrifugation at 44,000 g for 30 minutes at 4°C. The clear whey was collected and stored in aliquots at −20°C.
CpG ODN=TC-GACGTT-T-GACGTT-TT-GACGTT.
cCpG ODN=TC-GAGCTT-T-GAGCTT-TT-GAGCTT.
cCpG, control CpG; FICA, Freund's incomplete adjuvant; KLH, keyhole limpet hemocyanin.
Detection of KLH-specific antibodies in serum and milk
The antibody titer was measured by ELISA as described (Lee et al., 2005) with modifications. Briefly, 96-well flat-bottom plates were coated with KLH (1 μg/mL) and incubated overnight at 4°C. Then, the coating solution was removed followed by blocking the plates with 2% gelatin (Sigma) in PBS at 37°C for 1 hour. After washing the plates twice with PBS-T (containing 0.05% Tween 20), samples diluted with PBS-T (1:10,000 and 1:100 for serum and whey samples, respectively) were added into the wells. The plates were incubated at 4°C overnight. Thereafter, the plates were washed 3 times, and incubated with rabbit anti-bovine IgG1, IgG2, or IgM (Bethyl Laboratories, Inc.), diluted in PBS containing 0.2% Tween 20 for 1 hour at 37°C. After washing, alkaline-phosphatase-labeled sheep anti-rabbit IgG (1:2000) (Kirkegaard and Perry Laboratories, Inc.) was added to the wells and incubated for another 1 hour at 37°C. Substrate was added and the plates were read at OD405 nm.
Interferon-γ
A bovine IFN-γ ELISA kit (BioSource International) was used to determine the production of IFN-γ in whole blood in response to KLH. Briefly, 100 μL of KLH in RPMI 1640 (150 μg/mL) was added to 500 μL whole blood from each cow. The mixture was incubated at 37°C, 5% CO2, for 24 hours and centrifuged at 1700 g, 4°C, for 20 minutes. The supernatant was collected for analyzing the level of IFN-γ according to the manufacture's instructions.
Statistical methods
All data were analyzed by the statistical software, SAS (Version 9.0). For lymphocyte proliferation assay and the relative gene expression ratio, differences between the treatments were analyzed by analysis of variance (ANOVA) and compared by Duncan's multiple comparison. For the antibody responses and IFN-γ secretion, data were analyzed by ANOVA and Duncan's multiple comparison was used to test the significance between treatment groups throughout the study and at each time point. A P value of <0.05 was considered significant.
Results
In vitro studies
Lymphocyte proliferation in response to different CpG motifs
CpG ODNs containing 3 copies of GTCGTT, AGCGTT, GGCGTT, and GACGTT motif significantly (P<0.05) increased bovine lymphocyte proliferation in comparison with the Ctrl ODN (Fig. 1). GACGTT motif had the highest SI (7.91±1.18) and was significantly (P<0.05) higher than that of GTCGTT (SI=4.25±0.56).
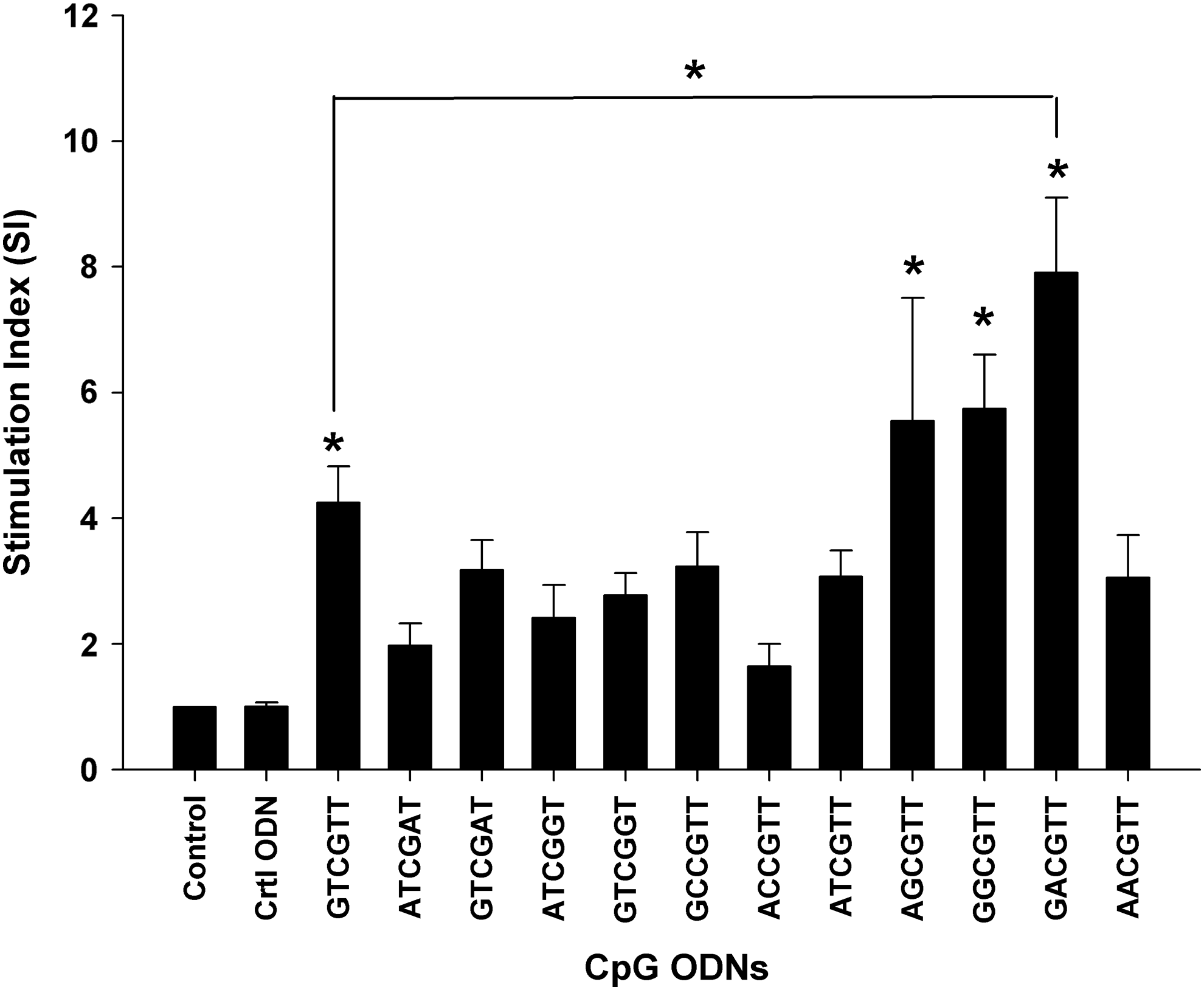
Stimulation of bovine lymphocyte proliferation by CpG oligodeoxynucleotides (ODNs) (3 μM) containing different motifs as listed in Table 1. The stimulation index (SI) was calculated by dividing the mean counts per minute (CPM) of each treatment by that of the medium control. Data are represented as the mean SI±SEM of 8 cows. *The mean SI is significantly different from that of the control ODN (Ctrl ODN) at P<0.05.
Upregulation of cytokines and TLR9 induced by GTCGTT and GACGTT CpG ODNs in bovine PBMC
As shown in Fig. 2, AGCGTT and GGCGTT CpG ODN did not upregulate mRNA expression of these genes. GTCGTT CpG ODN only significantly (P<0.05) increased the expression of IL-12 in comparison with the control. On the other hand, GACGTT CpG ODN significantly (P<0.05) upregulated the expression of IFN-α, IL-12, and IL-21. Neither IFN-γ nor IL-6 mRNA expression was significantly increased by the CpG ODNs. The PBMC stimulated by GACGTT CpG ODN had a significantly (P<0.05) increased gene expression of TNF-α and TLR9. However, the increase was relatively small (relative gene expression ratio was <2).

The expression of cytokine and Toll-like receptor 9 (TLR9) genes in bovine peripheral blood mononuclear cells (PBMC) stimulated by CpG ODNs containing 3 copies of GTCGTT, AGCGTT, GGCGTT, or GACGTT motif. Freshly isolated bovine PBMC were incubated with each CpG ODN (3 μM) for 6 hours. The expression of the target gene was calibrated by that of the reference gene, bovine β-actin, and converted to the relative gene expression ratio. Data are presented as the mean±SEM. of 5 cows and only means >2 are valid for statistical analysis. Treatments with different superscript letters in the same group are significantly (P<0.05) different. The group of bars without superscript letters indicates that the treatments were not significantly different.
In vivo studies
Antibody responses against KLH antigen in serum
The CpG ODN containing 3 copies of GACGTT motif was formulated with KLH antigen to examine its effect on antibody responses and antigen-induced IFN-γ secretion in dairy cattle. The responses of KLH-specific antibodies, including total IgG, IgG1, IgG2, and IgM, in serum were illustrated in Fig. 3. In cows immunized with KLH-antigen, all the subclasses of KLH-specific antibody peaked on D21 or D28, which were 1 or 2 weeks after the boost (D14). Formulation of CpG ODN containing 3 copies of GACGTT motif with KLH antigen significantly (P<0.05) increased the production of KLH-specific total IgG, IgG1, IgG2, and IgM in serum in comparison with other formulations and the control. Moreover, the antibody response (for all subclasses) in cows immunized with KLH+FICA+CpG was more persistent, which remained significantly (P<0.05) higher than other treatments and the control until the end of the study (D140), except IgG1. The antibody response of cows immunized with KLH+FICA+cCpG was not significantly different from that of cows immunized with KLH+FICA. In addition, cows immunized with KLH+FICA or KLH+FICA+cCpG did not have an increased production of KLH-specific IgG2 in serum.
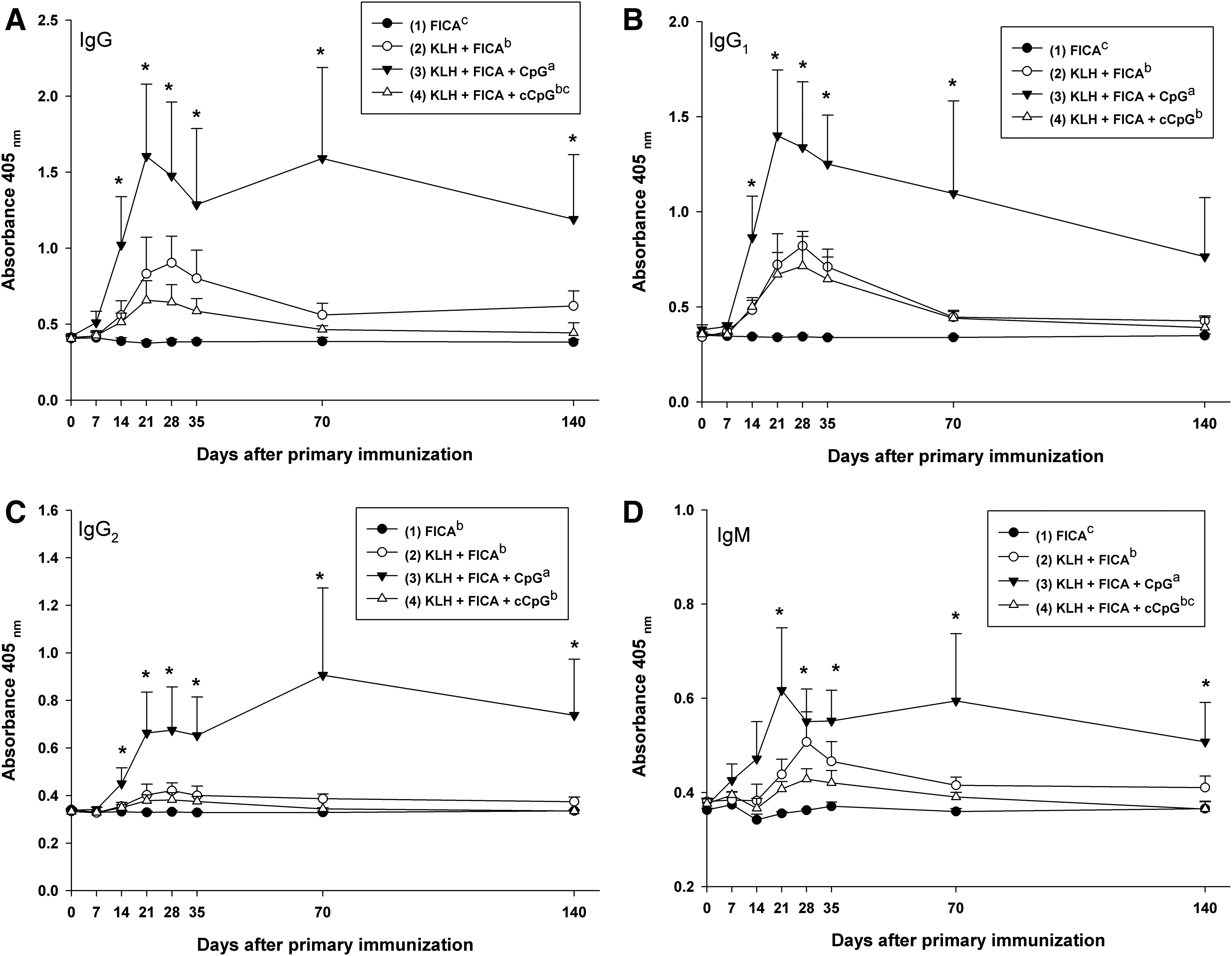
The response of keyhole limpet hemocyanin (KLH)-specific
Antibody responses against KLH antigen in milk
The response of KLH-specific antibodies, including total IgG, IgG1, IgG2, and IgM, in milk was illustrated in Fig. 4. Formulation of CpG ODN containing 3 copies of GACGTT motif with KLH antigen significantly (P<0.05) increased the production of KLH-specific total IgG, IgG1, and IgM in milk in comparison with other formulations and the control. Cows vaccinated with KLH+FICA or KLH+FICA+cCpG only significantly (P<0.05) increased the production of KLH-specific IgG1 in milk. None of the formulations increased the production of KLH-specific IgG2 in milk.
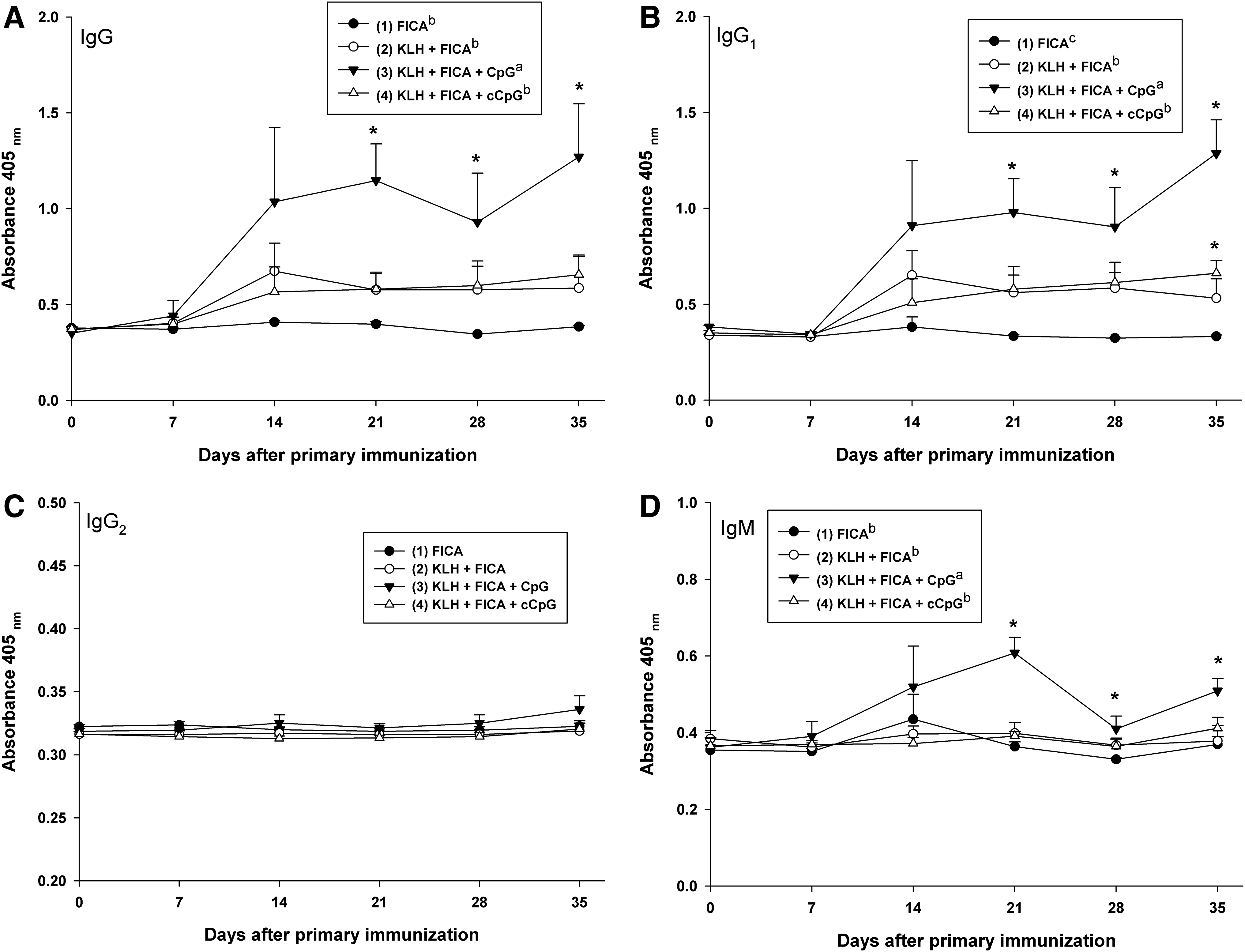
The response of KLH-specific
Production of IFN-γ in whole blood incubated with KLH
Antigen-specific secretion of IFN-γ in whole blood was measured on D0, D14, and D28 using a commercially available ELISA kit. As shown in Fig. 5, the secretion of IFN-γ in whole blood after KLH stimulation was significantly (P<0.05) increased in cows immunized with KLH+FICA+CpG on D14 in comparison with other formulations and the control. On D28, the secretion of IFN-γ in whole blood after KLH stimulation in cows immunized with KLH+FICA+CpG was still significantly (P<0.05) higher than those in cows received FICA and KLH+FICA+cCpG, but not KLH+FICA.
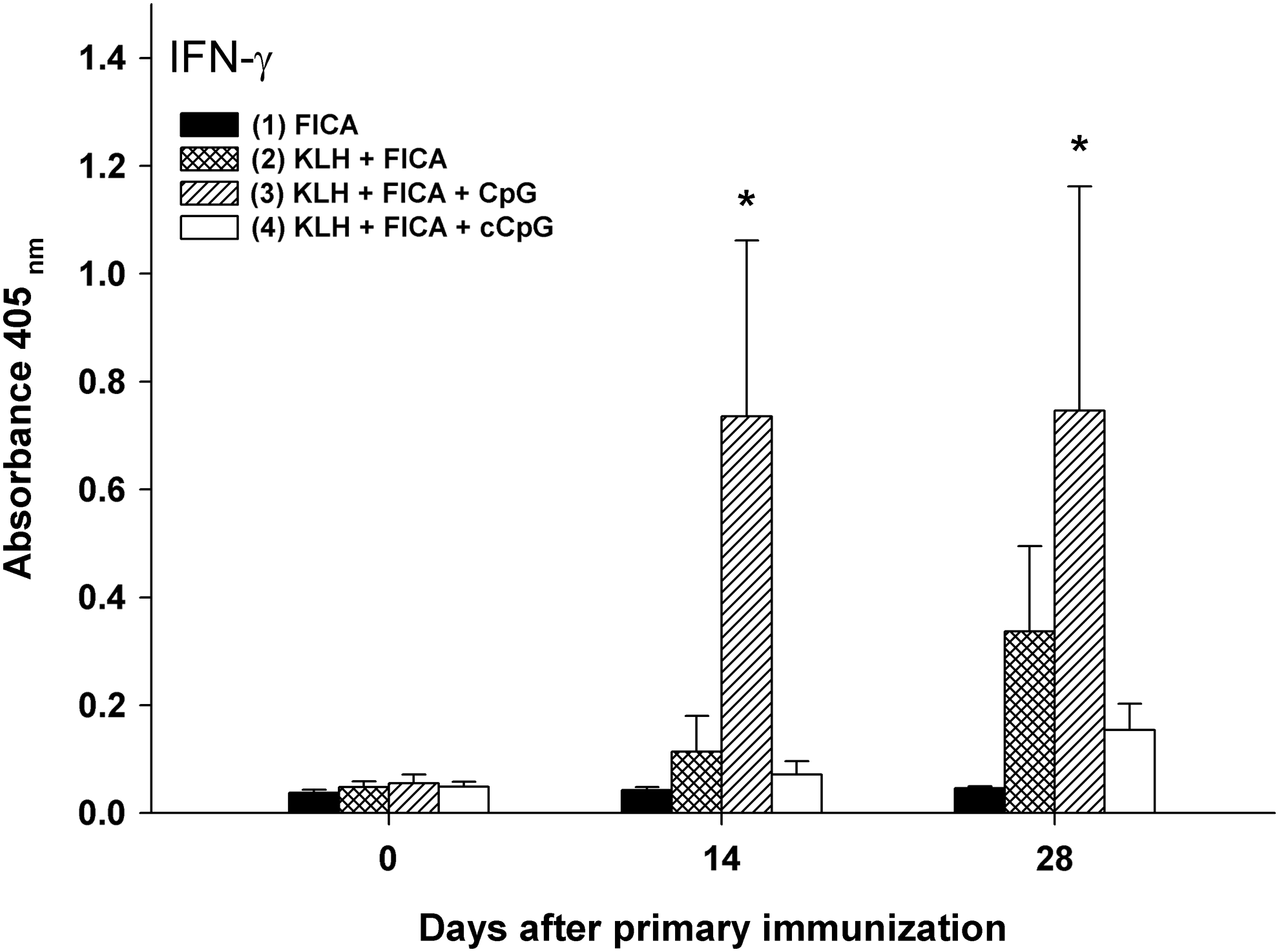
The expression of interferon (IFN)-γ mRNA in whole blood stimulated by KLH (25 μg/mL). Cows were immunized twice on D0 and D14 with (1) FICA, (2) KLH+FICA, (3) KLH+FICA+CpG, or (4) KLH+FICA+cCpG, and whole blood samples were taken on D0, D14, and D28. Data are presented as the mean±SEM of the OD values read at 405 nm from 4 animals. *The mean OD value is significantly (P<0.05) different from that of cows immunized with the adjuvant control (FICA).
Discussion
Eleven different motifs were used to replace GTCGTT without changing the flanking region in 2135 (TCGTCGTTTGTCGTTTTGTCGTT) and their stimulatory effects on bovine lymphocyte proliferation were examined based on the same general structure. Results showed that GACGTT had the highest SI (7.91±1.18), which was higher (P<0.05) than that of 2135 (4.25±0.56). CpG ODNs containing GACGTT were highly stimulatory for mice (Krieg et al., 1995) and rabbits (Rankin et al., 2001), and show only limited effects in humans (Hartmann and Krieg, 2000). Our results indicated that when all the CpG ODNs were synthesized with exactly the same structure, the one with 3 copies of GACGTT motif had the highest SI in the bovine lymphocyte proliferation assay. This surprising result warrants further studies.
Our results on cytokine and TLR9 gene expression by bovine PBMC also supported the notion that CpG ODN containing 3 copies of GACGTT motifs was more stimulatory than other CpG ODNs. GACGTT demonstrated stimulatory effects similar to those of 2135 (Fig. 2). In addition, GACGTT, but not 2135, significantly increased IFN-α mRNA expression even though the relative gene expression ratio was only slightly over 2. It is known that PBMC respond differently to various classes of CpG ODNs. The structure of CpG ODNs used in our study belongs to the K (or referred as B) type, which only induces minimum expression of IFN-γ and IFN-α in human and bovine PBMC (Verthelyi et al., 2001; Mena et al., 2003). The expression of IL-12 was significantly upregulated in PBMC stimulated by GACGTT and GTCGTT CpG ODNs. A stronger induction of IFN-α and IL-12 may be beneficial to enhancing Th1 type immune responses when the CpG ODN is formulated to a vaccine as an adjuvant.
To the best of our knowledge, this is the first investigation on the expression of IL-21 in response to CpG ODN stimulation. IL-21 is a crucial cytokine for the function of B cells, the cytotoxicity of lymphocytes, and the development and differentiation of NK cells (Parrish-Novak et al., 2000; EBERT, 2009; Konforte et al., 2009). CpG ODNs containing GACGTT, but not GTCGTT, motifs significantly increased the expression of IL-21. For IL-6, the induction of mRNA in CpG ODN-stimulated bovine PBMC was not significant in our assay. The expression of IL-6 in bovine PBMC was upregulated after stimulation with 10 μM CpG ODNs (Zhang et al., 2001). The concentration applied in our study was only 3 μM, which might have compromised the stimulatory effect. In addition, large variation among individuals in response to CpG ODNs was reported in humans, cows, and pigs (Kamstrup et al., 2001; Rankin et al., 2001; Leifer et al., 2003). The cellular receptor for CpG ODNs is TLR9 (Bauer et al., 2001). The expression of TNF-α and TLR9 was slightly increased in PBMC stimulated by the GACGTT, but not GTCGTT, CpG ODN. However, the relative gene expression ratio was relatively small (<2). In general, bovine PBMC treated by the GACGTT motif had more significantly upregulated cytokine genes than those treated by the GTCGTT motif. A report in humans indicated that genetic polymorphisms of TLR9 influenced the immune response to CpG ODNs (Kikuchi et al., 2005). Whether this is the case for bovine species requires further investigations.
Due to the in vitro effects of CpG ODN containing 3 copies of GACGTT motif, it was formulated with the KLH antigen to examine its effect on responses of KLH-specific antibodies and antigen-induced IFN-γ secretion in dairy cattle. The production of KLH-specific total IgG, IgG1, IgG2, and IgM in serum was elevated (P<0.05) in cows immunized with KLH+FICA+CpG in comparison with other vaccine formulations and the adjuvant control. It is noteworthy that an increased (P<0.05) production of IgG2 in serum was only observed in cows immunized with KLH+FICA+CpG, but not KLH+FICA and KLH+FICA+cCpG, in comparison with the control. Several previous studies have demonstrated that CpG ODNs containing 3 copies of GTCGTT motifs (assigned as 2006, 2007, and 2135) enhanced both humoral and cell-mediated immune responses of vaccines in bovine species (Ioannou et al., 2002; Rankin et al., 2002; Zhang et al., 2003; Wedlock et al., 2005a, 2005b; Mapletoft et al., 2006). Among all the subclasses of antibody, IgG2 has been the isotype that is analyzed most frequently as an indication of Th1 immune responses. CpG ODNs, when used as the adjuvant, is a strong inducer of Th1 immune responses, as characterized by increased production of IgG2 and IFN-γ expression in immune cells. Our results showed that CpG ODNs with GACGTT motifs not only enhanced the production of antigen-specific IgG1 and IgM, but also IgG2 which was not increased by other formulations. Both IgM and IgG2 are crucial opsonins for bovine phagocytes. Further, the responses of KLH-specific antibodies in serum were significantly much more persistent in cows immunized with KLH+FICA+CpG and remained high until 140 days after the primary immunization. Similar results were also reported in a recent study (Kovacs-Nolan et al., 2009), where CpG ODN alone, or in conjunction with other types of adjuvant, significantly induced long-lasting humoral responses in cows immunized with a hen egg lysozyme antigen.
The levels of KLH-specific total IgG, IgG1, and IgM, in milk were also elevated (P<0.05) in cows immunized with KLH+FICA+CpG. The increase of total IgG in milk was almost completely due to the increased level of IgG1 because they had very similar kinetic profiles. Immunization with KLH+FICA or KLH+FICA+cCpG only increased the level of KLH-specific IgG1 in milk, but not other subclasses. None of the vaccine formulations raised the level of KLH-specific IgG2 in milk. The concentration of antibodies in normal milk drops dramatically, as much as ∼100-fold lower, in comparison with those in colostrum and serum. Therefore, it was not surprising to see diminished antigen-specific antibody responses in milk. In bovine milk, the ratio of IgG2/IgG1 is much lower than that in serum. IgG2 has a higher affinity to FcRn, the receptor responsible for transportation of IgG across blood/milk barrier, and is recycled from milk back to blood at a faster rate (Medesan et al., 1998; Cianga et al., 1999). Therefore, the concentration of IgG2 is very limited in milk, which might not be able to reflect the increase of KLH-specific IgG2 in serum. In milk, the transient nature of IgM production was more obvious. The level of IgM dropped to that close to the basal level on D28.
The addition of CpG ODN with GACGTT motifs directed the immune responses to type Th1 as implied by the data of IFN-γ secretion in whole blood. This is in agreement with several other studies that used CpG ODN to enhance the efficacy of vaccines in bovine species (Rankin et al., 2002; Zhang et al., 2003; Wedlock et al., 2005b; Mapletoft et al., 2006). CpG ODNs did not increase the mRNA expression of IFN-γ in the in vitro assay. However, they significantly increased the expression of IL-12, which plays an important role in the development of Th1 immune cells, especially during the early stage of immunization.
Our results indicate that the CpG ODN containing 3 copies of GACGTT motif is a strong inducer of bovine lymphocyte proliferation, upregulated cytokine gene expression in bovine PBMC, and enhanced production of opsonic antibodies and secretion of IFN-γ. Although these preliminary data are still inconclusive, application of this CpG ODN as an adjuvant requires further investigations and may have positive effects on vaccines for dairy cows.
Footnotes
Acknowledgments
This study was supported by Grant NSC 95-2313-B-020-006 from the National Science Council of Taiwan and grants from the Dairy Cattle Genetics Research and Development Council of the Canadian Dairy Network and the Natural Sciences and Engineering Research Council of Canada.
Author Disclosure Statement
No competing financial interests exist.